栗山天牛成虫取食、交配和产卵行为
2020-07-07唐艳龙王丽娜张彦龙杨忠岐王小艺
唐艳龙,魏 可,王丽娜,张彦龙,杨忠岐,王小艺*
(1. 遵义师范学院生物与农业科技学院,贵州省赤水河流域动物资源保护与应用研究重点实验室,贵州遵义 563002;2. 中国林业科学研究院森林生态环境与保护研究所/国家林业局森林保护学重点实验室,北京 100091)
栗山天牛MassicusraddeiBlessig,属鞘翅目Coleoptera天牛科Cerambycidae山天牛属(陈世骧,1959),主要危害辽东栎QuercusliaotungensisKoidz和蒙古栎Q.mongolicaFisch. ex Ledeb等栎属植物,以幼虫钻蛀寄主韧皮部和木质部为害(唐艳龙,2011)。目前,国内大部分省份均有分布,国外分布于俄罗斯、朝鲜和日本。在我国,尤以在吉林、辽宁省和内蒙(赤峰市)危害最甚(唐艳龙等,2010)。据辽宁省1999年测算,栗山天牛危害失去利用价值的木材达200万m3(孙永平,2001)。2005年吉林省栗山天牛成灾面积8万hm2,直接经济损失达10余亿元(皮忠庆等,2007)。而且其危害有逐年加重的趋势。目前,栗山天牛已成为我国东北以栎木为主的天然林区的头号害虫(Yangetal.,2013),研究经济、安全、有效的防治技术成为亟待解决的问题。
栗山天牛成虫是裸露活动的唯一虫态,有取食、交配等群体活动行为。近年来,根据成虫的行为特点,探索大面积治理的途径,受到较为广泛的关注,也取得了明显的进展(高纯等,2008;姜静等,2010)。目前有关天牛成虫的交配行为已有较多的研究(Wang & Zeng, 2004; Wang & Davis, 2005)。如贺萍和黄竞芳(1993)把光肩星天牛AnoplophoraglabripennisMotsch.成虫的交配行为分为徘徊、接近、就位、舔翅(可以跳过)和交配5个阶段。杨洪等(2007)把松褐天牛MonochamusalternatusHope一次完整的交配分为相遇抱对、插入输精和配后保护3个阶段。多数天牛有交配后的保护行为(Hughes, 1981; 杨洪等,2007)。对于栗山天牛的取食、交配和产卵行为,侯义等(2000)简单描述过,但不甚详细。为了摸清栗山天牛成虫的取食、交配和产卵行为,找到新的防治措施,本文采用室内室外相结合的办法,观察了栗山天牛成虫的行为特点。
