真菌防治蝗虫研究进展
2021-05-11农向群闫多子王广君涂雄兵张泽华
农向群,闫多子,蔡 霓,王广君,涂雄兵,张泽华
(中国农业科学院植物保护研究所/植物病虫害生物学国家重点实验室,北京 100193)
蝗灾是世界经常性的自然灾害。据联合国粮农组织FAO统计,全球常年发生蝗虫的面积达4680万km2,威胁到100多个国家或地区,时常在非洲和亚洲造成严重灾害。几百年来,在我国以及澳洲、非洲都曾暴发多次大规模蝗灾,造成作物绝产、草地荒芜。2002—2013年我国天然草地年均危害面积2099万hm2,农区飞蝗年均成灾面积约320万hm2,对我国的农牧业生产造成严重破坏[1]。2004—2014年沙漠蝗Schistocerca gregaria在非洲西北部接连5次暴发,生态系统破坏加剧。2019年再次在肯尼亚、埃塞俄比亚等国暴发,短短数月内蝗虫数量激增6400万倍,并扩散迁移。2020年初,已扩张到其西南各国,并在当地又形成新生代种群,同时向东北一路越过了红海、沙特阿拉伯、伊朗,进入了巴基斯坦和印度西北地区,正发育成熟进入产卵繁殖季节[2],如果遭遇强劲的印度洋西南季风,蝗虫将有机会飞跃横断山脉,对我国构成潜在威胁,形势十分严峻。
全世界的蝗虫已知有1万种以上,其中对农、林、牧业可造成为害的蝗虫约300种,全球除南极洲、欧亚大陆北纬 55°以北地区外均可发生蝗害[3]。沙漠蝗被认为是世界上最具破坏力的迁徙性害虫之一,主要分布在北非、中东和西南亚的干旱和半干旱地区,其飞行能力强、繁殖快、食量大,可聚集形成巨大蝗群。每天可以随风飞行150 km,存活时间为3个月左右[4,5],一只雌蝗可产卵大约300粒[6],1平方公里规模的蝗群1 d的进食量相当于3.5万人1 d的进食[7],是非洲、亚洲热带荒漠地区的河谷、绿洲上的重大农业害虫。
非洲中部、中南部、萨赫勒地区是蝗灾频发地,常见蝗虫有非洲飞蝗Locusta migratoria migratorioides、红蝗Nommadacris septemfaciata、树蝗Anacridium melanorhodonI;在亚洲中部、东南部地区,如中国、越南、哈萨克斯坦的蝗虫优势种有15~20种,包括东亚飞蝗Locusta migratoria manilensis、亚洲飞蝗Locusta migratoria migratoria、亚洲小车蝗Oedaleus asiaticus、意大利蝗Calliptamus italicus等;在澳大利亚昆士兰、南澳大利亚、新南威尔士、维多利亚等5个州,常见蝗虫有几十种,有澳洲疫蝗Chortoicetes terminifera、飞蝗 Locusta migratoria、红蝗等;在北美洲和墨西哥北部有美洲沙漠蝗 Schistocerca americana、血黑蝗Melanoplus sanguinipes、edaleonotus enigma等;在欧洲有摩洛哥蝗 Dociostaurus maroccanus、白边雏蝗Chorthippus albomarginatus、意大利蝗等[8,9]。
在自然条件下,如果不加以人工防控,暴发的蝗害可迅速扩张,直至因食物绝尽或蝗虫病原在适宜条件下造成流行病才能最终控制。在20世纪40~70年代,化学农药兴起并应用到蝗虫防治中,在蝗虫大发生时能够快速杀灭种群。然而,蝗虫产卵滋生地通常较为偏远,发生面积巨大,成虫可远程迁移,施药措施难以全面跟进。大量施用化学农药带来了杀伤天敌、破坏生态等问题,加上气候异常变化,致使蝗灾频发甚至加剧。在寻求环境友好的可持续防蝗新策略、新技术研究中,利用昆虫病原真菌的生物防治受到重视并显现出明显优势。近30多年来,利用真菌防蝗的研究进展较快,并在蝗虫灾害防控中发挥了重要作用。
1 真菌杀虫机理
昆虫和真菌在漫长的进化过程中,形成了多样化的相互关系,包括致病、寄生、腐生、共生、携播、噬菌、竞争、捕食等[10]。昆虫病原真菌(Entomopathogenic fungi)是指能够侵入健康昆虫(或其他节肢动物)寄主,在其体内增殖并引起早期生理病变,影响发育、生殖和导致死亡的一类真菌。这是功能性的类别而非系统发生学的分类,这类真菌主要体现了上述的致病和寄生关系。
真菌对昆虫的致病性是二者长期共同进化的结果。真菌对昆虫感染致病过程即是与寄主相互作用,包括突破昆虫寄主体表屏障和克服寄主免疫系统的过程,其进程大致分为三个阶段,①侵入阶段:分生孢子在黏液和粘附蛋白的帮助下附着于昆虫体表,并萌发和形成侵染结构,穿透表皮;②与寄主互作早期阶段:侵入昆虫血腔的芽管通过断裂增殖形成虫菌体定居下来,同时影响昆虫的正常生理代谢,激活寄主先天免疫应答,包括体液和细胞防御;③病害形成阶段:入侵的真菌逐步战胜昆虫的免疫系统,在寄主体内增殖、产生毒素并致死昆虫(图1)。
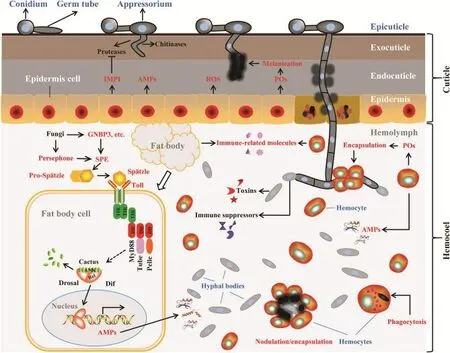
图1 昆虫病原真菌与昆虫免疫反应之间相互作用的示意图[11]Fig. 1 Schematic of interactions between entomopathogenic fungi and the insect immune responses[11]
1.1 突破寄主体壁屏障
昆虫病原真菌通常直接穿透外表皮来感染寄主昆虫。体壁是昆虫抵抗感染的第一道屏障。真菌分生孢子首先特异性或非特异性地吸附于昆虫体壁,萌发后可分泌孢外蛋白酶及粘附因子使之牢固地附着在昆虫体壁上[12,13]。研究发现了一些负责疏水性、粘附性和毒力的基因,例如,金龟子绿僵菌Metahizium anisopliae疏水基因 ssgA编码的疏水蛋白可以帮助孢子附着于昆虫体表[14]。金龟子绿僵菌在侵染寄主表皮时表达粘附蛋白 Mad1,促进真菌孢子稳定地吸附于寄主体表,如果缺失 Mad1基因,分生孢子的粘附能力与毒力都显著降低[15]。另外,其他细胞壁蛋白如CWP10和CP15,它们参与了耐热和抗氧化胁迫反应,在粘附和应激反应中发挥关键作用[16]。
真菌孢子在昆虫表皮上萌发后,形成附着胞。附着胞经过一段时间的膨大并形成侵入钉,借助机械压力及蛋白酶、几丁质酶、脂酶等水解酶的多重作用来降解昆虫表皮,穿透寄主体壁[17]。蛋白酶主要有两大类,一是对底物短肽 Sue-Ala-Ala-Pro-Phe-Nz具有活性的类枯草杆菌蛋白酶 Prl,另一类是对短肽Ben-Phe-Val-Arg-NA有活性的胰蛋白酶Pr2[18],它们的同工酶氨基酸序列中具有相同的活性区域,但其他部分序列变化较大[19,20]。类枯草杆菌蛋白酶Pr1能溶解昆虫表皮中的蛋白质成分,有助于菌丝的入侵,同时为菌丝的生长提供营养[21]。Pr1类蛋白酶在昆虫病原真菌侵入寄主和致病过程中是重要毒力因子,pr1a基因被破坏的金龟子绿僵菌菌株毒力显著下降,而pr1a高水平表达的菌株杀虫毒力明显提高,杀虫时间缩短25%[22];昆虫血淋巴中高浓度的Prla蛋白酶可引起昆虫酚氧化酶的过度表达,导致昆虫中毒死亡[23]。
