碳基纳米铜复合材料对普通小球藻胁迫作用的研究
2021-05-10吉喜燕唐静懿叶璟吴世超黄赛花侯梅芳李鑫许文武
吉喜燕,唐静懿,叶璟,吴世超,黄赛花,侯梅芳, ,李鑫,许文武
1. 上海应用技术大学生态技术与工程学院,上海 201418;2. 广东省科学院生态环境与土壤研究所/华南土壤污染控制与修复国家地方联合工程研究中心/广东省农业环境综合治理重点实验室,广东 广州 510650;3. 上海应用技术大学化学与环境工程学院 上海 201418;4. 上海应用技术大学轨道交通学院 上海 201418
纳米材料在医学、生物等领域的广泛应用使其不可避免地进入各种环境介质之中,其中水环境是纳米类材料的主要受纳介质(Chen et al.,2017)。不仅单质类纳米材料对水环境中生物的影响备受人们关注,碳基纳米复合材料对水生态的影响也成为了研究热点之一,碳基纳米铜就是其中的典型代表之一(Gao et al.,2020)。当碳基纳米铜进入水环境介质后会对水体中的藻类产生一定的影响(Windfeld et al.,2013)。由于普通小球藻具有普适性强、生长力旺盛等特点,因此常用于水生生物毒理学实验(孙红羽等,2019)。
据报道,碳基纳米铜对藻类的毒性作用与其他纳米单质或纳米氧化物相似,碳基纳米铜复合材料暴露下的藻细胞会被纳米材料包裹,形态会发生明显变化(刘雅等,2016)。当碳基纳米铜浓度超过30 mg·L-1时,普通小球藻细胞中活性氧(ROS)的水平升高,超氧化物歧化酶(SOD)活性增强,藻细胞的生物量降低,细胞活性下降(Costa et al.,2020)。碳基纳米铜悬浮液等水相碳基复合材料的毒性在不同水生生物体中存在差异,有研究者推断,这是由于碳基纳米铜对不同水生生物细胞表面的作用存在差异而导致的(Eltarahony et al.,2018)。不同浓度的碳基纳米铜复合材料对藻细胞的数量、叶绿素 a含量等具有明显的影响,当碳基纳米铜材料浓度范围为5—10 mg·L-1时,碳基纳米铜的存在对蛋白核小球藻(Chlorella pyrenoidosa)和斜生栅藻(Scenedesmus obliquus)的叶绿素a含量具有促进作用;而同等浓度下的单一碳粉则对二者的叶绿素 a含量无明显影响(Fazelian et al.,2019)。对于蛋白核小球藻而言,当碳基纳米铜的浓度高于60 mg·L-1时,其对藻细胞生物量的增长出现了严重的抑制现象,藻细胞的比增长速率几乎为0(Mahawar et al.,2018)。