蚕豆自噬基因鉴定及响应干旱胁迫分析
2021-05-25刘昌燕廖芳丽陈宏伟刘良军韩雪松万正煌卢碧林沙爱华
胡 杨,刘昌燕,廖芳丽,李 莉,陈宏伟,刘良军,韩雪松,万正煌*,卢碧林,沙爱华*
(1.长江大学农学院/主要粮食作物产业化湖北省协同创新中心/湿地生态与农业利用教育部工程研究中心,湖北 荆州 434025;2. 湖北省农业科学院粮食作物研究所/粮食作物种质创新与遗传改良湖北省重点实验室,湖北 武汉 430064; 3. 荆州市种子管理局,湖北 荆州 434020)
【研究意义】蚕豆(ViciafabaL.)属蝶形花科野生豌豆属,营养丰富并具固氮特性,能增加土壤氮素,缓解地力养分的消耗[1]。随着环境条件的不断变化,干旱等非生物胁迫逐渐成为影响农作物生长的限制因子,干旱造成的经济损失在众多非生物胁迫中居首位[2]。种子萌发是植物生长的起始阶段,土壤水分影响种子萌发,决定植物是否能够获得足够苗数,以保证获得足够的生物产量。蚕豆种子萌发需水较多,播种后如果干旱少雨,将导致种子萌发力下降,出苗少,蚕豆产量最终会受到影响[3]。自噬(Autophagy,ATG)是真核细胞面临营养缺乏时,在溶腔、液泡或溶酶体中降解大量细胞成分的系统[4]。在植物中,自噬作用响应各种环境胁迫,如养分缺乏、干旱、盐碱和高温,表明其具备用于提高作物应对环境胁迫的潜力[5]。因此,从蚕豆中鉴定自噬基因,分析其在种子萌发过程中对干旱胁迫的响应,将有利于鉴定出特定的自噬基因,用于改良蚕豆种子萌发耐旱能力,为培育高产耐旱蚕豆品种奠定基础。【本研究切入点】根据蚕豆转录组测序数据,鉴定蚕豆中ATG基因,并进行全面的生物信息学分析。【前人研究进展】ATG基因广泛存在于动物、植物、微生物中。许多ATG基因在动植物中功能保守[6]。目前,在酵母中已经鉴定出32个ATG基因[7],在拟南芥、水稻、马铃薯、辣椒、玉米中分别鉴定出了32、40、33、29、42个ATG基因[8-12]。其中一部分ATG蛋白的功能已得到验证。如过表达OsATG8c显著增加自噬活性,株高降低,有效分蘖和产量增加[13];番茄ATG18的激活提高番茄植株耐旱性,在干旱胁迫下降解泛素化蛋白质聚集体促进植株存活[14];过表达辣椒CaATG6提升了辣椒的耐热性[15]。自噬在植物生长发育过程中表达水平低,但在干旱等非生物胁迫下通过上调自噬水平帮助植物存活[16]。ATG基因过量表达增强自噬,能够提高植物产量和氮素转运效率[17-18]。但缺乏自噬会导致叶片过早衰老,并降低植物种子产量和质量[19]。在甘露醇诱导的渗透胁迫下,拟南芥ATG8家族中9个基因都有不同程度的上调表达,且渗透胁迫诱导的表达高于盐胁迫[20]。此外,渗透胁迫也诱导AtATG18a上调表达激活自噬[21]。在苹果中,自噬相关基因MdATG18a激活自噬基因ATG31提高了对病毒和干旱胁迫的抗性[22]。【拟解决关键问题】利用生物信息学方法鉴定蚕豆ATG基因家族,并分析其基因结构、亚细胞定位、蛋白保守结构域、系统进化发育、对干旱胁迫的响应,为进一步研究蚕豆ATG基因生物学功能,应用ATG基因提高蚕豆品种耐旱性奠定基础。
1 材料与方法
1.1 试验材料
供试蚕豆(ViciafabaL.)品种为埃塞俄比亚品种CDAS105和湖北品种鄂蚕1号,由湖北省农业科学院杂粮研究工程中心提供。主要试剂:RNA提取试剂盒(广东健仑生物科技有限公司)、甘露醇(天津市科密欧化学试剂科技有限公司)。