几丁质是昆虫体壁屏障的主要成分,真菌几丁质酶是真菌突破寄主体壁的另一个重要的因子。用昆虫生长调节剂Dimilin处理烟草夜蛾Manduca sexta幼虫,阻止其几丁质合成,结果试虫体壁很容易被金龟子绿僵菌降解。几丁质酶有多种类型,包括外切几丁质酶和内切几丁质酶,根据等电点分为酸性和碱性几丁质酶等。对多种几丁质同工酶作用研究表明,DGIDVDWEYP对真菌几丁质酶具有高度保守性,推定是底物结合酶(Processing enzymes)的作用位点[24]。几丁质酶是诱导酶,侵染寄主体壁过程中在稍后期才产生,需要蛋白酶作用使几丁质露出后再诱导产生[20]。
病原真菌如果突破体壁失败,原因可能是体壁的某些成分抑制了真菌孢子的萌发,或是体壁的分子筛功能阻挡了真菌分泌物的渗透,还有昆虫表皮自身的推陈出新,排除了病原真菌及其分泌物。另外,昆虫已选择进化了抵抗病原的特异蛋白酶的抑制剂,如IMPI(Insect metalloprotease inhibitor)[25],而真菌分泌的酶种类虽多但分泌量有限,从而影响了其对表皮的降解程度[26]。最新的转录组分析表明,在病原真菌附着寄主体表和侵入的早期(12~24 h)阶段,寄主因识别到病原相关分子模式(Pathogen-associated molecular patterns)激活了天然免疫系统,出现了信号转导基因(Myd88、Relish家族、Pelle等)以及下游一些氧化酶(PPO)、溶菌酶和防御素等基因的显著差异表达,产生对入侵病原的防御,这可能也是限制病原真菌突破寄主体壁的重要因素之一[27]。
群居昆虫还可以通过群体行为回避达到对真菌感染的有效防御。如白蚁和蚂蚁,可利用嗅觉线索从远处探测到被污染个体上的有毒真菌。大型白蚁可通过真菌释放的挥发性有机化合物区分真菌的毒性高低,并采取相应的防御行为[28]。对病原真菌的行为防御还包括自我修饰以及摄入或产生具有抗病性的化合物,如一些群居昆虫通过传播唾液腺或表皮后胸腺产生的抗真菌分泌物(如甲酸、抗菌肽和蛋白样液体),以保护自身及巢内同伴[29]。还有研究表明沙漠蝗可通过晒太阳来提高体温,以对抗绿僵菌感染[30]。当种群密度高时,昆虫宿主通过预防性免疫的增强对抗真菌,如群居性蝗虫通过预防性免疫的增强来降低对真菌的易感性[31]。
1.2 抑制寄主体液免疫
昆虫免疫系统由体液免疫(humoral immunity)和细胞免疫(cellular immunity)组成。体液免疫是指昆虫体内本身存在或受诱导由脂肪体和血细胞分泌的免疫因子,包括抗菌肽、抗菌蛋白、凝集素、溶菌酶、酚氧化酶等物质,对病原微生物产生免疫作用。昆虫体液免疫主要有4条信号通路,Toll、IMD通路是昆虫抵御病原真菌的核心通路,而JAK/STAT信号通路则主要对病毒的侵染作出免疫应答,JNK信号通路则是昆虫通过介导细胞凋亡抵御病毒侵扰[32]。病原真菌的侵入,激发了昆虫体液免疫功能。Toll、IMD通路包含多种蛋白因子介导的黑化反应、抗菌肽、溶菌酶、活性氧(ROS)、凝集素等产生的复杂免疫应答反应[33]。Toll的激活与反应如图1(左侧)所示,革兰氏阴性结合蛋白3(GNBP3)是β-1,3-葡聚糖识别蛋白家族(βGRP3)的成员,能识别并结合真菌细胞成分,级联反应启动Toll信号通路。Persephone和Späetzle加工酶(SPE)引起半胱氨酸结细胞因子(Cysteine knot cytokine)Späetzle(Spz)的裂解。当蛋白水解切割的配体Spz与受体结合时,Toll受体被激活。然后将信号传递到由MyD88、Tube和Pelle组成的细胞内信号转导级联中,最终通过有效的NF-κB转录因子Dorsal和Dif引起AMPs的表达。
酚氧化酶(Pheonloxidase,PO)及其相关酶类是重要的免疫激活机制。该酶平时以无活性的酚氧化酶原(Porpheonloxidase,PPO)形式存在,当外来生物入侵时,可通过特异性丝氨酸蛋白酶的级联反应(PPO级联)而活化,产生黑化反应,清除病原体[34,35]。抗菌肽(AMPs)在脂肪体、表皮细胞和血细胞中产生。目前共发现了1500多种抗菌肽,近700种抗真菌肽。昆虫受到微生物感染或损伤后,脂肪体和血细胞会迅速合成一些抗菌肽,分泌到血淋巴中消灭入侵的病原物。例如,在大蜡螟Galleria mellonella幼虫体内发现一种识别绿僵菌的抗真菌肽gallerimycin[36]。溶菌酶是由昆虫的脂肪体分泌并释放到血淋巴中的一种免疫因子,其通过攻击肽聚糖或水解微生物细胞壁上的 N-乙酰胞壁酸的糖苷位点,进入细胞内并破坏其结构,最终杀死病原物[26]。从经免疫诱导过的大蜡螟、家蚕Bombyx mori L.和甘薯天蛾Agrius convolvuli中纯化得到溶菌酶,对真菌和革兰氏阳性细菌均具有显著的抑制效果[37]。凝集素(Lectin)也是一种糖结合蛋白,通过改变真菌膜结构或渗透性直接抑制真菌生长,也可以通过连接到真菌细胞壁的糖蛋白上间接地影响菌丝的生长。
真菌通过多种方式躲避或抑制寄主免疫。在穿过体壁后初期,真菌在寄主血腔内以原生质体、类似原生质体和囊胞子等酵母状形式存在,以短菌丝出芽方式繁殖。这种生长形式一方面有助于在体血腔中的分散与群集,通过增加表面积获取更多的营养物质,快速繁殖,以数量优势“逃避”宿主有限细胞的免疫反应[38];另一方面,这类细胞表面的细胞壁多糖很少,可最大限度不激发或躲避寄主的免疫反应[39]。病原真菌通过分泌各种毒力因子对抗寄主体液免疫和导致寄主病变,如将金龟子绿僵菌破坏素Destruxin A注射到果蝇体内,可导致各类抗菌肽的表达量都下调[40]。金龟子绿僵菌在蝗虫血淋巴中分泌含锌的羧肽酶MeCPA和酸性磷酸酯酶,前者可消化寄主体内蛋白作为自己的营养来源,后者则改变其寄主血淋巴中与信号转导相关蛋白的磷酸化状态,进而干涉寄主体内免疫活动[41]。绿僵菌分泌的海藻糖酶能够有效降解昆虫血淋巴中的海藻糖为葡萄糖,获取能量吸收,破坏昆虫的血液生理,从而对昆虫起着致命的作用[42]。真菌分泌的氨,作为一种毒力因子,改变血淋巴 pH,破坏昆虫组织和干扰昆虫的免疫系统,间接减弱寄主免疫反应[43],同时真菌通过酶的代谢产物改变环境pH,使各种酶在最适宜的pH条件下产生并发挥有效功能[44]。
1.3 躲避寄主细胞免疫
昆虫细胞免疫是由血细胞介导,完成对病原物或异物的吞噬、包囊和结节等一系列免疫应答反应。免疫细胞主要有血淋巴中的浆细胞和粒细胞,浆细胞具有吞噬功能,而粒细胞可包囊大分子物质并去核化形成结节[45]。
当把绿僵菌孢子注射到金龟子幼虫体内,血淋巴中会形成典型的包囊体,孢子在包囊体内被黑化,丧失萌发能力[46]。当昆虫受伤时,血细胞会在伤口处迅速凝结,阻止血淋巴流失,防止病原微生物侵入[38]。细胞免疫通常与体液免疫协同作用,图1中(右侧)AMPs由脂肪、表皮细胞和血细胞产生,血淋巴中血细胞识别真菌,并产生吞噬作用和包囊反应。