据文献报道,实际河、湖水体中碳基纳米铜的水平最高可达约60 mg·L-1,水体中的碳基纳米铜复合材料除了会对藻细胞的生物量、形态、叶绿素a含量等造成一定的影响,还会对藻细胞的生理代谢过程也会产生扰动作用(Dimkpa al.,2012;Lammel et al.,2019)。以往关于碳基纳米氧化锌对小球藻的生物毒性的研究中,发现低浓度的碳基纳米氧化锌对小球藻的生理活性具有抑制作用,小球藻基因的完整性会受到碳基氧化锌破坏并出现氧化应激反应(Alho et al.,2020)。上述关于纳米类、碳基纳米类材料的藻细胞生理毒性研究中,并未涉及到藻细胞的代谢物丰度及相关代谢途径的分析,尤其是碳基纳米铜对普通小球藻代谢方面的胁迫作用的研究仍然较少。
为此,本文以普通小球藻为研究对象,从藻细胞叶绿素a含量和藻细胞代谢组学特性变化两个方面入手,研究不同浓度碳基纳米铜复合材料(0、0.05、0.5、5、50 mg·L-1)对普通小球藻的胁迫作用。本文旨在探究碳基纳米铜复合材料胁迫下普通小球藻的差异代谢物丰度和对应显著富集通路的变化,丰富碳基纳米材料对藻细胞胁迫作用相关研究的数据和资料,为碳基纳米类复合材料与水生生态关系的研究提供科学参考。
1 材料与方法
1.1 实验设计
本文实验设计包括普通小球藻叶绿素a含量变化和藻细胞代谢组学变化两部分,设置碳基纳米铜的浓度为0、0.05、0.5、5、50 mg·L-1。实验装置如图1所示。
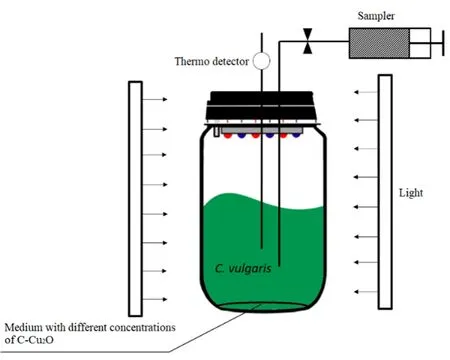
图1 实验装置图Fig. 1 The setup of experiment
如图1所示,实验装置置于无菌培养箱中,由反应器(250 mL反应瓶)、光源、采样管3个主要部分组成。锥形瓶中含有 150 mL人工成合成的具有不同浓度碳基纳米铜的BG-11培养基。每个实验组及对照组均设置平行组(n=3,n为每个平行组的数量)。调节光照为 120 μmol·(m2·s)-1,温度为 28 ℃,藻细胞初始浓度为 1×105cell·mL-1。
1.2 藻种培养
本研究选用普通小球藻作为受试对象进行实验,所用的藻种购买自中国科学院武汉水生所淡水藻种库,编号FACHB-8。藻种在光照培养箱中进行培养及扩培,选取BG-11培养基,恒温28 ℃,设置光暗比为 12 h∶12 h,光照强度维持在 120 μmol·(m2·s)-1。BG-11 培养基成分列于表1。

表1 BG-11培养基成分Table 1 The BG-11 medium components