1.2 样品采集、RNA提取及转录组测序
试验在长江大学农学院耐渍油料种质资源湖北省重点实验室进行。于2019年7月选取形态大小均等的耐旱品种CDAS105和干旱敏感品种鄂蚕1号种子,分别放入浓度为0.8 %的甘露醇和灭菌蒸馏水的培养皿中,置于28 ℃恒温箱中萌发,萌发后16和64 h切取含胚芽的1/4部分子叶,用液氮速冻后保存于-80 ℃冰箱,用于提取RNA。每个处理设置3个生物学重复。转录组测序由武汉古奥科技有限公司完成。
1.3 基因注释及ATG基因筛选
以蚕豆转录组测序数据为基础,根据Swiss-prot数据库中注释为ATG基因,用TBtools软件[23]提取相应的蛋白序列后,在NCBI进行Blast,同时利用SMART在线软件(https:/smart.embl-heidelberg.de/)对序列的保守结构域进行预测,去除非ATG编码序列、重复序列及冗余转录本。根据[24]描述方法,对筛选出的蚕豆ATG基因进行分类和命名。
1.4 系统发育进化树构建
从NCBI(https://www.ncbi.nlm.nih.gov/)数据库中下载酵母ATG蛋白序列后,并与在EnsemblPlants(http://plants.ensembl.org/index.html)数据库中下载获得玉米、水稻、拟南芥ATG蛋白序列利用MEGA-X软件的ClustalW程序对蚕豆及拟南芥、玉米、水稻、酵母ATG蛋白进行多序列比对,采用NJ法(Neighbor Joining)构建系统发育树,通过Bootstrapmethod (BM)模型算法进行估计,并进行Bootstrap分析,重复值设置为1000。利用在线程序iTOL(https://itol.embl.de/)美化系统发育树。
1.5 生物信息学分析
蚕豆ATG基因结构通过TBtools软件进行可视化分析。亚细胞定位利用在线软件ProtComp9.0 (http://linux1.softberry.com/berry.phtml) 预测。蛋白理化性质通过在线软件ExPASy-ProtParam tool分析;保守基序(motif)类型和排列顺序软件为MEME(http://meme-suite.org/)。蛋白结构域通过SMART软件分析。
1.6 ATG响应干旱胁迫表达分析
蚕豆干旱胁迫转录组测序文库中基因片段表达量采用FPKM (fragments per kilobase million) 计算。将数据标准化后,利用TBtools软件构建ATG在2个品种中不同处理时间的表达热图。
1.7 数据分析
实验数据采用Excel2010进行整理及制图。
2 结果与分析
2.1 蚕豆ATG基因家族的鉴定及分类
基于干旱胁迫转录组测序数据,根据Swiss-prot蛋白数据库中的注释,初步筛选到117个ATG相关基因。进一步通过NCBI BlastP比对和保守结构域预测,最终得到12个ATG基因。通过与酵母ATG进行比对,将它们分为5个基因亚族,分别为ATG3基因(VfAG3),ATG6基因(VfATG6),ATG8基因(VfATG8a、VfATG8b、VfATG8c、VfATG8d、VfATG8e、VfATG8f);ATG18基因(VfATG18a、VfATG18b、VfATG18c)和ATG26(VfATG26)基因(图1)。
2.2 Vf ATG蛋白的进化分析
