大草蛉雄虫卵黄原蛋白基因的鉴定和表达特征
2021-05-11冯彦姣刘小平张婷婷陈红印王孟卿刘晨曦李玉艳张礼生毛建军
冯彦姣,刘小平,张婷婷,2,陈红印,王孟卿,刘晨曦,李玉艳,张礼生,毛建军*
(1. 中国农业科学院植物保护研究所,北京 100193;2. 东北农业大学,哈尔滨 150040)
大草蛉Chrysopa pallens属脉翅目Neuroptera草蛉科Chrysopidae,成虫、幼虫均可取食农、林、果、蔬上的蚜虫、叶螨、其他昆虫卵及低龄幼虫等多种农林害虫[1],是粮食作物、果蔬等经济作物害虫的重要捕食性天敌昆虫[2,3]。由于具有分布广、食量大、食性广的特征,在害虫的天敌防治中具有非常广阔的应用前景。
卵黄原蛋白(Vitellogenin,Vg)是卵生动物卵黄蛋白的前体物质[4],在大部分昆虫中,由性成熟雌虫的脂肪体合成并分泌到血淋巴,运输到卵巢后被发育中的卵母细胞选择性地摄取,在卵子内沉积形成卵黄蛋白,为胚胎发育提供氨基酸、脂肪、碳水化合物、磷和硫等营养和功能性物质[5]。随着研究的深入, 在少部分昆虫雄性中也发现有Vg的表达。比如在社会性昆虫意大利蜜蜂Apis mellifera的表达模式中发现,Vg基因不仅在蜂王的脂肪体中转录表达,在雄蜂和工蜂的脂肪体和蜂王的卵巢中也能检测到该基因[6,7];在交配过的米蛾雄性成虫中检测到vitellogenin mRNA[8]。此外,在大草蛉的雄虫中也检测到了Vg的存在[9]。昆虫卵黄蛋白的研究是近年来昆虫生理学和生化学比较活跃的领域之一[10],但是对大草蛉雄虫的卵黄原蛋白的表达规律及功能研究还很少有文献报道。
本研究基于大草蛉转录组数据库设计引物,克隆获得1个大草蛉Vg蛋白基因的全长序列,并采用生物信息学方法对其序列进行分析;通过荧光定量PCR方法检测大草蛉vitellogenin基因在不同发育阶段、不同组织的表达特征,以期为深入研究vitellogenin功能奠定基础。
1 材料与方法
1.1 材料
供试昆虫:本试验中使用的大草蛉为中国农业科学院植物保护研究所天敌昆虫组室内连续多代饲养,饲养条件为温度(25±1)℃,光周期16L:8D,相对湿度为75%±10%。
试剂:TransScript® One-Step gDNA Removal and cDNA Synthesis SuperMix,Trans2K DNA Marker,6×DNA Loading Buffer,北京全式金生物技术有限公司;感受态细胞Trans 5α Chemically Competent Cell,克隆载体pEASY-T1,2x High-Fidelity Master Mix,北京擎科新业生物技术有限公司;Go Taq®qPCR Master Mix,promega公司;琼脂糖,江晨生物公司;质粒小提试剂盒,北京天根生化科技有限公司;所有引物合成和DNA测序,生工生物工程(上海)股份有限公司。
仪器:基因扩增仪ETC811,东胜公司;7500 Real-Time PCR Sys-tem荧光定量PCR仪,美国Applied Biosystems公司;台式离心机(sigma1-14);水平电泳仪,北京六一生物有限公司;凝胶成像分析仪 WD-9413B,北京六一仪器厂;NanoDrop 2000超微量分光光度计,美国Emplen公司;旋涡震荡仪QL-901,海林市其林贝尔仪器制造有限公司;微型离心机Mini-4K,珠海黑马医学仪器有限公司;电子天平,北京赛多利斯仪器系统有限公司;超净实验工作台HD-1360,哈东联公司;立式压力蒸汽灭菌锅LS-B5OL-I,江阴滨江医疗设备有限公司;微波炉 GalanzP7021TP-6。
培养基:LB(Luria-Bertani,LB)液体培养基:胰蛋白胨10 g、酵母提取物5 g、氯化钠10 g、蒸馏水1000 mL;LB固体培养基:在LB液体培养基中加入琼脂15 g。
1.2 大草蛉雄虫总RNA提取和cDNA第一链合成