将实验室扩培的藻及菌用于实验之前,需将普通小球藻和地衣芽孢杆菌分别以 8000×g离心 10 min并弃去上清液。用 0.1 mol·L-1的 PBS(PBS,Phosphate buffered solution)缓冲液(无菌超纯水配置)重复离心、洗涤3次,以确保藻类粘连的培养基去除干净才可正式接种于实验中培养体系中。
1.3 主要指标检测与表征
1.3.1 普通小球藻叶绿素a浓度检测
首先,将10 mL藻菌混合物样品进行离心处理,转速为4000×g,离心时间10 min,离心后弃去上清液。将所得沉淀溶于 10 mL体积分数 90%的丙酮中,将样品置于旋涡震荡仪混匀。然后置于4 ℃黑暗环境中处理24 h。将样品取出后再次进行离心处理,离心转速为4000×g,离心时间15 min。离心后的上清液用于叶绿素a浓度测定。紫外分光光度法测定波长为630、645、663、750 nm的吸光度,以90%丙酮溶液作为空白对照。叶绿素 a质量浓度(μg·L-1)通过式 1 计算(Ptushenko et al.,2017):

式中,Chl-a表示叶绿素质量浓度,单位为μg·L-1;OD663、OD750、OD645、OD630分别为对应波长下样品测得的吸光度数值;V为样品体积,单位为mL。
1.3.2 普通小球藻代谢组学检测
取适量样品,加入500 μL甲醇-水(4∶1)溶液,低温下采用高通量组织破碎仪进行破碎;加入 200 μL氯仿,冰浴超声萃取30 min,4 ℃、12000 g条件下离心20 min,取上清液,真空抽干;加入80 μL的甲氧胺盐酸盐吡啶溶液,于 37 ℃肟化反应 90 min;加入 80 μL 的 BSTFA(含 1%TMCS)和 20 μL的正己烷,70 ℃反应60 min,进行气相色谱-质谱联用(安捷伦8890B-5977B)分析。样品经HP-5MS毛细管柱(30 m×0.25 mm×0.25 μm)分离后进入质谱,全扫描模式,质量扫描范围为m/z50-500。在GC-MS检测分析的过程中,采用质控样品评价整个分析系统的稳定性。
采用 MassHunter软件进行预处理,同时利用Fiehn数据库对代谢物进行搜库鉴定,导出包含代谢物名称、保留时间、质核比和峰面积等的数据矩阵。采用偏最小二乘法判别分析PLS-DA分析来区分各组间代谢轮廓的总体差异,变量权重VIP值大于1,同时单因素方差分析显著性分析P<0.05(FDR校正)的代谢物被认为是发生显著变化的差异代谢物。采用 KEGG通路数据库(http://www.genome.jp/kegg)、MetaboAnalyst(https://www.metaboanalyst.ca/faces/home.xhtml)对差异代谢物进行代谢通路分析,获得差异代谢物的生物功能、显著富集的代谢通路等信息。