将蚕豆中鉴定的12个ATG与酵母、拟南芥、玉米、水稻中的ATG蛋白进行聚类分析,发现蚕豆的ATG分别与其他物种中对应的亚家族蛋白聚在同一分枝上,如VfATG3与拟南芥AtATG3、酵母ScATG3、玉米ZmATG3、水稻OsATG3位于同一个分枝上;VfATG8与ScATG8、AtATG8、ZmATG8、OsATG8 (abcdef)位于同一亚枝上;VfATG18、ScATG18、AtATG18、ZmATG18、OsATG18(abc)位于同一亚枝上(图2)。表明蚕豆中新鉴定这些蛋白与其他物种中同一亚家族蛋白结构和功能域相似。
2.3 蚕豆ATG蛋白的理化性质及亚细胞定位预测
蚕豆中鉴定的12个ATG蛋白的长度为118~623 aa,预测的蛋白分子量(Mr)大小为13 625.6~68 506.87(表1)。等电点(PI)4.8~9.06,其中VfATG8a、VfATG8b、VfATG8d、VfATG8e、VfATG8f、VfATG18c为碱性蛋白(PI为7.99~9.03),其他6个为中性蛋白(PI为4.8~7.0)。蛋白不稳定系数除VfATG8f和VfATG18b小于40外,其他蛋白均大于40,表明鉴定的大部分蚕豆ATG属于不稳定蛋白。12个蚕豆ATG平均亲水系数均为负值,表明其都是亲水蛋白。亚细胞定位预测除VfATG3位于细胞质,其余均位于细胞核外表明鉴定的蚕豆ATG蛋白可能主要参与细胞核外一些特定的生物学过程。

表1 VfATG理化性质及亚细胞定位预测
2.4 蚕豆ATG基因结构、蛋白结构域和保守基序分析
VfATG基因结构分析发现,VfATG基因家族成员无内含子结构,每个成员都含有UTR区与CDS区,同一亚族成员基因编码区长度大致相同(图3-A)。蛋白结构域分析显示,ATG3含有Autophagy_N、Autophagy_act_C和Autophagy_C 3个结构域,ATG6含有APG6、APG6_N 2个结构域其他蛋白都只含有1个结构域(图3-B)。不同亚家族拥有不同的结构域,但同一亚家族的结构域基本保守,如ATG18、ATG8(图3-B)。进一步分析ATG蛋白中的保守基序发现,VfATG蛋白中共鉴定出20个motif(mortif 1-mortif 20)),其长度为6~50个氨基酸(图3-C,表2)。在预测的20个motif中,motif 1第33~40位氨基酸具有保守性(3-D),motif 2第33~39位氨基酸也相对保守,motif 8中除了第5位氨基酸以外,其他位点氨基酸相对保守。ATG6拥有最多基序(motif 6,9,12,15,16,17,18,19),其次是ATG26(mortif 10,12,14,16,18,20)和ATG18(mortif 3,4,5,8,9,13,15)(图3-C)。不同亚家族含有的mortif各不相同,但同一亚家族蛋白之间具有相似的mortif,如ATG8 和ATG18(图3-C)。
2.5 VfATG响应干旱胁迫表达分析
根据转录组测序结果,分别提取鉴定的12个ATG的表达绘制热图(图4)。在干旱处理16 h后,ATG18b和ATG18c在抗旱品种CDAS105(T2)中较敏感品种鄂蚕1号(T1)表达上调,而ATG8c表达下调。在干旱处理64 h后,ATG8a在抗旱品种中较敏感品种表达上调。而在对照处理中,ATG8b和ATG8c分别在处理16 h的抗旱品种中上调表达和下调表达,ATG8a则在处理后64 h抗旱品种中上调表达。