1 材料与方法
1.1 取食行为的观察
2014年成虫期将人工捕捉的栗山天牛成虫置于3.5 m×3.5 m×3 m的纱笼内,放养24 h。选健康成虫(无受伤迹象,触角、翅和足完整,爬行和飞翔速度较快)3对置于1.5 m×1.5 m的纱笼内,同时将蒙古栎、辽东栎、水曲柳FraxinusmandshuricaRupr.、三角枫AcerbuergerianumMiq.等林间常见树种的完整树叶、嫩枝和多年生枝条及树皮置于纱笼内,每天用洒水壶向笼内喷水两次,所选材料每天更换1次。观察成虫的取食情况直至死亡。设重复6次。
2014年在成虫活动高峰时间,即晚上19 ∶00-23 ∶00,在林间观察记载成虫的取食情况,并照相。连续观察3 d。
1.2 相遇、攻击、交配和产卵行为的观察
选择人工捕捉的栗山天牛健康成虫50对,置于3.5 m×3.5 m×3 m的纱笼内,放长约1 m,胸径大于20 cm的辽东栎树干6段,供产卵,放带有树叶的辽东栎小枝20段供攀附和休憩,放浸入糖醋酒液的棉花团20个供补充营养。经过24 h的对环境适应后,观察栗山天牛成虫的相遇和攻击行为,包括雌成虫与雌成虫相遇、雌成虫与雄成虫相遇、雄成虫与雄成虫相遇及雌成虫或雄成虫与交配对的相遇。观察记载栗山天牛成虫每次发生争斗的目的及争斗的主要方式。同时观察记录栗山天牛成虫的交配过程,记录每次交配开始和结束的时间、交配过程中插入输精的次数和持续时间、间歇次数和持续时间、配后保护时间及插入输精的过程。观察栗山天牛雌成虫产卵的行为,记载产卵的过程、产卵的部位及所产卵粒的颜色等信息。
1.3 数据分析
用Excel 2003和SPSS 11.0等专业软件进行数据处理,用SPSS中的One-Way ANOVA进行方差分析,用Duncan氏新复极差法检验,比较各处理之间的差异水平。
2 结果与分析
2.1 栗山天牛成虫的取食行为