将单头大草蛉雌虫在液氮中研磨成粉末状,并迅速加入TENS缓冲液,1头大草蛉约加缓冲液1800 μL(分装到离心管中,300 μL/管),37 ℃下保育3~18 h。每管加入85 μL 5 mol/L 的NaCl,旋涡混匀15 s,1400 r/min离心5 min,用70%乙醇洗一次。室温下干燥沉淀(置于通风橱中,5 min),加入30~40 μL无菌水溶解。采用紫外微量分光光度计确定RNA浓度和质量,采用凝胶电泳检测总RNA的完整性,采用PrimeScript RT Reagent Kit进行反转录,所得cDNA于-20 ℃冰箱中保存,备用。
1.3 大草蛉vitellogenin基因克隆
根据已知的大草蛉雌虫卵黄原蛋白基因序列(MH400048)的信息,分别设计 1对简并引物 A1:5′-TCTTCATTACAGCACTTGGC-3′;B1:5′-ATTCTCGGCTGTATCCTTCA-3′。利用 ORF finder软件对其进行分析,其具有完整开放阅读框(open reading frame,ORF)。为验证序列的准确性,用于PCR扩增,引物均由生工生物工程(上海)股份有限公司合成。20 μL扩增反应体系:2×PrimeSTAR Max Premix 10 μL、ddH2O 7 μL、上下游引物各 1 μL、cDNA 1 μL。PCR 扩增反应条件:98 ℃变性 30 s,55 ℃退火 5 s,72 ℃延伸1 min,共35个循环。对PCR产物进行凝胶电泳检测并分离回收,将回收产物连接到pEASY-T1载体,然后转化到大肠杆菌Trans 5α感受态细胞中。选取已鉴定为阳性克隆的菌液,委托生工生物工程(上海)股份有限公司进行测序。
1.4 大草蛉卵黄原蛋白基因vitellogenin理化性质及结构域预测
使用ExPASy-ProtParam tool(http://web.expasy.org/protparam)软件分析vitellogenin基因编码蛋白的等电点及分子量。使用TMHMM 2.0(http://www.cbs.dtu.dk/services/TMHMM/)在线软件对vitellogenin氨基酸序列的跨膜结构域进行预测。使用SingalP 3.0软件进行vitellogenin蛋白信号肽的预测。利用DNAMAN软件进行氨基酸序列的多重比对和亲/疏水性分析。
1.5 大草蛉卵黄原蛋白的同源比对及进化树分析
从NCBI数据库中获得多个物种的vitellogenin基因,使用ClustalW将大草蛉与获得的物种vitellogenin序列进行同源比对后,使用软件Mega 6以邻接法(neigh-bor-joining,NJ)经1000次抽样分析将部分昆虫vitellogenin基因构建系统进化树。
1.6 大草蛉卵黄原蛋白基因的表达分析
测定大草蛉成虫羽化不同时间vitellogenin基因相对表达量:分别取羽化后第6、12、18、24、30 d的雄成虫,每个样品取样为雄成虫1头。
测定大草蛉成虫不同组织vitellogenin基因相对表达量:分别取羽化后第15 d的大草蛉的雌、雄成虫以及雄成虫的头、胸、腹部位。每个样品取样为雌、雄虫各1头,头3个,胸3个,腹2个。所有样品均设置3次以上重复,于液氮中速冻并保存于-80 ℃备用。根据测序结果设计定量引物:Q-AF:5′-AAGGAGC CGTATATGATGTTGC-3′;Q-AR:5′-TTTCAGCGGGACTTGTGG-3′,在生工生物工程(上海)股份有限公司合成,并通过普通PCR优化反应条件体系。选Actin作为内参基因,引物为:AcF:5′-AACTTCCCGAC GGTCAAGTCAT-3′;AcR:5′-TGTTGGCGTACAAGTCCTTACG-3′。采用以 SYBR Green 为染料的荧光标记法进行定量检测。20 μL qPCR体系:2×Fast Start Es-sential DNA Green Master 10 μL、引物各1 μL、模板cDNA 1 μL、ddH2O 7 μL。反应条件:95 ℃预变性3 min;95 ℃变性10 s,60 ℃退火10 s,72 ℃延伸10 s,共40个循环;95 ℃恒温10 s,65 ℃恒温60 s,97 ℃持续1 s制备熔解曲线,37 ℃冷却30 s。并使用2-△△Ct法[11]计算vitellogenin基因的相对表达量。