1.4 数据分析
本文使用 GraphPad Prisim进行单因素分析(ANOVA,Analysis of Variance)。代谢物分析使用SIMCA-P 14.0软件进行偏最小二乘判别分析(PLSDA,Partial Least Squares Discriminant Analysis)。使用MeV软件(4.8.1版)绘制代谢物相对含量的热图。本文中图主要使用Graphpad Prisim绘制。
2 结果与讨论
2.1 小球藻叶绿素a含量变化分析
叶绿素是光合作用的物质基础,光合作用是影响小球藻细胞生长的重要因素,叶绿素的含量骤降会导致小球藻光合作用效率降低,因此,在碳基纳米铜复合材料暴露下普通小球藻细胞内叶绿素a的降低将直接影响小球藻的光合作用和生长状态(Luimstra et al.,2019;朱津辉等,2019)。对实验周期内不同浓度碳基纳米铜体系第2、4、6、8天的叶绿素a含量进行了检测,结果如图2所示。
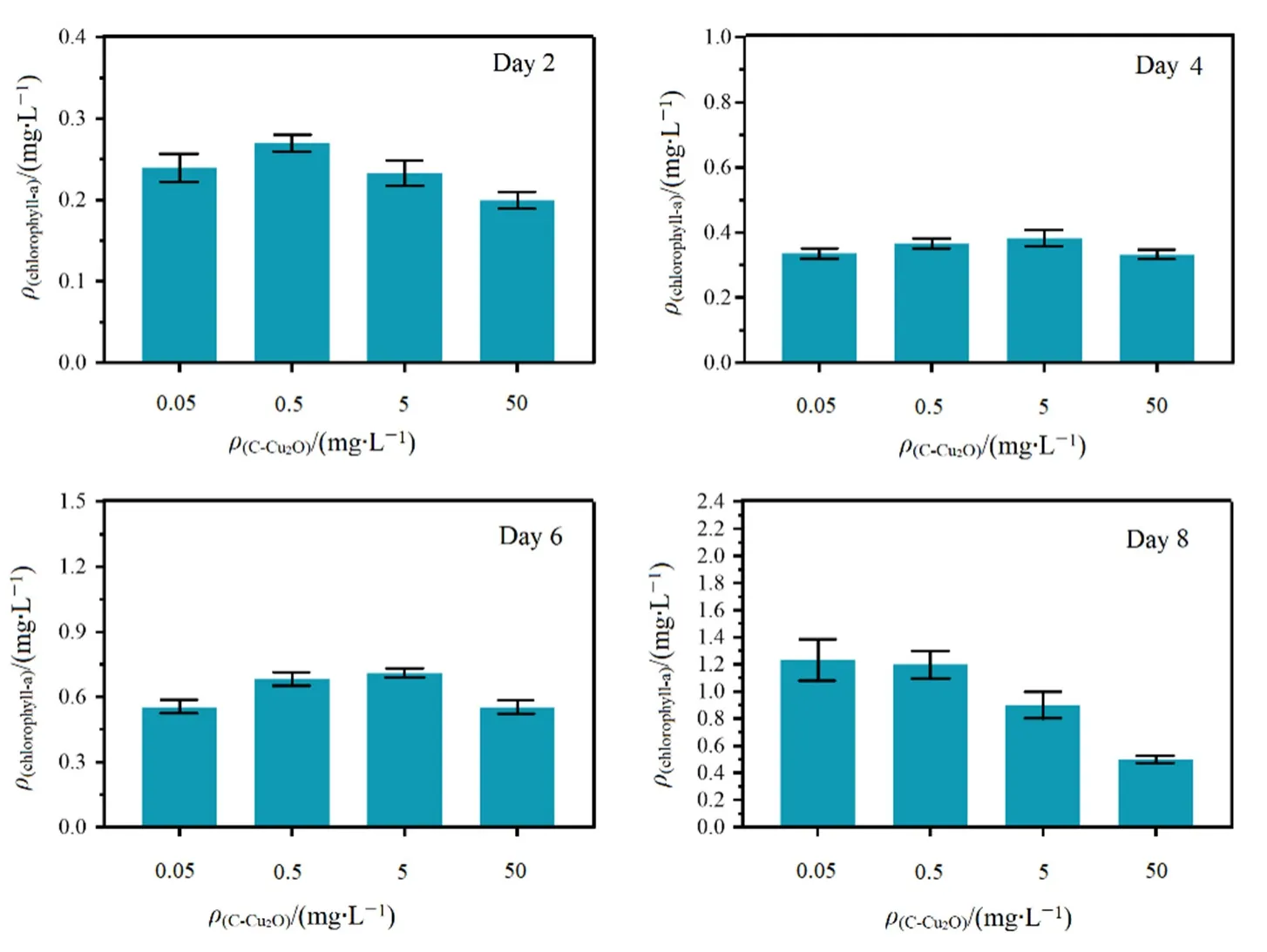
图2 不同浓度碳基纳米铜浓度条件下普通小球藻的叶绿素a含量变化Fig. 2 The chlorophyll-a content of C. vulgaris under different concentrations of C-Cu2O
由图2可知,0.05—5 mg·L-1碳基纳米铜暴露条件时,普通小球藻叶绿素a含量逐渐增加,这表明低浓度碳基纳米铜对普通小球藻叶绿素a含量变化的影响并不明显。50 mg·L-1碳基纳米铜暴露条件下的普通小球藻叶绿素 a含量随时间的推移明显降低,这表明高浓度碳基纳米铜对普通小球藻叶绿素a含量有明显的抑制作用。这与以往研究中低浓度碳基纳米铜(5—10 mg·L-1)对普通小球藻叶绿素a含量并无明显影响而高于20 mg·L-1的浓度时,碳基纳米铜对普通小球藻叶绿素a含量产生抑制作用的结果一致(Fazelian et al.,2019)。
2.2 碳基纳米铜迫条件下的普通小球藻代谢组学特性变化分析
2.2.1 不同浓度碳基纳米铜条件下普通小球藻的差异代谢物分析