室内试验发现栗山天牛成虫既不取食树叶,亦不取食树皮,只会舔吸树叶上的水分。野外观察发现,其主要吸食从栎树树皮受伤处溢出的汁液补充营养,天牛有强大的上颚,能够自己咬破树皮,但主要还是在已有的伤口处吸允,成虫往往聚集取食,属于因补充营养引起的聚集行为(图1)。另外,天牛与胡蜂、蛾类等竞争食物。在取食过程中,栗山天牛不会攻击和它竞争食物的其它种类昆虫,但会攻击同类。
2.2 栗山天牛成虫的相遇和攻击行为
当两头栗山天牛雌成虫相遇时会相互避让,只有当1头雌成虫遇到了交配对时,偶尔会出现将交配对拆散的情况,而且多数情况下是交配中的雄成虫放弃正在与之交配的雌成虫,转而追逐从旁边经过的这头雌成虫。交配中的雌成虫不理睬任何从身边经过的成虫,即使是交配对象被赶走也不攻击。也就是说,雌成虫一般不会主动攻击同类(表1)。

图1 补充营养的栗山天牛成虫Fig.1 Feeding of Massicus raddei adult
栗山天牛雄成虫比雌成虫更具攻击性,当两头雄成虫在爬行的过程中相遇时,一般会相互避让,很少会发生打斗;雄成虫在争夺食物和雌成虫的过程中,极易发生打斗(表1)。当1头雄成虫遇到取食中的另一雄成虫时,约有45%的概率发生打斗(n=29),约有一半的雄成虫会选择避让,只有极少数的雄成虫会接纳后来者共同取食,取食中的雄成虫赶走入侵者的概率在50%左右(n=18)。当1头雄成虫遇到交配对时,发生打斗的概率在80%以上(n=34),一般是交配中的雄成虫驱赶靠近的雄成虫。同时,交配中的雄成虫也更具攻击性,对于从身边触角范围内经过的天牛(不分雌雄)均表现攻击。交配中的雄成虫赶走入侵者的概率高达85%(n=46),只有极少数会失败,亦有少数雄成虫被干扰后主动放弃交配,这种情况在交配后期发生的概率较高。
栗山天牛雄成虫遇到爬行中的雌成虫时,有相当一部分会追逐进而试图交配,雄成虫获得成功的概率约在70%左右,只有少数雌成虫会逃脱(表1)。当雄成虫遇到正在产卵的雌成虫时,90%会选择离开,亦有少数会强行交配或者爬到雌成虫体背等待机会。当遇到正在取食的雌成虫时,部分雄成虫会试图交配,也有部分雄成虫会攻击雌成虫,将雌成虫赶走,进而占领取食地补充营养。
