封育方式对荒漠草原群落特征及土壤理化性质的影响
2021-05-10刘建康张克斌冯湘刘新月
刘建康,张克斌,冯湘,刘新月
1. 宁夏大学/西北土地退化与生态恢复国家重点实验室培育基地,宁夏 银川 750021;2. 宁夏大学/西北退化生态系统恢复与重建教育部重点实验室,宁夏 银川 750021;3. 水土保持国家林业局重点实验室/林业生态工程教育部工程研究中心/北京林业大学,北京 100083
中国草地多分布于降水稀少的西北内陆地区,生态环境极其脆弱。近年来在人类活动日趋频繁及全球气候日渐变化影响下,中国大部分天然草场正面临不同程度退化(Wang et al.,2017;Gang et al.,2014),甚至可能引起土地沙漠化和荒漠化。这直接威胁着当地畜牧业的发展和当地居民的生产生活,现阶段草地退化已成为一项制约中国生态经济发展的环境问题。而放牧作为西北地区植被演替的直接和主要驱动因子(Akiyama et al.,2007),是中国草地生境恶化的主要人为因素之一,如何恢复由超载放牧引起的退化草地受到政府和广大生态学者的广泛关注。为改善草地生态环境,中国开展了一系列的生态治理工程,治理方针主要以保护自然植被或构建人工植被为主,选择的治理措施多种多样,其中围栏封育因投资少、实施简单、见效快和可扩展性的特点已成为退化草地恢复的主要措施之一(Liu et al.,2019;Cheng et al.,2016),主要利用生态系统自我修复能力恢复退化植被,而封育的恢复效果、机理及过程一直是学者的研究重点。
有生态学者指出放牧干扰可通过选择性采食、践踏等行为直接影响植被群落生长发育,又可通过排泄物沉积行为影响土壤理化性质,最终反作用于植物群落(Tao et al.,2013;刘晨,2015)。而在草地围栏封育后,由于缺乏牲畜干扰,草地土壤-植被生态系统中的养分和能量向食草动物的循环流动过程显著降低,这时充足的自然资源以及较弱的种间竞争压力,利于植被群落的自然繁殖更新,促使植被群落迅速恢复(Zhu et al.,2016;Jing et al.,2014;刘小丹等,2015)。封育后群落恢复是一个植物与土壤、环境之间长期相互作用的演替过程,有学者研究指出围栏封育可通过直接或间接方式改变群落物种组成、提高植被覆盖度和群落生产力、增加物种多样性、提高群落稳定性、增加种子库、改善土壤理化性质(Li et al.,2017;聂莹莹等,2020;尹亚丽等,2019;Socher et al.,2012)。植被恢复效果除与草地封育方式密切相关外,还受区域环境特征、样地围封时间等因素影响,这主要是由于区域环境因素(如地形、土壤养分、水分、地貌、光照等)分布的复杂性和不规则性,以及自然资源的有限性致使不同地区在不同封育时间后恢复效果存在差异(冯湘等,2020;Zhu et al.,2016;Guo et al.,2011)。封育后经过长时间积累,群落种内及种间对资源、空间的竞争逐步加强,导致生境相对较差的斑块植物繁殖与发育受到影响,造成区域群落特征指标和土壤理化性质的波动变化(刘文亭,2018;刘小丹等,2015),可能引起植被群落的再度退化(Su et al.,2015)。
宁夏东部荒漠草原位于毛乌素沙地西南边缘,处于中国生态系统严重退化带,生态环境极为脆弱。近年来在日趋频繁的人类干扰活动驱使下,该地区荒漠草原正发生不同程度的退化,有研究指出目前该地区约有 96.92%的荒漠草原面临不同程度的沙漠化,其中重度沙漠化草地约占50%(唐庄生,2018)。现阶段对该地区草地的研究多集中草地植被动态变化过程(冯湘等,2020;刘小丹等,2015),而在不同封育方式下荒漠草原群落数量特征及土壤理化性质的研究相对较少。本研究以位于宁夏东部荒漠草原区的固定监测样地为研究对象,通过连续3年的调查,分析物种多样性、群落稳定性及地上生物量关系及土壤理化性质等指标动态,旨在探讨:(1)不同封育时间和封育措施下植被群落数量特征的差异如何,各个数量特征之间是否具有相关性;(2)不同封育方式下土壤理化性质的会发生怎样的动态变化。研究结果可为中国中西部干旱、半干旱地区退化植被恢复与重建过程中生态系统的可持续经营管理与利用提供理论指导。
1 研究方法
1.1 研究区概况