不同浓度碳基纳米铜材料会对普通小球藻的差异代谢物产生一定的影响,碳基纳米铜的浓度越高,对代谢物种类的影响也越大,实际自然水体中检测到的碳基纳米铜的浓度范围为0.5—50 mg·L-1(Alho et al.,2020;Miao et al.,2020)。在实验周期的第 8天,观察发现 5 mg·L-1和 50 mg·L-1实验组中,叶绿素a浓度变化差异最为显著,而叶绿素a的产生是普通小球藻细胞内代谢调控的结果,基于此,对 5 mg·L-1和50 mg·L-1实验组进行代谢组学检测,以验证碳基纳米铜复合材料对普通小球藻胞内代谢所产生的影响。对各浓度碳基纳米铜体系中的普通小球藻进行洗涤,进行代谢组学测试分析。对5 mg·L-1和 50 mg·L-1碳基纳米铜体系及空白对照组体系中的普通小球藻代谢轮廓进行 PCA(Principal component analysis)分析,结果如图3A所示,5 mg·L-1碳基纳米铜组与对照组代谢轮廓有部分交叉,而50 mg·L-1碳基纳米铜组则距离二者较远。此结果表明,50 mg·L-1碳基纳米铜体系中的普通小球藻出现严重的代谢紊乱,代谢轮廓发生明显偏移,而5 mg·L-1碳基纳米铜则未能明显扰动普通小球藻的代谢。这表明50 mg·L-1碳基纳米铜对普通小球藻的代谢特性产生了较为明显的影响。相较之下,5 mg·L-1碳基纳米铜则对普通小球藻胞内代谢物变化的影响不明显。造成这种现象的原因是由于普通小球藻本身对外界胁迫具有较强的耐受性,相较之下,同属绿藻门的斜生栅藻则对碳基纳米铜复合材料更为敏感,当受到水体环境中同等浓度碳基纳米铜复合材料的胁迫时,其代谢轮廓更容易发生变化,会比普通小球藻更易发生较为明显的质壁分离现象,同时,斜生栅藻也比普通小球藻更易内吞水体环境中的碳基纳米铜复合材料,从而造成胞内细胞器的损伤(Landa et al.,2017;Wang et al.,2020)。
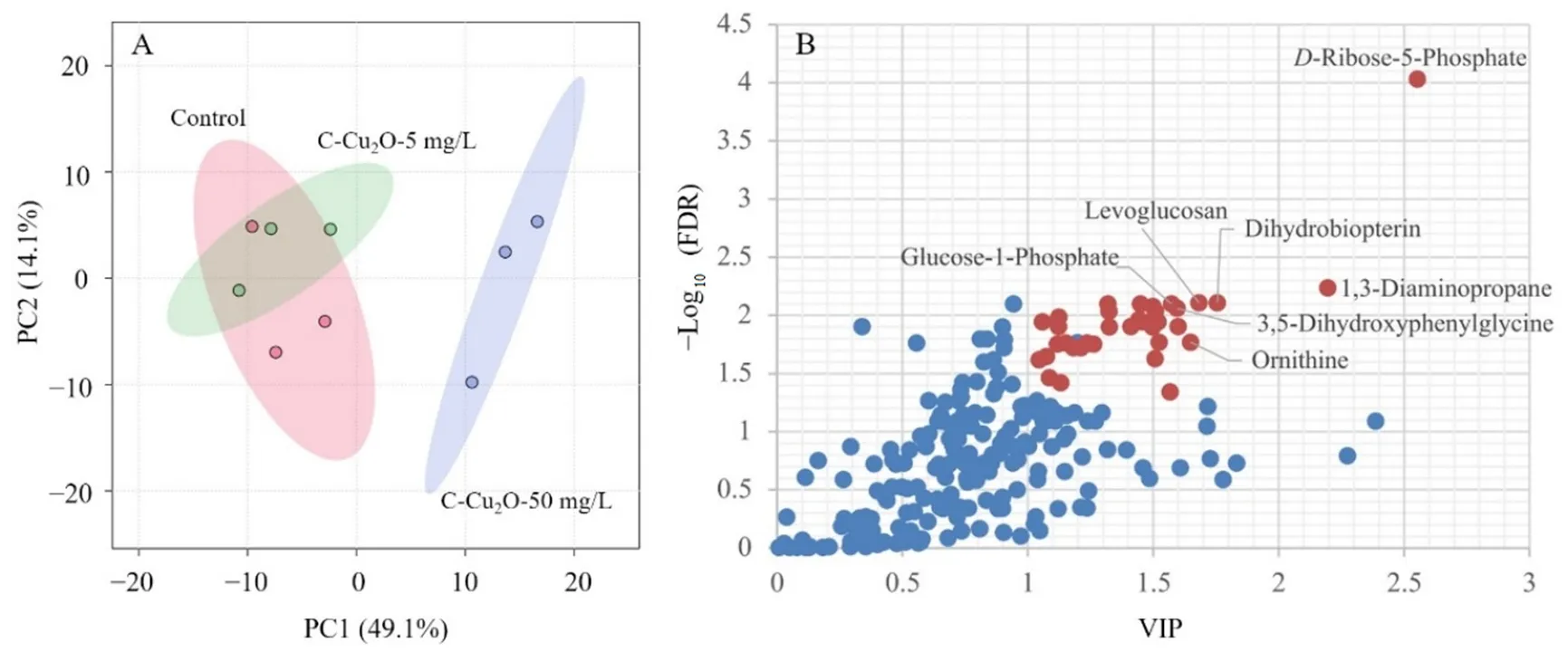
图3 不同浓度碳基纳米铜材料体系中藻细胞代谢物主成分分析Fig. 3 PCA of C. vulgaris metabolites in C-Cu2O nanoparticles system with different concentrations
为了进一步有效地找出与高浓度碳基纳米铜相关的内源代谢物,进一步对普通小球藻代谢轮廓数据进行PLS-DA分析,获得能直观反映3组间代谢差异贡献率的VIP图(图3B)。通过对相关数据进行独立样本ANOVA检验,筛选VIP值大于1且P值小于0.05(FDR校正)的差异代谢物,通过HMDB、KEGG和MetPA等数据平台进行检索,最终确定了如表2所示的36种差异代谢物。