栗山天牛的攻击方式主要是用上颚咬住对手的触角或者足进行拖拽。种群密度和资源的多寡(如食物或雌成虫的多少)是影响种内竞争的重要因素。争斗的结果,多数情况是其中1头雄成虫战败逃走,获胜的雄成虫一般占据雌成虫或者占领取食地补充营养,不追赶攻击;有时其中1头雄成虫会被咬坏足或者触角,甚至咬断;偶尔有旗鼓相当的,野外观察到最长的争斗时间超过 30 min。多数成虫在被攻击时,会发出“吱吱”的声音。
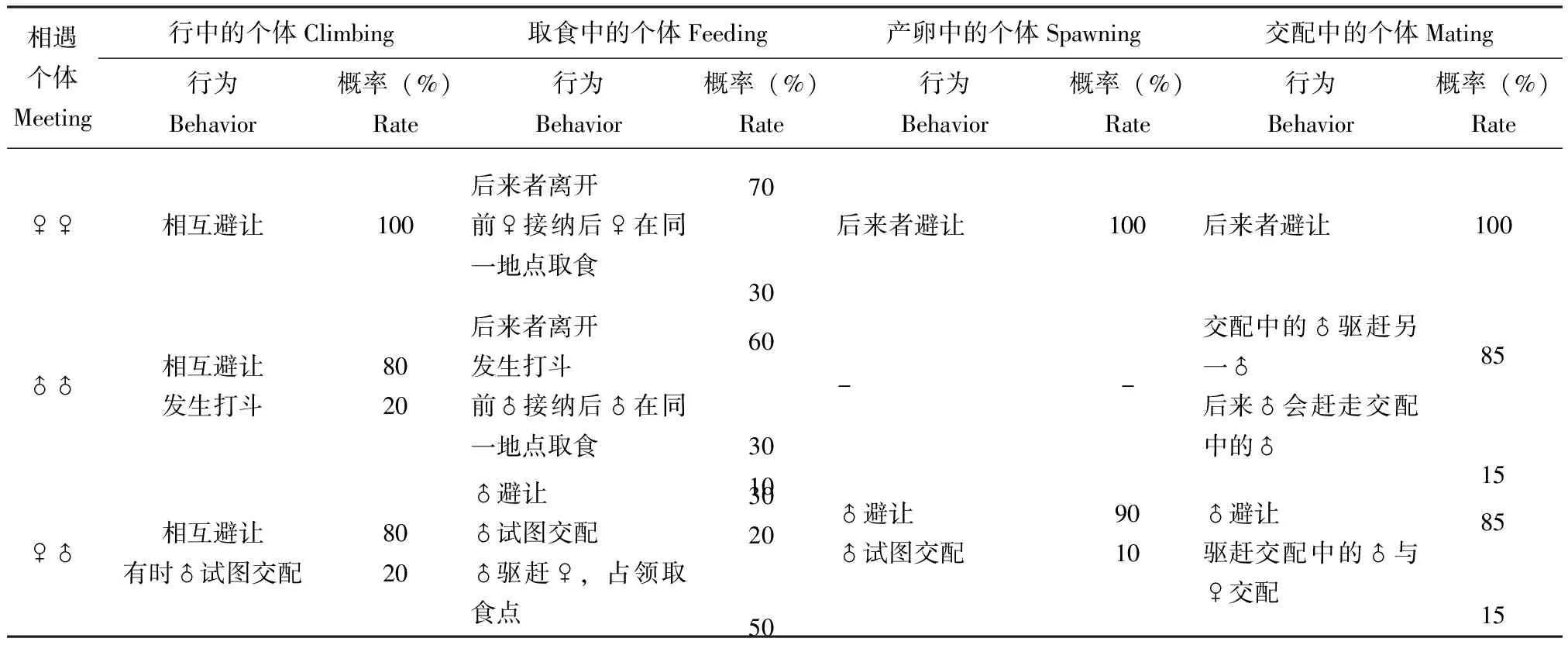
表1 栗山天牛成虫的相遇行为观察
70%的栗山天牛雄成虫争斗是为了争夺配偶,与其它目的差异极显著(F=1294.096,df=2,8,P=0.0001)(图2),其次是为了争夺食物资源,比例略低于30%。只有极少数雄成虫打斗是因为其它原因,观察到两头雄成虫狭路相逢而发生打斗,还观察到1头雄成虫在追逐雌成虫失败后去攻击旁边的雄成虫。
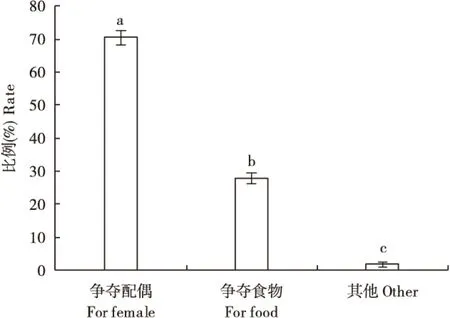
图2 栗山天牛雄成虫争斗的目的Fig.2 Fighting purpose of Massicus raddei male
2.3 栗山天牛成虫的交配行为
2.3.1栗山天牛成虫的交配过程
栗山天牛成虫完成一次完整的交配经过4个阶段,即:识别、追逐-攀附、抚慰、试探性插入-插入输精、输精后插入保护、交尾间歇-配后保护、离开(图3)。
在一定的距离内,栗山天牛雄成虫会不停晃动触角,定位雌成虫的方位。当两头天牛成虫相遇时,触角会在第一时间进行接触,接触时间持续不到1 s即能决定是否交配,判断触角上可能有接触性信息素。栗山天牛雄成虫选择配偶行为上占有主导地位。当有交配欲望的雄成虫碰到雌成虫,即进行追逐,持续时间较短,一般约2 s。雄成虫追逐到雌成虫后即强行攀附到雌成虫的背部,用1对前足在雌成虫鞘翅的前端外缘抱握住雌成虫,用中足和后足攀附住寄主树干支撑身体,并尝试交尾和抚慰背部。这个过程视雌成虫状态持续时间长短不一,如果雌成虫较配合,约1~2 s后即进入交尾,如果雌成虫不配合,则雄成虫用下颚须在雌成虫鞘翅的翅基区抚慰,待雌成虫安静后再进行交配。这种情况下需经过较长的时间才能进入交尾,而且往往容易挣脱造成交配中断,在受到干扰后也更易中断交配。有交配欲望的雄成虫选择雌成虫交配的成功率很高,极少有雌成虫能够逃脱。