病原菌主要通过3个方面逃避昆虫的细胞免疫:(1)杀死血细胞及抑制细胞吞噬和结节形成;(2)利用体表的脂类成分来逃避昆虫血细胞的免疫识别和包裹;(3)分泌毒力因子,金龟子绿僵菌侵入烟草天蛾血腔后,大量表达MCL1蛋白覆盖于菌体表面,以此来逃避烟草天蛾幼虫血细胞的免疫识别[47]。莱氏野村菌Nomuraea rileyi感染棉铃虫Helicoverpa armigera后,在寄主血淋巴中分泌毒蛋白或次生代谢物等毒力因子,通过破坏血细胞的细胞骨架,抑制其延展能力,阻遏了血细胞的免疫反应,从而能够成功地在棉铃虫血腔中生长和繁殖[48]。
病原真菌分泌的毒素可抑制寄主细胞免疫反应。已知的毒素主要有白僵菌素(beauverolides)、环孢素(cyclosporin)和破坏素(destruxin)等。已知破坏素由 6个氨基酸残基通过酯键首尾相连形成环缩肽,环外加上不同类型羟基酸和氨基酸残基,至少有19种类型[49]。绿僵菌通过释放破坏素抑制沙漠蝗和美洲大蠊Periplaneta americana Linnaeus的酚氧化酶活性,破坏其参与体液免疫功能[50],而白僵菌感染甜菜夜蛾后,能够抑制寄主酚氧化酶的激活,使酚氧化酶的活性和分布明显降低[51]。研究发现,破坏素通过改变寄主细胞的Ca2+平衡和细胞内高分子蛋白的磷酸化来抑制吞噬细胞的粘附和扩散能力,并影响浆细胞的细胞骨架形成[52],破坏素还可引起昆虫细胞的程序性死亡,从而减弱寄主的免疫能力[53]。结合表型反应,破坏素可激活肌肉细胞的Ca2+通道,引起昆虫痉挛后麻痹而死[54];有些破坏素可影响寄主蜕皮激素合成,从而影响昆虫的变态,如抑制烟草夜蛾新生4龄幼虫向5龄的转化进程[55]。生理和病理的多重作用导致寄主机体病变直至死亡。
2 防蝗菌株选育
2.1 杀虫真菌种类
在自然条件下病死的昆虫中,有约60%起因于真菌感染罹病所致。全世界已报道的昆虫病原真菌有约1000种。在早期,一些昆虫病原真菌常常只见其无性循环的过程形态,如菌丝、分生孢子等,但未见或未知其有性循环过程,因此暂且归为半知真菌类(门或纲)。随着研究的深入和分子系统学的发展,现在半知真菌类的属种多数都能够鉴定到相应的有性型,重新归入分类系统的适当地位,“半知菌(Imperfecti fungi)”、“不完全菌纲(Deuteromycetes)”等名称已经被放弃[56,57]。按照目前的系统发生学,真菌类主要分7门和4个单列亚门[58,59](图2),其中子囊菌门Ascomycota和担子菌门Basidiomycota为高级进化类群;接合孢菌(Zygosporic fungi)和游动孢子菌(Zoosporic fungi)进化相对初级,且不是单系进化,形态学上显示多样化;而微孢子菌门Microsporidia为低等真菌,最初认为是散发的胞内寄生物,归为原生动物 Protozoa,现根据多基因座测序和基因组学的证据表明应属于真菌类[60,61]。

图2 真菌门类和大约物种数关系图示[59]Fig. 2 Diagrammatic representation of relationships of fungal taxa and approximate number of species in each phylum or subphylum[59]
昆虫病原真菌多属于虫霉亚门的虫霉目Entomophthorales和子囊菌门的肉座菌目Hypocreales,还有一些属于壶菌门 Chytridiomycota、球囊菌门Glomeromycota、担子菌门Basidiomycota和微孢子菌门 Microsporidia[59]。一般虫霉目真菌的寄主比较专一,而肉座菌目真菌多数有较宽泛的寄主范围(表1)。在传统的形态学分类系统中,有的物种可能是复合种或含有不同种群类型,例如,最为常见和研究最多的球孢白僵菌Beauveria bassiana和金龟子绿僵菌已积累众多菌株,显示具有高度的遗传多样性,近年分子系统研究已经可以较好地区分和细化绿僵菌属和白僵菌属内物种和类型[62,63]。即使同一物种,不同株系在寄主范围、侵染水平、萌发率等方面也有所差异,表现了株系种群对不同环境的适应性。总体上。昆虫病原真菌寄主范围涵盖了昆虫纲中的大多数目类,至少有800种。最常见的绿僵菌属和白僵菌属寄主分别有200多种和700多种,包括鳞翅目Lepidoptera、鞘翅目Coleoptera、直翅目Orthoptera、同翅目Homoptera等中的多种重要农林害虫[64]。

表1 重要昆虫病原真菌属种的分类地位和寄主Table 1 Taxonomic status and hosts of important genera and species in entomogenous fungi

续表1
虽然蝗虫类群庞大,直翅目蝗总科共有8科含1万余种,中国已知700余种,但丰富的病原真菌种类为蝗虫生物防治提供了广泛资源。在虫霉亚门的虫霉目和子囊菌门的肉座菌目中都发现了蝗虫的病原真菌。虫霉目研究较多的是蝗噬虫霉复合体 Entomophaga grylli complex,其至少包括 4种致病型,I型 E.macleodii主要寄生斑翅蝗亚科Oedipodinae的蝗虫;II型E. calopteni主要寄生黑蝗亚科Melanoplinae的蝗虫;III型E. praxibuli两亚科均可寄生;IV型E. asiatica是从日本的一种蝗虫体分离得到的。用分子标记可将它们区分鉴别,我国主要有蝗噬虫霉I型。噬虫霉类真菌可采用接种虫体培养,对人工培养基要求较高,如需要牛奶、蛋黄或加牛血清等,目前还难以大量繁殖利用。肉座菌目广泛存在于自然界,是蝗虫的主要病原真菌,包括绿僵菌 Metarhizium属的金龟子绿僵菌、蝗绿僵菌 M. acridum、黄绿绿僵菌 M.flavoviride,白僵菌属Beauveria的球孢白僵菌、曲霉属Aspergillus的黄曲霉A. flavus等。绿僵菌和白僵菌是蝗虫的主要寄生性病原真菌,种群类型多、致病力强且易于人工培养,尤其绿僵菌具有对蝗虫的致病性优势,成为国内外研究与开发利用的热点,并不断取得可喜进展。
2.2 菌株筛选评价
通常可以从病死虫体上或从土壤中分离获得菌株,在实验室对靶标害虫和其他昆虫接种进行生物测定,评估菌株的杀虫毒力和寄主范围,同时接种在培养基上,评估菌株的碳氮源等营养需求、适宜温度、pH范围、生长速率和产孢量等参数。在著名的撒哈拉地区蝗虫生物防治国际合作LUBILOSA项目(1989—2002)中,英联邦国际生物防治研究所联合国际热带农业研究所等多国团队,从西非、也门、马达加斯加等地区的土壤或病蝗中分离出300多个对蝗虫有效感染的真菌菌株,进行了大规模生物测定以筛选高毒菌株,还评估了菌株遗传稳定性、产孢能力、孢子稳定性、对紫外线和高温的抗性、田间有效性、对非靶标生物的安全性、对环境的影响和循环性等特性,确定了高效的金龟子绿僵菌IMI330189等菌株以及球孢白僵菌、小团孢属Sorosporella sp.的几个优良的潜力菌株。菌株IMI330189最初鉴定为黄绿绿僵菌,后来根据Driver等[65]的分子系统订正为金龟子绿僵菌蝗变种M. anisopliae var. acridum,后来在Bischoff等[62]的多基因位点分类中将其独立为蝗绿僵菌M. acridum。该菌株为项目的顺利开展和后来产品的研发商业化奠定了基础。