1.7 大草蛉卵黄原蛋白的表达分析
通过SDS-PAGE检测。测定大草蛉雄成虫不同组织卵黄原蛋白的相对表达量:分别取羽化后第15 d的雄成虫及头、胸、腹部位。每个样品取整虫1头,头3个,胸3个,腹2个。所有样品均设置3次以上重复,于液氮中速冻并保存于-80 ℃备用。
1.8 数据统计与分析
使用GraphPad Prism 6.0软件对结果进行统计分析,大草蛉卵黄原蛋白基因mRNA定量测定数据采用ANOVA单因素方差分析,采用Tukey’s HSD法进行差异显著性检验。
2 结果与分析
2.1 大草蛉卵黄原蛋白基因的克隆与进化分析
以大草蛉cDNA为模板进行vitellogenin基因的扩增,经测序拼接得到产物的大小为5561 bp,其中包含完整开放阅读框(ORF)1501 bp,编码501个氨基酸残基。vitellogenin编码蛋白含有GLVG 基序、半胱氨酸残基、DGXR基序、DXXR基序等(图1),说明产物为大草蛉卵黄原蛋白基因。通过软件分析可得,其成熟肽的预测相对分子量为206 kD,理论等电点为8.11,其中包含酸性氨基酸残基(Asp+Glu)189个,碱性氨基酸残基(Arg+Lys)194个。其分子式为C16975H28519N5433O6979S1060,蛋白的不稳定系数为44.00,推测其为不稳定蛋白,且属于分泌性蛋白。跨膜结构域预测结果显示:Vg不存在跨膜螺旋区,其蛋白亲水性总平均值0.874,脂肪指数为36.06。利用DNAMAN将其与雌虫的Vg1基因序列比较发现两者的相似度高达99.8%,如图2。

图1 大草蛉部分vitellogenin序列及其推导的氨基酸序列Fig. 1 Partial vitellogenin gene sequence and its derived amino acid sequence

图2 大草蛉雄虫部分vitellogenin序列与雌虫Vg1的比对Fig. 2 Comparison of partial vitellogenin sequence between Chrysopa pallens males and females Vg1 gene
利用NCBI对大草蛉雄虫Vg基因序列进行同源检索,并选取了半翅目、网翅目、鞘翅目、膜翅目、鳞翅目的23个代表性昆虫的基因序列。BLAST比对结果表明,雄虫卵黄原蛋白基因序列与骚扰锥蝽Triatoma infestans(GenBank登录号:AIA09041)的相似性最高,为32.75%,与其他昆虫vitellogenin基因一致性不太高。基于膜翅目、鞘翅目的23种vitellogenin氨基酸序列,以邻接法构建的进化树结果显示,vitellogenin分别聚为4支,大草蛉vitellogenin与网翅目的德国小蠊、马德拉蜚蠊先聚合,其次与半翅目的大田鳖、骚扰锥蝽、赤须盲蝽、褐飞虱、琉璃叶蝉聚为一支;而鞘翅目的异色瓢虫、大猿叶甲、黄粉虫、疣状天牛、棉铃象甲以及水椰八角铁甲聚为一支;膜翅目的丽蚜小蜂、蝶蛹金小蜂、日本瘤姬蜂、角额壁蜂聚为一支,鳞翅目的舞毒蛾、斜纹夜蛾、稻纵卷叶螟、野桑蚕及家蚕则聚为一支。vitellogenin聚类结果能较好地反映昆虫间的亲缘关系。
2.2 vitellogenin基因表达分析
2.2.1 大草蛉雄虫不同发育阶段vitellogenin基因的表达分析 大草蛉vitellogenin基因表达不具有明显的发育阶段特异性,在羽化后第6 d表达量最低,但是之后的表达水平基本呈现稳定趋势,差异不大,这一结果与前面研究人员的结果是相吻合的[13]。
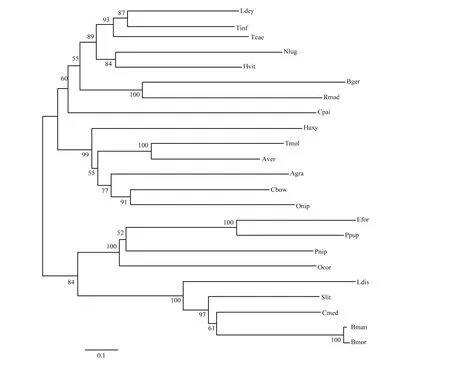
图3 基于vitellogenin氨基酸序列以邻接法构建大草蛉和其相近昆虫的系统发育树Fig. 3 Phylogenetic tree of vitellogenin from Chrysopa pallens and related different insect species by using neighbor-joining method.