在正式输精之前,雄成虫往往会插入生殖器1~2次,每次持续1~2 s。这一过程,可能是雄成性在探测雌成性的发育程度。在这一过程之后,有相当一部分雄成虫会离开雌成虫。
插入输精和交尾间歇是整个过程的重点,持续时间长短不一。雄成虫前几次输入精子时持续时间较长,之后持续时间逐渐下降。雄成虫一次插入后输精的过程可以分为3个阶段,开始约1 s/次,持续3~5次,之后加快到约0.5 s/次,持续1~20次,最后有3~4 s的时间停止动作。之后雄成虫休息约3~5 s,即再次插入生殖器,整个过程持续时间不等。随后,即进入较长时间的交尾间歇,交尾间歇持续的时间与交配次数关系较大,前几次交尾时,间歇时间较短,之后随着交尾次数的增加,间歇时间逐渐增加。在这一过程中,插入输精和交尾间歇会循环重复进行,次数与雌雄成虫的交配欲望和以往的交配次数有关,交配次数较少的雌雄成虫这一过程持续时间较长。
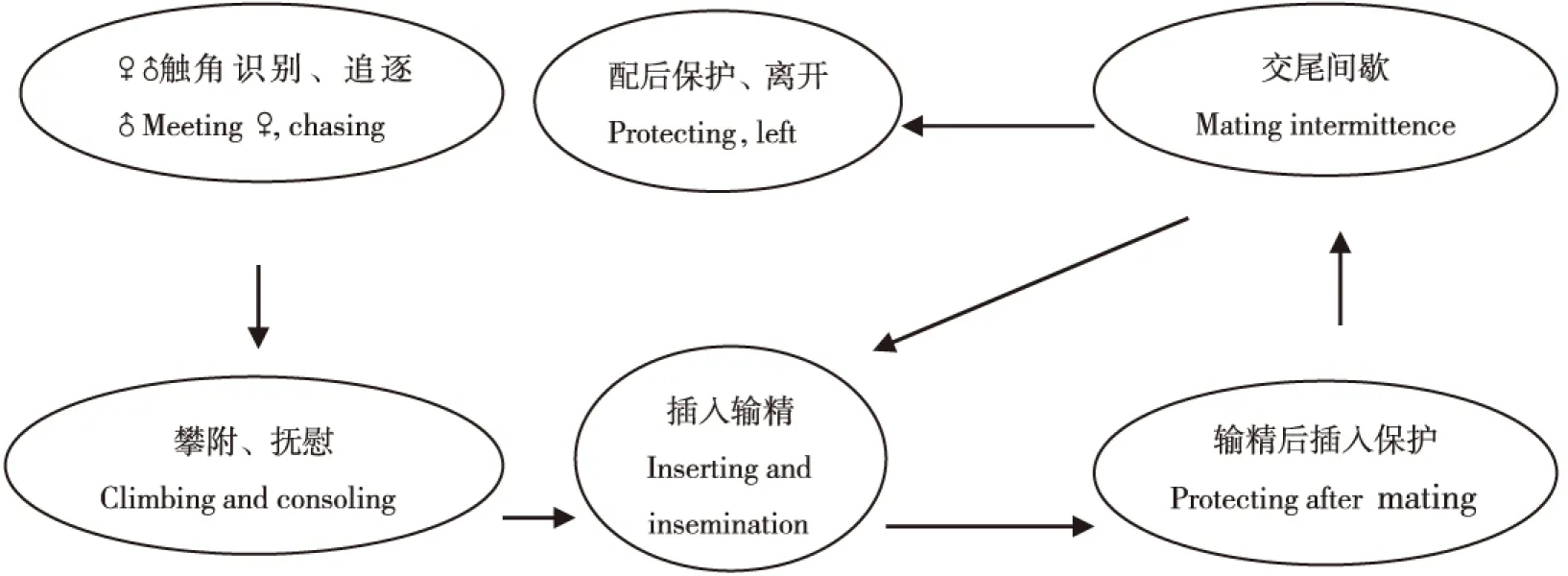
图3 栗山天牛的交配过程示意图Fig.3 Mating process of Massicus raddei adult
配后保护这一过程视雄成虫状况不同,有的雄成虫交配完之后就直接离开,几乎没有这一过程,特别是交配持续时间较短的情况。而部分雄成虫有明显的配后保护过程,即交尾完全结束之后,雄成虫攀附在雌成虫体背,并驱赶附近的雄成虫,有的甚至会将头部调向雌成虫腹部保护。观察到最长的配后保护时间达8 min。交配完成后雄成虫即自行离开,寻找树皮伤口补充营养。
栗山天牛成虫交配的高峰期在下午4时至晚上9时,其余时间也会交配,但比例较少。交配过程中,雌成虫驮着雄成虫爬行、取食,甚至产卵。
2.3.2栗山天牛成虫交配持续时间、输精持续时间及交尾间歇时间
栗山天牛成虫交配持续时间最长可达2 h,最短只有几分钟,平均约40 min。交配过程输入精子的频次差异较大,最少的只有1次,最多的有12次,平均约5.5次。输入精子的频次与交配持续时间呈正相关关系,即交配持续的时间越长,输入精子的次数越多。关系方程y= 0.0918x+1.6821,F检验结果表明,两者之间的相关性达到了极显著水平(F=96.7128,df=1,16,P=0.0001)(图4)。