在澳大利亚,针对当地灾害性蝗虫Chortoicetes terminifera、Phaulacridium vittatum、Locusta migratoria、Austracris guttulosa、Valanga irregularis等进行了生物测定筛选,获得了高毒力的金龟子绿僵菌FI-985菌株[66,67]。在国内,1993年中国农业科学院生物防治研究所通过国际合作,自英联邦国际生物防治研究所(IIBC)引进了防蝗系列菌株,并对东亚飞蝗进行了毒力测定,显示菌株Arsef 324(后来确定为金龟子绿僵菌)和IMI330189有较高毒力[68];同时该研究所科研团队也从本土菌株中筛选到对东亚飞蝗、亚洲飞蝗、意大利蝗、亚洲小车蝗等高毒力菌株。其他研究者也先后报道了不同地区采集分离的高毒力的防蝗菌株[69,70],以及利用温度梯度热激胁迫、诱导提升耐热性和抗UV-B的防蝗优良菌株[71]。
2.3 菌株遗传改良
为了提高菌株杀虫毒力和抗逆生存能力,可以利用生物技术进行菌株改良。真菌遗传改良方法主要利用有性(或准性)循环、原生质体融合、辐射诱变和基因工程途径来实现。真菌有性循环和准性生殖(parasexual reproduction)是两个异性生殖细胞或异核菌丝体结合,通过染色体交换产生遗传重组的个体。真菌繁殖过程中普遍存在异核现象和准性生殖。利用准性生殖,可以将两种基因型不同的具有亲和性的个体进行体细胞杂交(准性杂交),得到预期新性状的个体。用绿僵菌野生型菌株与营养缺陷突变株作材料证实了准性杂交现象,并获得二倍体重组体且具有较高的产孢率和较强的致病性[72]。
辐射诱变除了常规的紫外照射处理,航天搭载能够使生物种质在太空环境的强辐射、超速度、高真空、微重力等因素的复合作用下,发生大幅度的生理变异和遗传变异,因而成为高效率育种手段。从航天搭载后的绿僵菌突变株中检测到的形态、生长速度、产孢量、致病力等多项生物学特性发生了不同程度的变异,变异率高,变异趋向有正负双向性,其中一些突变株的生长特性和致病力优于原始菌株[73]。
原生质体融合是指不依靠生物体本身所具有的性别能力,而是人为地利用酶降解去除细胞壁,在渗透压稳定和pH适宜条件下,通过电脉冲或化学助溶剂使细胞融合,细胞壁再生后获得新种细胞。这种方法可以克服菌丝不相容性,甚至能跨越种、属间的界限,实现远缘杂交。以原生质体进行紫外照射处理可获得较高的诱变效率。以白僵菌和绿僵菌为例,将0.5%纤维素酶和0.5%蜗牛酶的混合酶液、0.6 mol/L NH4Cl或KCl为渗透压稳定剂;在pH 6.5时30 ℃处理菌丝,可从1 g湿菌丝中获得5×107个以上的原生质体[74,75]。以球孢白僵菌和金龟子绿僵菌为出发材料,进行相同物种不同菌株之间、不同属物种之间的原生质体融合试验,以 β-葡萄糖醛酸苷酶和 KCl作裂解酶和渗透稳定剂,以聚乙二醇(PEG)为融合剂,融合再生率达到 0.84%~1.08%,获得了菌丝生长快和产孢量高的融合子,并发现由白僵菌和绿僵菌原生质体融合的重组菌株中,胞外蛋白酶Pr1和Pr2的活性比亲本增加了两倍[76]。对蛹虫草Cordceps militaris原生质体进行紫外诱变处理,筛选出两株虫草多糖和子实体产量都明显高于原始菌株的优良菌株[77]。
遗传工程改良菌株是近年的主要方向,至今已发现一些昆虫病原真菌的毒力基因、抗逆基因及其调控相关基因,外源基因的导入方法日臻成熟。向丝状真菌中转化外源 DNA的方法有农杆菌介导,或制备原生质体并借助PGE-CaCl2促进融合,或借助电脉冲来完成转化。虫生真菌抗性育种中,第一个利用基因转移技术的成功范例是构建重组质粒 pBENA3,将构巢曲霉 Aspergillus nidulans的抗苯菌类基因BenA3导入金龟子绿僵菌的原生质体,转化率为9个转化子/50 μg DNA/2×106原生质体,转化菌株对苯菌灵的抗性比原菌株高 10倍[78]。基因工程对昆虫病原真菌遗传改造的另一个突破性进展是试验选择了MCL1作为启动子,利用绿色荧光报告基因,首次将一种北非蝎的昆虫专化性神经毒素多肽AaIT(中性蝎毒)反编码成真菌基因,构建了带有抗性标记的替换质粒,经原生质体转化后,筛选抗性菌落,在受控操纵子的调节下使金龟子绿僵菌在侵染至昆虫体内时高水平表达。转化的工程菌株显著提高了杀虫速率,对烟草天蛾的毒力提高22倍,对埃及伊蚊Aedes aegypti的毒力提高9倍;与此同时,还显著减少了昆虫的活动性,降低了昆虫的取食率[79]。还有,把来自一种钳蝎Buthotus judaicus的杀虫蝎毒素(Bj alpha IT)基因整合到蝗绿僵菌中,增强了对东亚飞蝗的毒力,使 LC50降低了 18.2倍,局部接种和注射工程菌的LT50分别减少了28.1%和30.4%[80]。近年研究陆续发现了更多的昆虫病原真菌毒力相关基因、抗逆生长相关基因和产孢调控基因等,都可作为菌株遗传改良的好材料。将酯酶基因(MestI)从通常广谱寄主的罗伯氏绿僵菌M. robertsii转移到特定的蝗绿僵菌,扩大了宿主范围[81]。如黑色素酶(melanin)在金龟子绿僵菌中的表达使得菌株对紫外线和高温的抵抗力增强[82];将高效的古细菌光解酶(photolyase)基因整合到罗伯氏绿僵菌基因组中,可以提高真菌的光修复能力,从而提高对阳光的抵抗力[83];黄绿绿僵菌菌株用于防治巴西蚱蜢Rhammatocerus schistocercoides,转化了苯菌灵抗性基因(beta-tubulin gene)后,高抗转化子对苯菌灵抗性提高了 30倍,在非选择性培养基上连续转代 20次后仍保持抗性稳定,而且一些糜蛋白酶(Chymoelastase,Pr1)分泌量比野生型更大,分生孢子萌发率更高,在生产和应用上更具潜力[84]。
3 规模化生产与应用
在获得对蝗虫高致病力菌株的基础上,还需要根据菌株生长特性,建立大量培养或规模化生产的方法,以获得足够的批量产品。
3.1 规模化生产
3.1.1 液固两相发酵工艺 大多数昆虫病原真菌具有由分生孢子-萌发-菌丝体生长-产生分生孢子的无性繁殖过程,此过程增殖快,分生孢子是侵染昆虫的基本单元。因此,可利用营养生长和产孢的生活史两个阶段进行人工介入,通过发酵实现菌剂的规模化生产。菌株营养和环境条件是调控生长和产孢的关键因子,有很多关于不同种类真菌或菌株的营养要求和生长环境条件的研究报道。总体来看,多数真菌或菌株均有一定适宜范围的碳源、氮源、无机离子和微量元素,温度、湿度、pH和耗氧性,因此相关研究有较大的共通性。绿僵菌和白僵菌的大量培养和规模生产的方法和基本工艺流程大致相同,可以进行液体深层发酵(Submerged fermentation)、固态发酵(Solid-state fermentation)和液固双相发酵(Liquid-solid two-phase fermentation)。固体发酵相对于液体发酵的优缺点列于表2。液固双相发酵是以大量增殖的菌丝体接种到固体基质,虽延长了流程,但较好地取长补短,发挥液、固相发酵优势。3种类型发酵参数各有不同,最终产物也不同。液体发酵的产物是菌丝体、芽生孢子或液生分生孢子,而固态发酵产物是气生分生孢子。不同的产物形式各具特性,从产物的活性、稳定性和发酵控制条件考虑,气生分生孢子是适合于规模化生产和应用的产品形式。为了获得高产并降低成本,通常需要根据所选用菌株的营养需求和生长条件进行多因子优化,以达到最优效益。