2.2.2 大草蛉雄虫不同组织vitellogenin基因的表达分析 大草蛉雄虫vitellogenin基因的表达具有组织特异性。vitellogenin基因的表达量在雄虫的腹部最高,其表达量分别为头部表达量的3.51倍(F1,4=99438,P<0.01),为胸部表达量的 2.64倍(F1,4=949.778,P<0.01)(图 4)。雄虫头部和胸部的 vitellogenin表达量没有显著差异,表达丰度基本相当。但是雌成虫中的表达量均显著高于雄虫及各组织部位的表达量。

图4 vitellogenin基因在大草蛉雄虫羽化后不同天数的相对表达量Fig. 4 Relative expression of vitellogenin mRNA in male in different days after emergence
2.2.3 不同组织 vitellogenin的表达分析 对大草蛉雄成虫各组织部位的卵黄蛋白进行提纯与 SDS-PAGE电泳分析后发现, 其整虫和腹部的卵黄蛋白分别由约165和46 kD左右的大小亚基组成,但是雄虫的头部和胸部的卵黄原蛋白只有小亚基的部分,没有检测到大亚基。
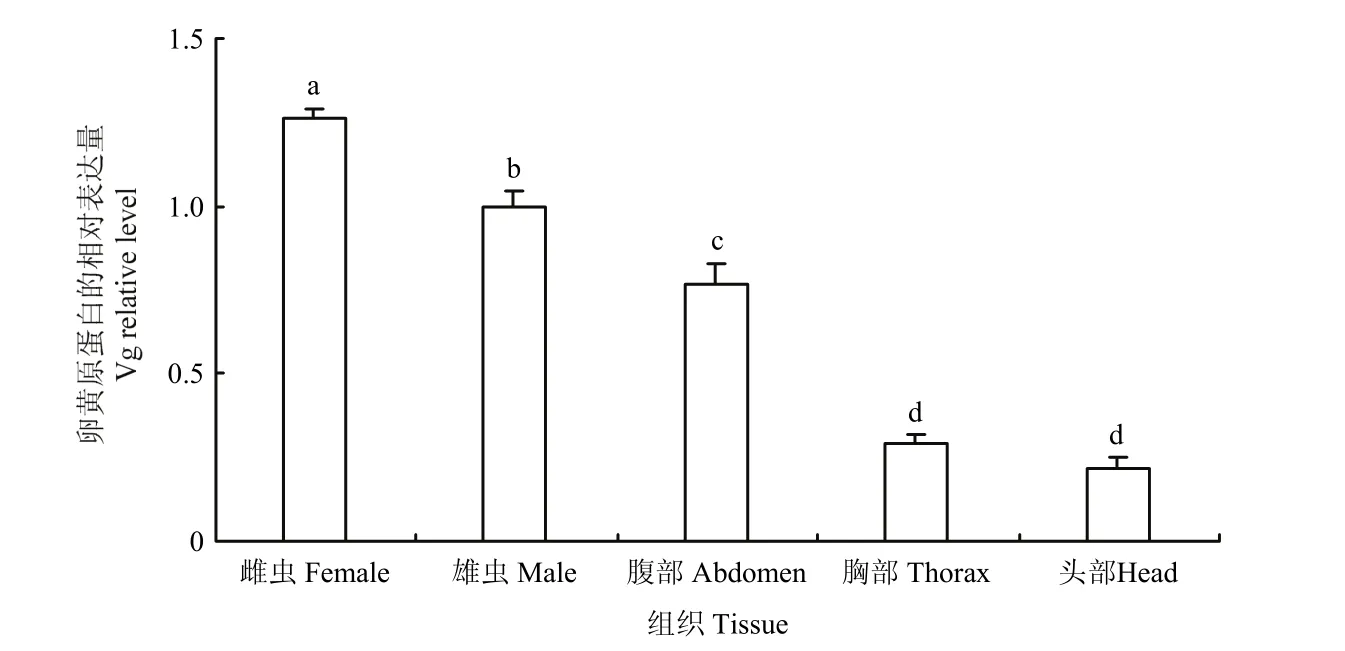
图5 大草蛉vitellogenin基因在雌雄成虫及不同组织中的相对表达量Fig. 5 Relative expression of vitellogenin mRNA in female and male adults and different tissues
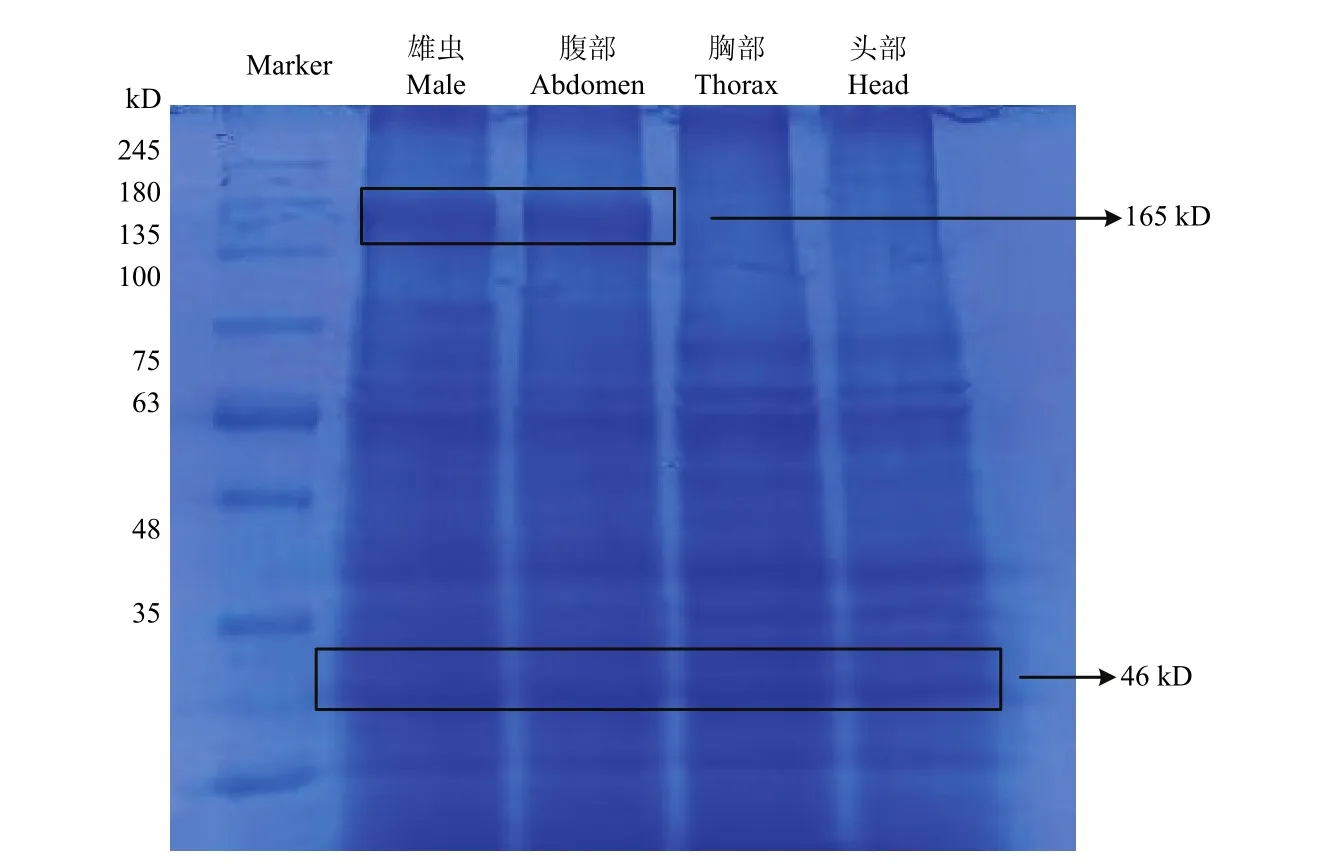
图6 大草蛉vitellogenin蛋白在雌成虫及不同组织中的表达Fig. 6 Expression of vitellogenin protein in female adult and different tissues
3 讨论