表明干旱处理16 h后,ATG18b和ATG18c迅速上调表达可能与CDAS105的抗旱性相关;而ATG8b在对照处理中上调表达,以及ATG8c在干旱和对照处理中均下调表达,可能是由于品种的生理特性差异决定的,与品种抗旱性差异关系不大。ATG8a在抗旱品种中干旱和对照处理64 h后均上调表达,表明其可能在抗旱品种中本底表达较高,从而导致抗旱品种的抗旱性更强。同时通过比较ATG基因在同一品种干旱处理不同时间表达发现,ATG8a在T2中干旱处理64 h较16 h表达上调,而ATG8e在干旱处理64 h后,在T2和T1中均较处理16 h表达上调。表明随着干旱处理时间的增加,ATG8a在T2中表达进一步增强,从而更有利于增强其抗旱性。而ATG8e在抗旱品种和敏感品种中均随着干旱处理时间的增加表达增强,表明其可能参与干旱胁迫的基础防卫反应,与品种的抗旱性差异关系不大。
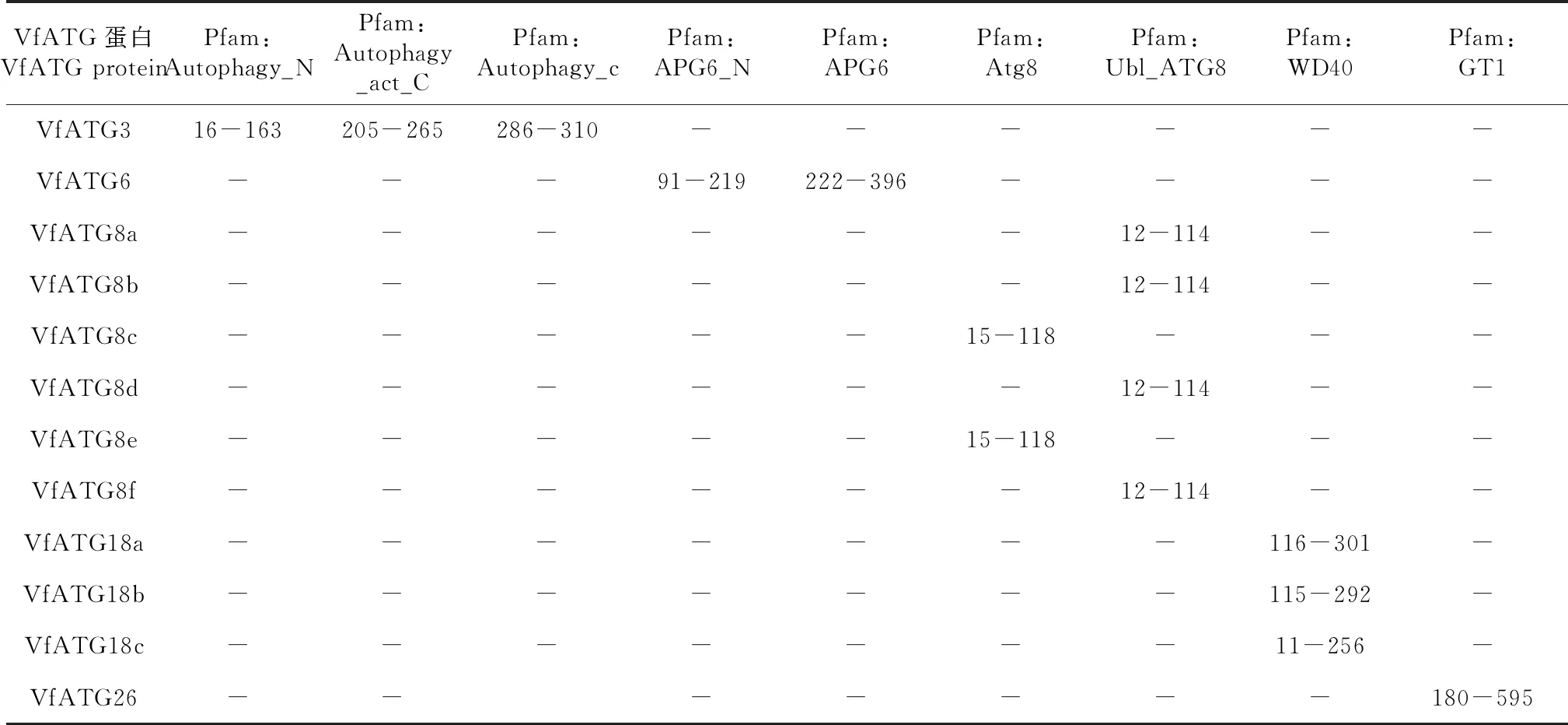
表2 VfATG蛋白结构域在肽链中的位置
3 讨 论
自噬基因响应养分缺乏、干旱、盐碱和高温等各种环境胁迫,可用于改良作物的抗逆性。尽管在拟南芥、水稻、玉米等植物中鉴定了一些ATG基因,但在蚕豆中还未见报道。
本文根据转录组测序结果,结合生物信息学分析,首次从蚕豆中鉴定了12个ATG基因。通过分析这些基因对干旱胁迫的响应,鉴定出ATG8a、ATG18b和ATG18c在蚕豆抗旱品种和敏感品种中存在表达差异,表明这些基因可能参与蚕豆种子萌发过程中的干旱胁迫调控。前人研究也发现,ATG8、ATG18家族成员在拟南芥[17-19,22,24,26]、番茄[12]、苹果[19]、水稻[27]中参与渗透或干旱胁迫调控。表明ATG8和ATG18基因家族成员在不同物种中具备调控干旱胁迫的功能保守性。因此,本文从蚕豆中鉴定的ATG8a、ATG18b和ATG18c可以作为潜在的基因资源,用于改良蚕豆种子萌发的耐旱能力,进一步应用于培育高产耐旱蚕豆品种。
此外,本文鉴定的12个蚕豆ATG基因分布于ATG3、ATG6、ATG8、ATG18和ATG26这5个亚基因家族,聚类分析中分别与水稻、玉米中的相应亚家族成员位于同一分枝。一般情况下,同一分枝上的亚家族蛋白具有相似的结构和功能域,因此功能也趋于保守。苹果中MdATG3a和MdATG3b响应叶片衰老、氮饥饿、干旱、盐和氧化胁迫,在拟南芥中过量表达MdATG3a和MdATG3b均增强渗透和盐胁迫耐性,并改善植物生长[24]。拟南芥中ATG6为花粉萌发和生长发育所必需[29],水稻中ATG6响应热、冷和干旱胁迫[28],过表达辣椒CaATG6提升了辣椒的耐热性[30]。拟南芥中过量表达ATG8提高N素重新移动效率和促进籽粒充实[31]。拟南芥中过量表达甜橙CsATG18a和CsATG18b,提高了转基因植物对渗透胁迫、盐、干旱和冻害的耐受性[32]。因此,蚕豆中鉴定的12个ATG除了ATG8a、ATG18b和ATG18c参与抗旱胁迫响应外,包括这3个基因在内的其他ATG可能还参与调控生长发育或其他环境胁迫反应,其生物学功能还有待于进一步鉴定,以期能充分利用这些基因改良蚕豆的相关性状,提高蚕豆生产的效益。
4 结 论
基于蚕豆干旱胁迫转录组测序数据,鉴定了12个蚕豆ATG基因,明确了其进化关系、基因结构、亚细胞定位、理化性质等,并筛选出VfATG8a、VfATG18b和VfATG18c响应蚕豆干旱胁迫,能够用于改良蚕豆种子萌发的耐旱性。