表2 固体发酵相对于液体深层发酵的优缺点[85]Table 2 Advantages and disadvantages of solid state fermentation in comparison to submerged fermentation
在LUBILOSA项目中,对高效防蝗菌株金龟子绿僵菌IMI330189进行了液体深层培养、固体表面培养和液固两相培养研究。深层培养的菌丝体制成干粉,使用时可加水喷雾或喷洒,而在大米或小米表面培养获得分生孢子粉,产量可达100 g/kg固体基料,使用时将孢子粉配置成油悬浮剂进行超低量喷雾[86]。比较几种大量培养球孢白僵菌和拟青霉Paecilomyces fumosoroseus的培养基表明,在松散固体培养基中,大米最适合生长和孢子形成;在液体培养基中,椰子水最适合[87]。对绿僵菌金龟子和黄绿绿僵菌的大量标准化培养条件研究显示,以大米为固相产孢基质,高密度聚乙烯袋为容器,接种分生孢子悬浮液,在25 ℃下孵育15 d,产孢水平分别达到30×108和70×108CFU/g,显著高于接种孢子经液体培养基培养2 d的产物[88]。对金龟子绿僵菌IMI330189的液体发酵动力学进行研究,利用Sigmoid函数构建了该菌株液体发酵过程中的菌体生长和底物消耗的动力学模型,所建模型能够较好地拟合金龟子绿僵菌IMI330189液体发酵过程,并拟合求解出各模型参数,优化参数后,生长速率在发酵第22.8 h达到最大值,为0.084/h;总糖比消耗速率在第9.6 h达到最大值,为0.246/h;总氮比消耗速率在第10.3 h达到最大值,为0.007/h;菌体对总糖的得率系数在39.8 h达到最高,为0.861 g/g[89]。比较2株金龟子绿僵菌菌丝生长及次代产孢的环境因素,经过优化得到最佳培养条件,菌株1为28 ℃、pH 7、装液比为75 mL/250 mL、加入微量元素Mn、全光照培养;菌株2为28 ℃、pH 9、装液比为75 mL/250 mL、加入微量元素Cu,黑暗培养时生长最好、次代产孢量最高。可见不同菌株对光照、通气、微量元素的需求有明显差异[90]。
对于固体培养,基质有多种选择,可直接用谷物粒或疏松载体加谷物粉为主的营养配方,并采用适宜的温度、光照等控制条件,主要考虑最终产孢量和价格成本。在金龟子绿僵菌的三级固体放大培养中,以90%麦麸加10%玉米粉作为发酵培养基,加水量约60%,经过优化,得到最佳产孢培养条件为初始基质pH 6.0、27 ℃、避光,培养6 d孢子产量可达1.21×1010孢子/g干培养基[91]。研究表明,蛭石和稻壳均可作为绿僵菌固相培养的优良载体,玉米粉和麦麸组合是良好的营养源,与不同载体配合的产孢量达到20×108孢子/g以上。液固接种比以8:10最佳;接种后适宜的基质含水量约为55%,产孢量达28.9×108孢子/g;恒温培养时以25 ℃为宜,而采用调温方式是先在25 ℃下培养16 d后转移到21 ℃下再培养5 d,可获得最高产孢量;光照调节对菌株生长和产孢有重要影响,前期暗环境培养14 d,再光照培养7 d的产孢量最高;高通气量利于菌株生长和产孢[92]。为了降低固体基质成本,考察了重复利用大米基质的产孢量,通过响应面方法,确定重要变量为时间和温度,相互作用为温度×糖蜜和温度×时间,重复利用大米基质和新大米同样在25 ℃培养20 d内获得了最高产孢率。鉴于糖蜜具有增加产孢作用但其浓度对产率并不关键,建议设定 5%以降低成本[93]。还有不少类似的研究结果,可构成金龟子绿僵菌规模化的基础平台和技术参数。为了控制产品质量,建议使用基质碳、氮比作为控制指标,确定接种量、温度等培养条件,使用农副产品为潜在基质,加上可循环利用的油形成配方,这可能是一种低成本且对环境可持续的规模化生产技术[94]。
3.1.2 其他培养方式 金龟子绿僵菌的分生孢子的另一种形成方式是微循环产孢。孢子萌发后,当培养条件适宜时,可不经过营养菌丝生长阶段,或营养菌丝生长受到极大限制时,就直接重复产孢[95]。理论上,如果能够控制好微循环产孢条件,将可以缩短发酵周期,降低生产成本,但这种微循环产孢方式只在一些菌株观察到,并与培养成分和培养条件密切相关,且不易控制,产量有限。
微菌核(microsclerotia,MS)是丝状真菌生活史中一种较常见的特化结构,是由菌丝紧密连接交织而形成的黑化、紧实的休眠体,可形态多样、大小不一。菌核外面通常有一层由紧密相连的菌丝顶端构成的皮层,中间菌髓由松散的、宽大的菌丝构成,是微菌核的主要部分,菌髓的细胞中聚集了许多营养物质,使其能够应对不良环境。当环境适宜时,能萌发产生新的营养菌丝或分生孢子等繁殖体。这种结构的抗逆性和稳定性具有十分诱人的开发潜力。第一例报道的昆虫病原真菌微菌核是在金龟子绿僵菌发酵培养中发现的,由此提出了以微菌核作为一种产品的新形式。研究还报道,微菌核只能在液体深层发酵后期才能生成,碳源浓度与碳氮比影响微菌核的产量,碳源浓度为36 g/L、碳氮比在30:1和50:1时MS的产量最大,分别达到2.7×108和2.9×108MS/L,产微菌核方法获得了美国专利[96]。
绿僵菌破坏素的培养条件也被积极探讨,以期开发作为杀虫剂。用响应面法优化了金龟子绿僵菌破坏素A的培养条件,得到1个拟合程度高、误差小的模型,该模型给出的最佳培养条件为:蔗糖22.7 g/L、蛋白胨13.4 g/L和发酵时间9.70 d,预测绿僵菌破坏素A最大质量浓度为6.90 μg/mL,实际测得质量浓度为6.89 μg/mL[97]。此结果为绿僵菌大规模发酵获得破坏素产品提供理论依据,有望使绿僵菌毒素得到广泛的开发和应用。
芽生孢子是通过出芽方式产生的无性孢子,许多真菌在液体发酵中都可以大量产生。在3 L容量的搅拌釜式生物反应器中,通过响应面方法得到了优化的蝗绿僵菌M. acridum芽生孢子生产的培养基成分和发酵参数,玉米浆69.25 g/L、酪蛋白80.68 g/L加磷酸氢二钾15.17 g/L,控制条件为26 ℃,pH 3.3,恒定搅拌速度635 r/min,发酵120 h,可获得最大产量1.25×108芽生孢子/mL[98]。这些可作为扩大规模生产的基础。但由于芽生孢子难于保存,至今未得到推广。
3.2 制剂工艺