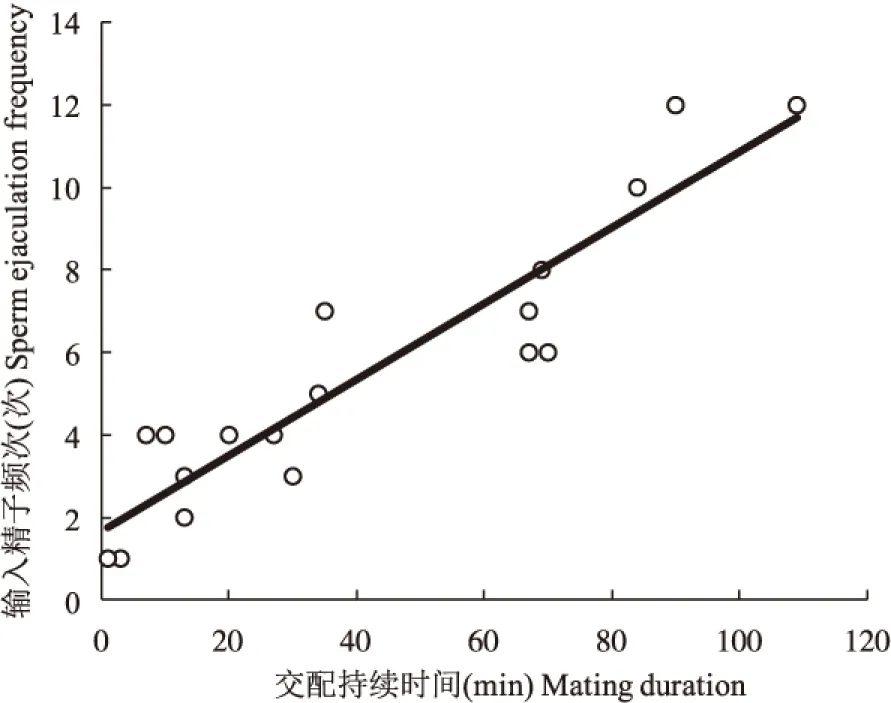
图4 栗山天牛雄成虫输入精子频次与交配持续时间的关系Fig.4 The relationship between sperm ejaculation frequency of Massicus raddei male and its mating duration
栗山天牛雄成虫输入精子持续时间与输入频次显著相关(F=29.628,df=11,63,P=0.0001)(图5)。第一次输入精子时持续时间最长,平均约为17 s,第二次持续时间稍短,略为15 s,第3次下降到13 s左右,第4次下降到只有10 s,与前3次差异显著,之后持续时间逐渐下降,到第10次时,已经下降到不足2 s,最短的持续不到1 s。观察过程中,只有2头雄成虫输入精子次数达到了12次,最后一次持续时间略有上升,平均约4 s。
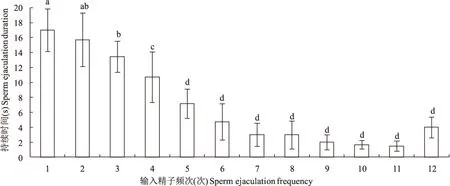
图5 不同频次下栗山天牛雄成虫输入精子的持续时间Fig.5 Thesperm ejaculation duration in different sperm ejaculation frequency of Massicus raddei male
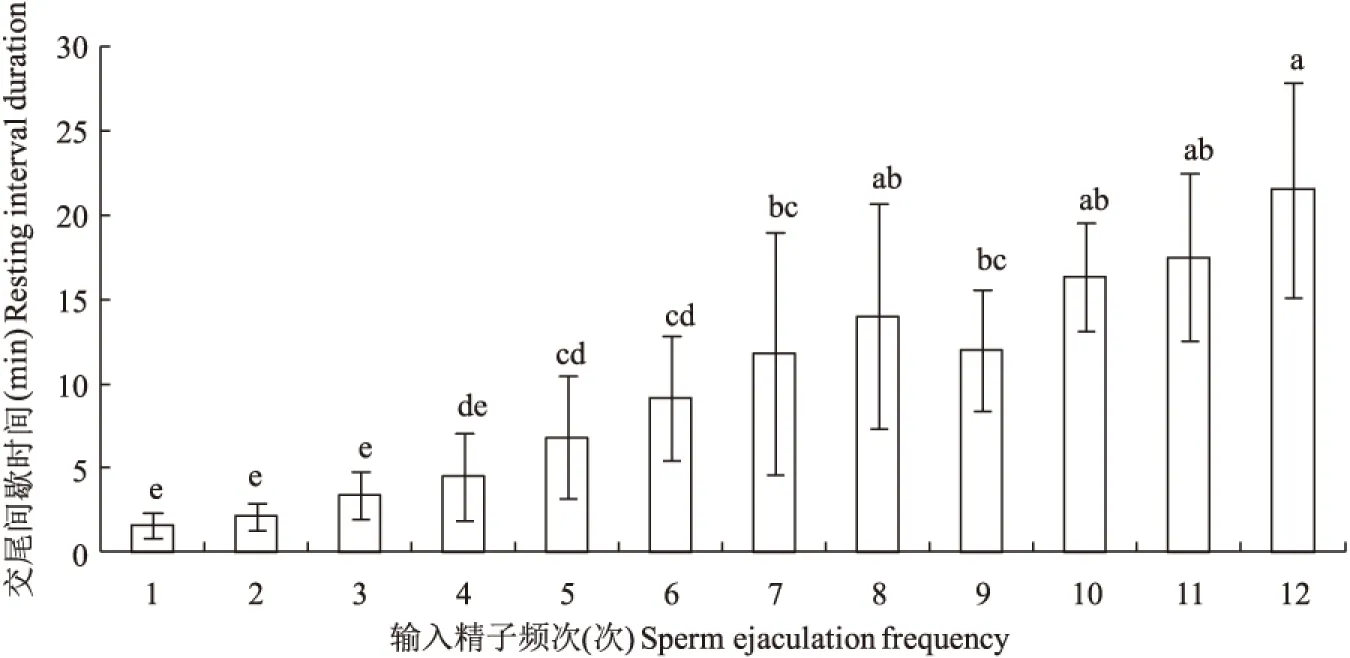
图6 不同输精频次下栗山天牛雄成虫的输精间歇时间Fig.6 The resting interval duration in different sperm ejaculation frequency of Massicus raddei male