表2 空白对照组与5 mg·L-1、50 mg·L-1碳基纳米铜体系中普通小球藻的差异代谢物Table 2 Different metabolites of C. vulgaris in control group and 5 mg·L-1, 50 mg·L-1 C-Cu2O nanoparticles
表2中,碳基纳米铜与空白对照组间代谢物相对含量升高,用“↑”标记;代谢物相对含量降低,用“↓”标记。与对照组比,5 mg·L-1和 50 mg·L-1碳基纳米铜体系中,左旋葡聚糖(Levoglucosan)、葡萄糖-磷酸(Glucose-Phosphate)显著上调;其中,5 mg·L-1碳基纳米铜体系中,哌啶酸(Pipecolic Acid)、氧代戊二酸(Oxoglutaric acid)显著上调。与空白对照组相比,5 mg·L-1和 50 mg·L-1碳基纳米铜体系中,1, 3二氨基丙烷(1, 3-Diaminopropane)、腺苷-5-但磷酸盐(Adenosine-5-Monophosphate)、丁烷-2, 3-二醇(Butane-2, 3-Diol)、二羟丙酮磷酸( Dihydroxyacetone phosphate)、羟胺(Hydroxylamine)、草酸(Oxamic Acid)、核酮糖-5-磷酸盐(Ribulose-5-Phosphate)、维生素C(Ascorbic Acid)、反丁烯二酸(Fumaric acid)、二十一烷酸(Heneicosanoic Acid)、木酮糖(D-Xylulose)、磷酸海藻糖(Trehalose-6-Phosphate)等代谢物显著下调。上述差异代谢物变化表明,较高浓度碳基纳米铜(50 mg·L-1)可对普通小球藻的糖类和小分子酸类代谢物产生明显影响,导致这两大类代谢物的下调。将所得到的差异代谢物进行聚类分析,得到如图4所示的热图。

图4 空白对照组、5 mg·L-1、50 mg·L-1碳基纳米铜体系普通小球藻代谢物热图Fig. 4 Heatmap of the C. vulgaris metabolites in control, 5 mg·L-1, and 50 mg·L-1 C-Cu2O nanoparticles groups