为了提高田间应用效果,还需要研制菌剂配方和改进加工技术,以保持菌剂中真菌的活力,并使之满足贮存、运输和田间施用的要求。
早期的昆虫病原真菌田间应用都是直接采用发酵产品,并不受制剂加工,产品贮存和施用方式的限制。为了便于产品贮藏、施用和提高防效,需要将生产的菌体制剂化。制剂化的产品有利于标准化和登记管理,是产品商业化的重要前提。产品制剂化技术随着生产技术和规模的发展不断改进和成熟,但由于商业利益的原因,制剂关键技术很少公开,只能从产品数量、用量及有限的文献中获知一些信息。以绿僵菌为例,2015年,全球有13个国家或地区的83个绿僵菌产品注册在案,适用于蝗虫防治应用的主要有可超低量喷雾的油悬浮剂(有的称油剂,应纠正)[99],在不同环境和使用器具条件下可湿性粉剂、粉剂、饵剂也适于应用。
当初开展防治沙漠蝗国际合作项目 LUBILOSA的一个重要基础是英国已经研发获得了绿僵菌的油悬浮制剂,他们筛选到适宜的油基,能够在应用时很容易地将孢子均匀悬浮在油基中,制成可超低量喷雾的配方。该制剂优点是使感染性孢子很好地附着于昆虫表皮,同时孢子在油滴中得到保护,可避免在高温低湿环境下因快速失水干燥而失活。后来该产品经过毒理安全性、环境安全性评估,最终由南非的BCP公司和法国的NPP公司成功开发,在几个非洲国家获得注册,商品名为“GreenMuscle”,被FAO准许为蝗虫防治产品,并大量生产,多年在非洲和亚洲多个国家用于防治沙漠蝗[100]。
澳大利亚针对当地严重为害的蝗虫,也成功研发了高毒力金龟子绿僵菌FI-985的油悬浮剂,获得澳大利亚联邦科学与工业研究组织(Commonwealth Scientific and Industrial Research Organization,CSIRO)许可登记注册,商品名为“Green Guard”,并由SGB Australia Pty Ltd公司进行了大量生产,多年用于蝗灾防控,取得了巨大的成功。应用表明绿僵菌对其他昆虫、水生环境都没有不利影响,且无残留物,是对环境可持续发展友好的产品。Green Guard油悬浮剂(金龟子绿僵菌60亿孢子/mL)产品曾在2008年由澳大利亚贝克安德伍德公司在中国取得注册登记和应用。在美国,已注册了一种基于球孢白僵菌的产品,名为“Mycotrol”,可用于防治各种害虫,包括蝗虫和蚱蜢。
近20年来,我国真菌制剂研发和注册登记进展加快,自2002年第一个真菌杀虫剂获得注册登记,至今已登记的防蝗真菌农药有6个(种次)金龟子绿僵菌和5个(种次)球孢白僵菌(表3),防治对象为飞蝗、草原蝗虫、竹蝗等。近年来,有研究开发绿僵菌的饵剂、粉剂、可乳化粉剂,以利于在各环境下选择适合的剂型应用。产品的防治对象为飞蝗、草原蝗虫、竹蝗等。
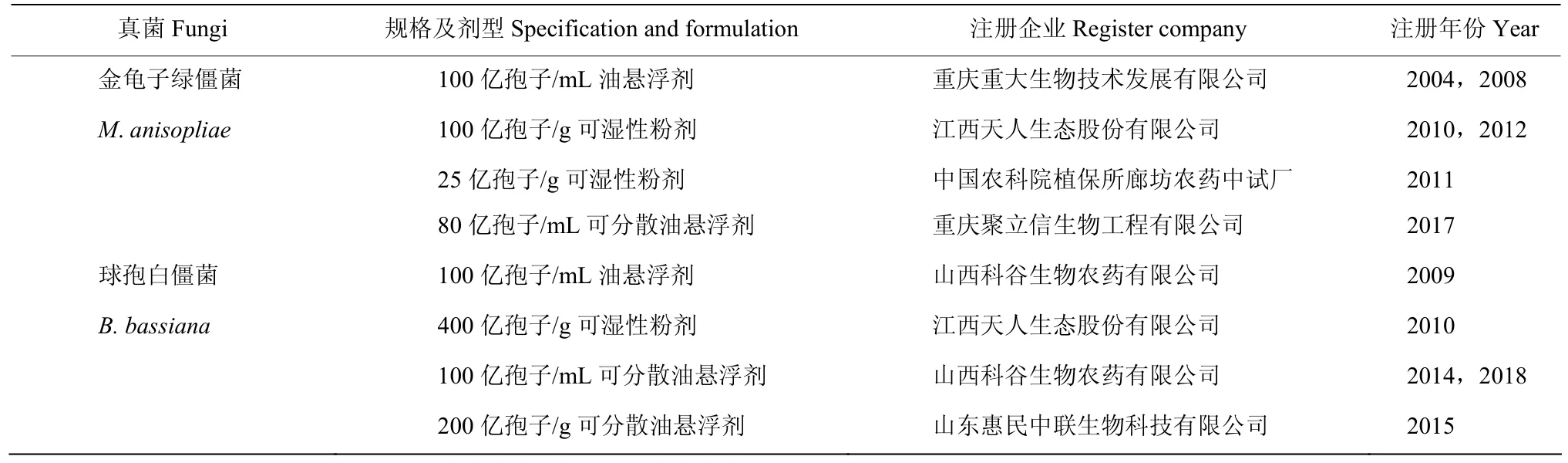
表3 我国已注册的真菌防蝗产品Table 3 Fungal pesticides registered for controlling locusts and grasshoppers in China
4 真菌防蝗技术和应用
4.1 真菌防蝗技术及其评价
真菌病害在蝗虫中的自然流行机率很低,需要人工增殖并释放到自然蝗虫种群中,提高蝗虫的感病率,才能将蝗虫种群密度控制在经济为害水平以下。
已报道了很多有潜力的蝗虫病原真菌在田间的防效试验,但付诸大规模应用的较少。应用产品主要是绿僵菌和少量白僵菌,剂型主要是孢子的油悬浮剂、可湿性粉剂,还有饵剂和粉剂。施用与评价方法参照化学农药,以施药处理后在较短时期内的蝗虫死亡率作为指标。总体来看,报道的防效是在施菌后1周内可见蝗虫死亡,在2~4周是死亡率上升期,4周后仍有少数死亡,总死亡率可达到60%以上,有的超过90%。由于产品中菌株毒力差异、剂型和施用方法差异等导致最终死亡率有较大范围波动。相对于化学农药,真菌的致死速率较慢而持续时期较长是一个显然的事实和重要特征。
实际上,对于田间蝗虫发生的不同情况如发生面积、发生量和种群发育阶段等,可采用不同的防治方案,包括在严重发生区、农作区的应急控害方案和在荒漠区、蝗虫幼龄阶段的适度容忍持续控制方案。方案的选择可依据经济与生态损失评估来制定阈值。对于我国内蒙古草原,根据5种优势蝗虫即亚洲小车蝗、毛足棒角蝗Dasyhippus barbipes、宽须蚁蝗Myrmeleotettix palpalis、狭翅雏蝗Chorthippus dubius和小蛛蝗Aeropedellus variegates minutus的自然种群结构和数量、蝗蝻和成虫的平均寿命、不同发育阶段的日食量等数据,通过估算蝗虫造成的牧草损失,提出了优势种蝗虫(3龄蝻期)的防治经济阈值为17~38头/m2,其中亚洲小车蝗最小(16.9)、小蛛蝗最大(37.4)[101]。对于不同区域的生产效益和生态承受能力,经济阈值有所变化,特别对于天然草场和荒漠区,其生态功能更为重要。然而,由于生态因子较多且变化复杂,很少有蝗虫防治的生态阈值的报告。生态阈值需要增加考虑生物多样性资源指数、受害指数、干旱因子、补偿易害指数等,一个可参考的生态阈值(Ecological threshold)简化模型定义为:ET=[(-1)/(D×LnR)]×[K/FL_n+CC/(FL_n×P×EC)],其中R为草原投影盖度;K为α%草原平均生产力水平;FL_n为蝗虫混合种群每虫损失估计;CC为防治费用;EC为防治效果;P为产品价格;D为干旱因子[102]。一项在内蒙古通辽草场进行的天然罩笼与人工模拟试验,对主要为害的亚洲小车蝗不同蝻龄的取食量与产草量损失曲线进行了拟合,并且考虑了补偿生长因子,确定该草场的防治指标应在5~15头/m2,防治适期为3龄或3龄以前,并结合防蝗生态阈值模型,计算出当地草场的生态经济阈值为11.04头/m2,可见计算结果与试验结果相符,说明采用生态经济阈值比传统的经济阈值评估更贴合实际[103]。
在防蝗应急方案中,针对大量发生的迁移性蝗虫,研究人员研发了菌药互补、混配复配与间隔施药等应急防控技术,可克服真菌起效慢的弱点,提高防治效果,同时有效减少化学农药用量,有利于蝗虫可持续防控。研究发现,金龟子绿僵菌与一些化学农药具有协同增效作用。比较氯虫苯甲酰胺(chlorantraniliprole)与金龟子绿僵菌处理的蝗虫体上10种酶活性变化,表明谷胱甘肽-S-转移酶(GST)、酯酶(ESTs)和苯酚氧化酶(PO)的活性在初期被抑制,而超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)的活性则在处理后的较晚期下降;芳基酰化酶(AA)和几丁质酶(CHI)在初期增加,而乙酰胆碱酯酶(AChE)在以后一段时间增加,它们分别在不同阶段调节昆虫对病原体的免疫和抗性能力。而且菌药联合处理增加了蝗虫死亡率,研究者推测协同作用破坏了宿主中Ca2+的响应机制和宿主体内的状态的平衡,从而诱导死亡[104]。经过兼容性筛选,确定了1%高效氯氰菊酯与100亿孢子/mL的绿僵菌油悬浮剂复配,共毒系数为263;喷施处理东亚飞蝗,第5 d的死亡率达到90%以上,比单独100亿孢子/mL绿僵菌油悬浮剂处理提前7~8 d,并降低化学农药用量90%,减少了对环境的非目标生物影响[105]。