栗山天牛成虫交尾间歇时间随着输精频次的增加而增大,不同频次之间差异显著(F=12.304,df=11,61,P=0.0001)(图6)。第一次插入输精之后的间歇时间最短,只有不到 2 min,之后间歇时间逐渐增加,前3次插入输精间歇时间均较短,差异不显著。第5次插入输精之后,平均间歇时间超过5 min,第7~11次增加明显,平均间歇时间都超过10 min,第12次平均间歇时间超过了20 min。这说明,栗山天牛雄成虫在交配前期,每次插入输精之后,需要休息的时间较短即可以再次进行插入输精,而后期需要相对更长的时间。
2.4 栗山天牛雌成虫的产卵行为
栗山天牛雌成虫主要在栎树树干上产卵。产卵前,雌成虫一边在树干上爬行,一边用产卵器在树皮上试探,试探树皮裂缝的深度,找到合适的产卵缝隙之后,即将产卵器插入树皮裂缝中产卵。卵粒多数为单产,亦有少部分是两个产在一起。卵的大头朝内,小头朝外,幼虫孵化后即钻入树皮取食危害。产卵结束后,雌成虫会分泌一种乳白色胶状物将卵粒覆盖,之后胶状物逐渐氧化变成绿色同时变硬。
3 结论与讨论
在绝大多数的昆虫种类中,卵受精都必须通过雌雄个体的交配行为来完成,因此,交配行为在种群的延续中具有重要的作用,对交配行为的研究也成为昆虫行为学的热点(Fauziahetal., 1987; Kimetal., 1992; 李德家等,1999)。目前,国内有关天牛成虫交配行为的研究报道不多,研究比较详细的只有光肩星天牛Anoplophoraglabripennis和松褐天牛Monochamusalternatus(贺萍等,1993;杨洪等,2007)。贺萍和黄竞芳(1993)研究证实光肩星天牛存在有外性信息素,不过只在短距离内起作用。有交配欲望的栗山天牛雄成虫在找到雌成虫之前,会不停晃动触角,碰到天牛成虫之后,触角会在第一时间接触,因此,判断栗山天牛也有外性信息素(魏建荣等,2013),但这种信息素在多大的范围内起作用,还需要深入研究。杨洪等(2007)研究了松褐天牛的交配行为,摸清了松褐天牛的交配时间、次数和过程,为松褐天牛的防治提供了依据。栗山天牛的交配过程和松褐天牛比较相似,交配前没有明显的求偶行为,交配过程持续时间长短不一,交配过后部分有配后保护的行为。