图4中,红色色块表示对应的代谢物相对丰度的增加,蓝色色块代表对应代谢物的减少,颜色越深,代表增加或减少的程度越重。当碳基纳米铜的浓度从 5 mg·L-1上升至 50 mg·L-1时,大部分差异代谢物的相对含量降低,这些代谢物多为糖类、氨基酸类和小分子酸类。这表明,高浓度的碳基纳米铜对普通小球藻产生了显著的抑制作用,同等胁迫条件下,蓝藻门的铜绿微囊藻对碳基纳米铜的耐受能力远弱于普通小球藻,高浓度的碳基纳米铜复合材料会导致铜绿微囊藻细胞内部的淀粉粒数量明显增加,大量的铜绿微囊藻细胞出现细胞膜破裂,与此同时,高浓度(50 mg·L-1)碳基纳米铜的存在会使铜绿微囊藻的氨基酸循环遭到严重破坏(Faraz et al.,2019;Mavrikou et al.,2019)。
2.2.2 不同浓度碳基纳米铜体系中藻细胞差异富集通路分析
在讨论了不同碳基纳米铜浓度对普通小球藻细胞代谢物种类数量变化、差异代谢物相对丰度的基础之上,对差异代谢物进行 MetPA(Metabolomics Pathway Analysis)分析。结果如图5所示,横坐标为impact,纵坐标为KEGG通路富集程度,图中实心圆的大小代表该通路impact值。图中圆点越大,表示通路impact值高,impact大于0.1为重要扰动通路;图中颜色表示显著性,颜色越深,表示P值越小。发现了与这些生物标记物相关的9种代谢物显著富集通路,分别为精氨酸合成通路(Arginine biosynthesis)、戊醣酸通路(Pentose phosphate pathway)、葡萄糖合成通路(Gluconeogenesis)、半乳糖代谢通路(Galactose metabolism)、抗坏血酸和醛酸代谢通路(Ascorbate and aldarate metabolism)、嘧啶代谢通路(Pyrimidine metabolism)、氨基糖和核苷酸糖代谢(Amino sugar and nucleotide sugar metabolism)。
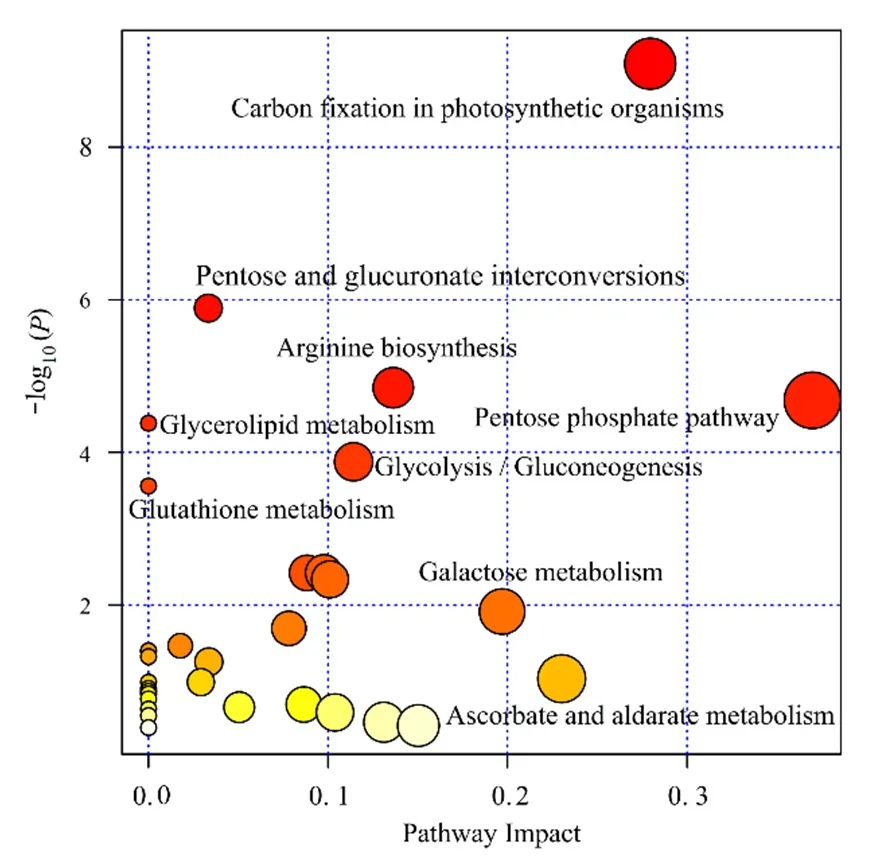
图5 不同浓度碳基纳米铜材料体系中藻细胞MetPA通路图Fig. 5 MetPA pathway of C. vulgaris in different concentrations of C-Cu2O nanomaterials
如图5所示,代谢通路分析表明,差异代谢物富集最多的是光合作用碳固定代谢通路(Carbon fixation in photosynthetic organisms),其次是氨基酸糖代谢通路(Amino sugar and nucleotide sugar metabolism)。