荒漠区的经济价值相对较低,蝗虫幼龄阶段为害相对有限,可适度容忍蝗虫种群短期生存,此类情形适于应用真菌防控。施用的真菌可在几天或十几天内感染致病蝗虫,抑制种群发展,并且在种群及环境中定殖下来,持续防治当代和后代蝗虫。在非洲贝宁北部防控稻蝗Hieroglyphus daganensis的田间试验期间,监测到应用黄绿绿僵菌油制剂的感染性呈指数下降,半衰期为6.8 d,但残存孢子可继续感染蝗虫并占施菌12 d时总感染量的40%~50%;还开发了一个简单的宿主-病原体模型用于检测残留孢子在当季内和季后的感染,结果表明,残留孢子浓度的少量增加可使总死亡率大大增加,在蝗虫种群长效控制中起至关重要的作用[106]。在墨西哥使用蝗绿僵菌防治农田蝗虫Sphenarium purpurascens试验中,用分子方法和培养技术,监测了喷洒的绿僵菌在空气、土壤和营养覆盖层中的生存持久性,表明在施菌后的 66个星期内,绿僵菌在地表持续存活,种群数量显著波动,最高时达到初始浓度的4倍,后期逐渐下降到其初始浓度的一半左右。空气和营养覆盖层中绿僵菌浓度则稳步下降,8个月后完全消失[107]。在我国内蒙古锡林浩特牧场,应用金龟子绿僵菌防治蝗虫2年后,从试验区23个随机样点采集土壤,进行了菌株特异性SCAR标记检测,证明了施用的绿僵菌在田间可长期保持存活[108]。金龟子绿僵菌能够在植物根际和根内的宿存是其维持田间种群的重要原因,在干旱的内蒙古草原监测,绿僵菌施用后种群数量在30 d内快速下降,但能够在蝗虫喜食的羊草Leymus chinensis、针茅Stipa grandis根际土壤中以低密度延续宿存至少75 d;采用菌株荧光蛋白标记基因egfp的特异PCR检测,证明了绿僵菌在羊草和针茅根内宿存[109]。
绿僵菌能够在蝗虫种群中扩散传播,是维持长效防控的重要基础。当种群中病虫达到一定比例,绿僵菌疾病可由病虫传播给健康虫体。亚洲小车蝗3龄蝗蝻的罩笼试验表明,将绿僵菌处理3 d后的蝗蝻与未处理蝗蝻按 10:50、20:40、30:30、40:20混合饲养时,疾病传播几率分别可达 24.6%、31.0%、39.0%和52.0%。以同样处理和比例进行短星翅蝗Calliptamus abbreviatus的混合饲养,疾病传播几率分别为15.6%、23.5%、32.7%和40.0%。进一步田间试验发现,在圆形核心区施用绿僵菌后,施菌区外都出现蝗虫感染,感染率随时间的推移逐渐上升,在处理后40 d施药区外400 m八个方向的疾病感染率均值可达8.95%。由此,研究者提出间隔施菌的技术方案,并进行了试验评价,结果在间隔30、60、120 m 的间隔区域,施菌后49 d校正虫口减退率分别达到 58.39%、63.41%、57.17%,证明了蝗虫移动扩散的习性可以有效地将病原传播到未处理区域,这种间隔施药方式可以降低成本、提高效益,在推广绿僵菌防治蝗虫技术中有重要意义[110]。
4.2 真菌防蝗应用
鉴于沙漠蝗的大规模暴发和为了减少化学农药的污染,联合国粮农组织在 21世纪初成立了一个独立的咨询机构,即农药裁判组(PRG),其作用是基于粮农组织获得的各类科学数据,对可能用于蝗虫防控的制剂提出有效性、环境安全性和对非靶标生物风险性等预测,为受蝗灾影响的国家选择合适的杀虫剂提供建议。在咨询的备用产品库中,生物农药绿僵菌和昆虫生长调节剂被推荐为新的替代品[111]。在美国,为了防治西部种植区每年夏天的蝗虫和蟋蟀侵扰,美国农业部农业研究服务局和相关科研机构专家建议采用绿僵菌、白僵菌等真菌杀虫剂,并对种植者进行技术培训[112]。在我国,20年来持续开展以绿僵菌为主的草原蝗虫绿色防控技术研究和推广,在内蒙古、新疆、甘肃等草原区域进行了大规模示范,自 2012年起绿僵菌产品被列入农业农村部防控蝗虫灾害的推荐产品。
利用绿僵菌防治蝗虫是国内外微生物防治害虫最成功的实例之一。国际合作 LUBILOSA项目成功研发了绿僵菌产品GreenMuscle,连续多年在撒哈拉地区和非洲其他地区应用。通常的用法是在2 L的油基中加100 g孢子粉,配置成均匀悬浮剂,超低容量喷雾用于1 ha,即用量大约5×1012孢子/ha。在贝宁3个区域的环境系统进行了防蝗示范,调查显示,贝宁南部区域在施菌后10 d,蝗虫5龄或成虫的数量下降了90%;贝宁北部试验区中,茂密的植被不适于喷雾,但蝗虫的取食活动能从植物中补偿到一定孢子,在施菌后14 d防效可达70%。在其他区域的示范也获得了类似的结果[113]。项目结束后,产品继续商业化应用于非洲、亚洲多国的沙漠蝗和其他蝗虫防控。
另一个成功应用案例是绿僵菌防蝗产品 Green Guard被澳大利亚蝗灾管理委员会 APLC(Australia’s Plague Locust Commission)批准,在1998年11月到1999年3月间,用于超低容量的飞机喷洒,在东亚飞蝗的若虫发生地带进行防治。经调查,在高粱作物、开阔的草原或开阔的林地,施菌量3~4×1012分生孢子/hm2的条件下,处理后蝗虫的存活率下降到10%以下。在较低剂量或茂密的林地中,蝗虫种群下降较小,施菌处理后一周内,蝗虫种群数量变化不大,但随后移动范围迅速减小、速度变慢。在植被高大、茂密的区域,喷洒后,幼虫种群数量下降较慢,有的幼虫移动范围扩大,有的甚至移出处理区域。在用绿僵菌处理过的地带,若虫很少蜕变为成虫,即种群受到了抑制。相比之下,在邻近未经处理的地区有许多成虫集群,必须使用化学杀虫剂加以控制。澳大利亚APLC据此制定并实行了蝗虫防控指导方案,为在集群小、直接为害力有限的若虫阶段开始介入处理。试验示范明确了绿僵菌Green Guard能够有效抑制本地低容度飞蝗种群,它可作为蝗害预防控制计划的重要组成部分。绿僵菌对蝗虫的致死作用较慢,但不会妨碍其在此类方案中抑制蝗害的作用[114]。中国在实验室内和田间试验下也评估了澳大利亚Green Guard产品的防蝗效果。在实验室中,Green Guard产品导致东亚飞蝗4龄若虫70%~80%的死亡率。在天津和河南省进行的田间试验中,应用Green Guard油悬浮剂,使用背负式喷雾器处理了40 hm2(2002年)和160 hm2(2003年)草地,剂量为50~125 g孢子配制在1125~1500 mL油基中,用于1 hm2的超低容量喷雾,施菌处理8~11 d后,田间种群减少了65%~97%[115]。
在墨西哥,也利用绿僵菌防治包括蝗虫在内的多种害虫。根据国家农业食品质量安全管理局SENASICA(Servicio Nacional de Sanidad, Inocuidad y Calidad Agroalimentaria)2017年的报告,在墨西哥不同州,应用蝗绿僵菌在 41698 hm2进行了施菌处理,以控制不同种类的蝗虫和蚱蜢,包括 Schistocerca piceifrons piceifrons,Melanoplus sp.,Sphenarium sp.,Brachystola sp.[116]。