多数昆虫都表现有多次交配的行为,栗山天牛亦不例外,雌成虫一生中会与不同雄成虫进行多次交配,而雄成虫也会与不同雌成虫多次交配(Arnqvist & Nilsson, 2000; 长有得和康乐,2002)。和多数天牛一样,栗山天牛并没有固定的配偶关系。因此,雄成虫和雌成虫都面临比较大的自然选择和性选择压力。雄性和雌性在面对这些压力时,所采取的策略是不一样的(长有得和康乐,2002)。具体到栗山天牛,雄成虫会在交配之前对雌成虫进行一系列的评估,通过触角的接触以及试探性的插入来评估雌成虫的发育状态和已交配的情况,试验过程中观察到相当一部分雄成虫在经过前期评估之后选择了离开雌成虫,亦有一部分雄成虫会与雌成虫交配,但持续时间较短,只有少量雄成虫会与同一雌成虫进行长时间的交配,交配完成之后还有比较明显的配后保护过程。雌成虫在选择雄成虫的初期是比较被动的,交配发生之后一段时间,雌成虫的反应差异很大,有相当一部分雌成虫会挣脱从而导致交配中断。关于雌成虫多次交配的适应性意义,许多学者进行了深入的研究并提出了许多假说(Arnqvist & Nilsson, 2000; 长有得和康乐,2002)。面对激烈的雄性竞争,天牛雌成虫必然要进行多次交配,雌成虫通过多次交配能补充受精囊内精子的消耗,并能提高雌成虫的产卵量和卵孵化率(Fukaya, 1999; 杨洪等,2006)。此外,经过多次交配,雌成虫有足够的精子进行筛选,从而降低产生劣质后代的风险。
MacArthur和Pianka在1966年就提出用经济学观点来研究动物的行为,并发展了最适选择理论。之后,McFarland(1977)在此基础上发展了后来的进化稳定性策略(ESS),这种策略具有不可侵蚀性,这也是动物应对复杂多变的环境采取的有效策略。对于栗山天牛而言,吸食寄主树干伤口处流出的汁液比啃食寄主树皮要容易的多,而且树干流出汁液的伤口是比较少的,这就能引起天牛成虫的聚集,从而大大增加雌雄成虫接触的机会。这是栗山天牛明显不同于其他天牛的地方。这种取食对策可以看做是栗山天牛长期进化而采取的一种稳定性对策。另外,雄成虫在寻找配偶中占有主导地位,它们比雌成虫爬行更快、飞翔更多,一旦找到雌成虫,就积极寻求交配,而大多数雌成虫接受雄成虫交配是比较被动的,推测它们可能通过筛选雄成虫的精子等其他策略来争取产生更优质的后代。进化稳定性策略是比较复杂的,要想弄清天牛行为的各种进化稳定性策略,还需要详细和定量的研究。