高浓度碳基纳米铜影响普通小球藻的机体代谢,抑制光合作用的碳固定过程,造成藻细胞体内糖、氨基酸等物质的稳态变化(Chen et al.,2019;Gong et al.,2018)。研究表明,藻细胞内氨基酸的合成与叶绿素a的形成密切相关(Lammel et al.,2019;Yarmohammadi et al.,2019),这与高浓度碳基纳米铜导致普通小球藻叶绿素a含量明显下降的现象吻合。当藻细胞处于外界胁迫后,细胞会启动自我防御机制,这种防御的常见表现之一是细胞内淀粉粒的增加,而淀粉粒的增加与糖代谢通路的显著富集有密不可分的关系(Kumar et al.,2015)。
图6为不同浓度碳基纳米铜复合材料对普通小球藻产生胁迫作用后,藻细胞内部几大类主要代谢通路和通路途径中代谢物的变化。红色标记的为上调的代谢物,蓝色标记的为下调的代谢物。高浓度碳基纳米铜的胁迫作用使藻细胞中糖代谢、氨基酸代谢、核酸代谢这三类代谢的特性发生了明显的变化。高浓度的碳基纳米铜会导致小球藻主要通路活性的降低(Lauritano et al.,2019)。图6中,卡尔文循环(Calvin cycle)中的大部分代谢物降低,光合碳循环代谢被显著削弱;同时,藻细胞内的糖代谢也被明显抑制,作为三羧酸循环(TCA cycle)上游产物的三磷酸甘油,其在卡尔文循环中含量的降低也导致了三羧酸循环中一部分代谢物的降低。这一变化进而间接导致抗坏血酸和鸟氨酸等氨基酸的代谢降低,从而减弱整个氨基酸代谢通路(Hu et al.,2020)。

图6 不同浓度碳基纳米铜材料体系中藻细胞代谢通路变化Fig. 6 Metabolic pathway of C. vulgaris in different concentrations of C-Cu2O nanomaterials
3 结论
本文以普通小球藻为受试对象,设置了不同浓度的碳基纳米铜暴露条件,在实验周期内进行了藻细胞浓度变化、叶绿素a含量、代谢组学变化的测定,得出以下结论:
当碳基纳米铜质量浓度范围为 0.05—5 mg·L-1时,其对小球藻生长及叶绿素a含量并没有明显的抑制作用;当碳基纳米铜浓度达到50 mg·L-1时,其对普通小球藻产生了较为严重的胁迫作用,多数氨基酸的相对丰度降低,这表明高浓度的碳基纳米铜复合材料使得藻细胞内氨基酸代谢活力降低,这也导致藻细胞内差异代谢物和代谢路径的变化。高浓度的碳基纳米铜复合材料导致藻细胞内甘氨酸代谢被破坏,其相对丰度显著降低,这直接导致了藻细胞光合作用的碳固定能力减弱;这些内在的胁迫作用直接导致高浓度碳基纳米铜复合材料胁迫条件下藻细胞内叶绿素a含量的下降。