本研究根据大草蛉雌虫的卵黄原蛋白序列信息数据设计了雄虫 Vg的基因引物,成功克隆了雄虫 Vg基因的部分cDNA序列。经变性聚丙烯酰胺凝胶电泳检验了其表达,大草蛉雄虫卵黄原蛋白由约165 kD和44 kD大小的两个亚基组成,与陶淑霞等[13]的研究结果一致。根据蛋白氨基酸序列特征分析,Vg具有卵黄原蛋白的典型保守特征,包含GL/ICG 基序、DGXR基序、半胱氨酸残基等结构域,研究表明GL/ICG基序和半胱氨酸残基在昆虫胚胎形成过程中发挥功能性作用[17,18]。D、E、P、Y等氨基酸残基在C-末端高度保守。此外,GL/ICG基序是鉴定卵黄原蛋白基因的依据[19]。这与天蚕、中华蜜蜂、小菜蛾、椭圆食粉螨等昆虫Vg的研究结果一致[12-16],且本研究的系统进化分析结果显示,亲缘关系较近的昆虫Vg发育树的位置也较近,与传统形态学分类的结果是一致的。
RT-qPCR结果表明,雄成虫Vg在不同发育阶段的表达不具有显著性差异,表达初期Vg表达量最少,之后的表达趋势趋于稳定,这种趋势和已报道的大草蛉雌虫中先增加后逐步下降的趋势完全不同[10]。羽化初期Vg基因的表达量较低可能由于该时期各神经、生殖等系统尚未发育完全,因此其功能也不能完全发挥。大草蛉雄虫Vg基因在头、胸和腹部的转录表达结果显示,Vg基因均可以在大草蛉雄虫的头、胸和腹部表达,其中头部表达丰度最低,其次是胸部,而腹部的表达量最高,极显著高于头部和胸部。这一结果与中华蜜蜂工蜂中的Vg基因表达结果是一致的[20]。但是,大草蛉雌虫Vg基因的mRNA表达丰度是显著高于雄虫及其各组织的。SDS-PAGE结果显示,大草蛉雄成虫整虫和腹部的卵黄蛋白均由165和46 kD的大小亚基组成,与大草蛉卵和雌虫Vg蛋白一样,属于第1类型表达模式,而头部和胸部的Vg蛋白只含有小亚基,与果蝇、家蝇等高等双翅目昆虫的Vg蛋白一样,属于第3类型表达模式[11]。Vg是一种雌性蛋白,由雌性脂肪体合成。为揭示雄虫Vg蛋白的合成部位,后续还需要分别从雄虫的脂肪体、睾丸、血淋巴等器官或组织提取蛋白以进行鉴定。