在埃及,对商业制剂Bioranza(金龟子绿僵菌)和Biovar(球孢白僵菌)进行了沙漠蝗防效评估,结果表明,无论对若虫直接喷洒油性菌剂,还是通过土壤处理的间接施用,绿僵菌对蝗虫的感染死亡率和快速效果均比球孢白僵菌高得多[117]。
在国内,中国农业科学院植物保护研究所(原生物防治研究所)从20世纪90年代初期开始昆虫病原真菌防蝗应用研究,不仅评估了不同绿僵菌菌株对东亚飞蝗的毒力[67],还在天津北大港水库蝗区,以绿僵菌孢子不同浓度的油剂喷雾进行防蝗试验,在田间处理后第10 d校正死亡率为81.5%,人工模拟小区试验在第10 d供试虫全部死亡[118],另外进行了10 hm2的田间试验,证明绿僵菌田间治蝗的良好效果[119]。20多年来,绿僵菌菌剂在北方多省区草原大面积应用。在新疆,用绿僵菌油剂防治干旱荒漠草原和山地草原蝗虫试验,表明真菌杀虫剂油剂具有杀虫谱广(对蝗总科昆虫均有效)、容易大规模生产、便于低容量喷雾和适于干旱气候等优点[120,121]。在内蒙古草原进行了用绿僵菌油剂超低量喷雾防治蝗虫的试验,表明菌剂对5种蝗虫(亚洲小车蝗、红翅皱膝蝗、毛足棒角蝗、白边痂蝗、轮纹异痂蝗)的防效显著。喷药后8 d,对主要蝗虫亚洲小车蝗(占混合虫口的92.2%)的防治效果达48.0%,12 d后增至88.1%。死亡虫体上的绿僵菌孢子的扩散有可能继续起着控制蝗虫虫口密度的作用[122]。在甘肃用黄绿绿僵菌饵剂和油剂防治草原蝗虫,虫种有祁连山痂蝗Bryodema qilianshanenensis等几种,投撒饵剂后用样筐取样调查,显示7和12 d的平均防效为66.5%和76.7%;油剂试验以静电超低容量喷雾,防效高于绿僵菌饵剂,7和12 d的平均防效为75.8%和84.6%。施药后3~4 d感病死亡的虫态多为4~5龄若虫,25 d后出现死亡成虫,之后2周仍陆续有死亡虫体,表明绿僵菌有较好持续控制效果[123]。多年来,在新疆、内蒙古、甘肃、青海等地都有大面积示范推广试验,表明绿僵菌制剂可迅速达到控制高密度蝗群的目标,还能实现长期传播控制,致使蝗群不能为害成灾[1]。绿僵菌产品还在河滩、湖区荒滩等蝗虫滋生地防治东亚飞蝗的低龄蝗蝻[124,125],有效抑制其种群发育,防止成虫集群起飞,把可能对农区的大规模为害遏制在种群迁飞之前。
另外,还有广谱的球孢白僵菌产品可用于防治蝗虫及其他害虫,如在美国注册的产品Mycotrol可防治各种害虫,包括蝗虫和蚱蜢。LUBILOSA项目在西非的试验中也测试了球孢白僵菌,但发现在田间环境的高温度下,其效力不如绿僵菌菌株。在中国黑龙江齐齐哈尔索伯台草原,应用白僵菌100亿孢子/mL油悬浮剂防治草原蝗虫,用量为2250~3000 mL/hm2,药后7 d达到作用高峰,平均防效为68.24%~77.44%,防效可持续7 d[126]。在竹林中用球孢白僵菌防治黄脊竹蝗Ceracris kiangsu跳蝻,得到的校正死亡率达到68.2%[127]。
有记录噬虫霉类真菌可导致田间发生蝗虫流行病,说明此类真菌对蝗虫具有高致病力,也分离到相关菌株,但由于难以人工大量培养和贮存,至今未能用于蝗虫防治实践。
5 讨论与展望
5.1 昆虫病原真菌防蝗的优势与不足
昆虫病原真菌资源广泛,专性侵染昆虫(节肢动物)而对脊椎动物无毒无害,与环境友好兼容,还具有对寄主不易产生抗药性、许多种类可人工大量培养等优点,使之成为最有前景的生物杀虫剂。近 30年来,针对蝗虫的研究进展显示,从绿僵菌、白僵菌等种类可筛选出对蝗虫高毒力的菌株,并且可通过常规筛选或遗传改良来提高菌株毒力、生长性状和抗逆能力,通过优化培养基和可控条件大量培养菌剂,通过改善剂型技术来维持贮存活力和增强对蝗虫的侵染,研究改善施用技术,使其尽可能适应蝗虫习性和生态,以提高防治效果。利用绿僵菌和白僵菌防蝗已经从实验室走向田间应用并商业化。
然而,研究和实际应用中仍存在一些问题,以至于产品注册和大规模示范后也未能大规模持续商业化应用。究其原因,在研究和技术方面可能有以下几点:1)蝗虫种类繁多,而菌株杀虫毒力通常只对某些蝗虫种类强而对其他蝗虫种类较弱,因此产品对不同区域的蝗虫的防效有波动,而市场的普适性要求较高;2)根据自然规律,真菌对蝗虫的致病过程需要一定时间,通常需要3~5 d甚至更长时间才能见效,而市场通常期待快速应对严重蝗害;3)真菌防蝗产品价格相对较高,即使已研究改进了发酵工艺,仍然由于工艺过程较复杂,活体贮存条件较高,在应用规模有限的情况下,成本竞争力仍不敌化学农药;4)真菌制剂的使用技术要求相对较高,一般需要专业人员或需经过专业培训,指导用户使用时把握蝗虫发育进度和掌握避免真菌失活的时机,以达到高效防治。
5.2 真菌防蝗应用的策略与展望
鉴于真菌防蝗产品的特点以及当前的研究进展,作者认为,在未来研究中,可进一步通过菌种的遗传改良来提高菌株的毒力、普适性和杀虫速度,通过改进发酵工艺降低产品成本,通过制剂技术提高田间杀虫效果。然而,一个重要的防治策略是在蝗蝻幼龄期施菌干预种群发展以防止蝗灾暴发。因为基于真菌与蝗虫各自的生物学特点及其致病关系,蝗虫在幼龄阶段扩散较慢、为害较小。试验证明,低龄期(2~4龄)比高龄期(5龄)或成虫更容易感然病害;当出现末龄幼虫或成虫迁飞时施菌,可能效果甚微或无济于阻止和减少已经造成的损害[64,128]。在幼龄蝗蝻期间施用菌剂如绿僵菌等,可使病原真菌在蝗虫种群中侵染定殖,即使没有快速引起多数虫体发病死亡,也能够抑制发育、减缓扩散,同时虫体携带病原真菌移动扩散,一旦罹病死亡,便能够大量增殖病原体并释放出来,作为新的侵染源再度影响蝗虫发育和种群扩张。另外,病原真菌具有田间扩散性和在根际土壤及植物体内中有一定的宿存能力[129,109],加上一些制剂手段有助于真菌的宿存和增殖,有可能在蝗虫栖境中长期保存一定密度的病原种群,借助环境因子实时捕捉机会侵染寄主,介入蝗虫种群的消长,从而抑制蝗虫种群暴发增长。
为了实施蝗蝻早期人工干预的策略,对蝗虫滋生发源地带的发现和监测十分重要。蝗虫多在水域附近荒滩产卵孵化,这些区域通常远离监视,无人治理或无主管理,因此蝗蝻种群得以顺利发展、集群乃至暴发迁飞到农田为害作物。对于无人治理或无主管理的虫源地带,因经济效益低,还不能商业化进入,只能依靠政府和公益性组织的联合,通过大数据如高空遥感监测结合气象与生物学信息等,发布监测预警,及时组织人工干预或征购商业服务。
蝗灾防控涉及经济和生态的长期综合管理,当前沙漠蝗严重扩张为害,再次引起各国政府和相关行业的重视。从真菌防蝗研究与应用进展上看,以绿僵菌为主的真菌防蝗技术已经相对成熟,无论在不同地区的防蝗效果或规模化生产技术都能够支持参与大规模蝗害防控,可作为蝗虫防控的重要组成部分。根据其特点和防效评估,真菌防蝗更适于在蝗蝻期施用,以干预种群发展、避免暴发成灾。通过技术完善和市场优化,具有高效、长效和环境友好特性的昆虫病原真菌产品必将在蝗灾防控中发挥更重要的作用。