在大部分昆虫中Vg是在昆虫的脂肪体中合成的,在对社会性昆虫意大利蜜蜂的表达模式研究中也发现,Vg不仅在蜂王的脂肪体中转录表达,在雄蜂和工蜂的脂肪体和蜂王的卵巢中也均能检测到[21]。卵黄原蛋白是合成卵黄蛋白的前体物质,与卵黄的生成密切相关,在生殖调控中有着非常重要的作用。Moriyama等[22]发现在臭虫中Vg基因被干扰后,它体内相关营养物质在从脂肪体到卵母细胞的转运过程中受阻,从而抑制了产卵;在棉铃象甲中,Vg的表达被沉默后,虽然不影响象甲的产卵量,但会显著降低有效卵的产生[23]。这些结果均说明,Vg基因无论是直接影响产卵还是间接影响孵化,都在昆虫繁殖中起到不可忽视的作用。有报道称,在社会性昆虫蜜蜂中,Vg不仅与生殖行为相关,还有一些特殊的功能,比如还可以帮助蜜蜂适应气候、延长寿命和转化食物,通过清除自由基,减少氧化应激,从而延长工蜂和蜂王的寿命。此外,卵黄原蛋白还与蜜蜂的气候适应、生殖竞争等相关[24-27]。还有报道发现,蜜蜂卵黄原蛋白的合成受到保幼激素和蜕皮激素等的严格调控[28]。卵黄原蛋白基因是编码卵黄原蛋白的结构基因,是大草蛉最重要的功能基因之一[29]。截止目前,对于大草蛉雄虫卵黄原蛋白基因表达调控的研究未见报道。其功能以及基因表达调控机制是怎么样的呢?这些课题都亟待深入研究。尤其是在天敌昆虫应用越来越广泛的今天,卵黄原蛋白基因的研究对大草蛉的扩繁提供基础,为开发实际应用大草蛉这一资源昆虫提供支持。
本研究对大草蛉雄虫的卵黄原蛋白Vg基因进行了克隆,分析了该序列与其他昆虫卵黄原蛋白之间的进化关系,并完成了在不同发育阶段、不同组织的表达规律的测定,确定了Vg的基本性质,对比其他昆虫卵黄原蛋白基因的研究,推测Vg基因对生殖活动和行为的调节起重要作用。这些结果丰富了对雄虫卵黄原蛋白基因的认知,并为进一步探索雄虫卵黄原蛋白基因对其生殖等行为的调控机制提供了理论基础。