本研究所选样地位于宁夏盐池县柳杨堡人工封育区(37°50′N,107°24′E)。该地区常年干旱少雨,日照强烈,气候干燥,属于典型的温带大陆性气候。年均气温为8.1 ℃,极端最高均温为34.9 ℃,极端最低温为-24.2 ℃。年均蒸发量多达2024 mm,年均无霜期约为165 d。年均降水量仅为285.5 mm,且降水季节性分配不均,超过 80%的降雨发生在6—9月。在1955—2015年这60年间降雨量有逐渐变小的趋势(图1a),在2013—2015年降水总量和季节性分配差异较大(图1b),2013年降水总量(288.2 mm)和多年平均值(285.5 mm)基本相同,为平均降水年;而2014年(346.6 mm)和2015年(365.6 mm)降雨量显著高于多年平均降雨量,为丰水年。土壤类型主要为灰钙土,其次分布有基岩风化沉积土和风沙土。植物区系属于欧亚草原区亚洲中部亚区,是中国中部草原区的过渡地带,以旱生和中旱生草本植物为主。
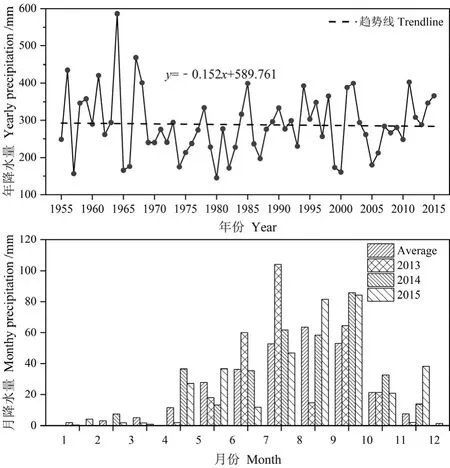
图1 盐池县1955—2015各年降雨量(a)和2013—2015年各月降雨量(b)变化Fig. 1 Variation of annual average precipitation from 1955-2015 (a) and monthly precipitation from 2013-2015 (b) in Yanchi County
1.2 样地布设与调查
本研究所选草地位于地势较为平坦的连续地段,生境梯度差异较小,环境条件相对均质,这有效降低了空间异质性对本研究的潜在干扰。因此本文基于“空间代时间”方法,依据草地封禁时间及封禁方式不同选择4个草地,即22年完全封育草地(LG)、11年完全封育草地(FG)、11年生长季封育草地(SG)和自由放牧草地(CG),每块草地大小约为20 hm2(500 m×400 m)。其中LG和FG分别自1991年和2002年开始实施完全封禁,方法是草地外围布设围栏,以杜绝外界牲畜干扰;SG自2002年开始进行封育,但在生长季末期,即在秋冬季节在草地内进行自由放牧,载畜率约为0.75只·hm-2·a-1,为该地区较适宜放牧强度(刘王锁等,2008);CG未对样地做任何保护措施,完全受当地传统放牧的影响,载畜率为 1只·hm-2·a-1,属于中度偏重度放牧强度(刘王锁等,2008)。
在2013—2015年生长季旺季(8月中上旬)对样地进行植被调查,方法是在每块样地随机布设15个2 m×2 m的小样方,通过利用卷尺多次测量植物自然高度,取平均值的方法测定植物高度;利用针刺法测定植被盖度;利用计数法统计,按科分类,分种记录的方法测定植物密度;利用枝剪齐地刈割,分种称重,测定植物地上生物量。并在2015年在每个样地随机选择5个点进行取土,取土深度为0—30 cm,分为两部分。一部分带回实验室自然风干并过筛(2 mm)后,用凯氏定氮法测定土壤全氮,用硫酸-高氯酸消煮法测定全磷,用重铬酸钾氧化-外加热法测定有机质含量;另一部分用烘箱烘干后,用以测定土壤含水量。
1.3 数据分析
1.3.1 重要值测度
本文将相对生物量也加入重要值的计算公式中,利用5种相对指标的平均值计算各个物种的重要值:

1.3.2 α多样性测度
本研究选取 Gleason丰富度指数、Shannon-Wiener优势度指数、Simpson多样性指数、均匀度指数等α多样性指标来指示研究区内物种多样性水平,其计算公式如下:
(1)Gleason物种丰富度计算:

(2)Shannon-Wiener指数:

(3)Simpson多样性指数:

(4)群落均匀度指数(Species evenness):

式中:S为调查样方内植物种总数;A为样方面积;Pi为第i个植物种的相对重要值;H为Shannon-Wiener指数。Pi=Ni/N;Ni为调查样方中第i种物种的重要值;N为调查样方内所有植物种的重要值和。
1.3.3 M. Godron稳定性指数
M. Godron稳定性指数采用贡献定律法(Godron,1972)。其原理是将样地内出现植物种的频度先按照从大到小的顺序排列,换算成相对频度后按物种排列顺序逐步累积得到相对频度的累积百分数,然后将植物种数的倒数百分数同累积相对频度一一对应,以植物种类倒数百分数为x轴,累积相对频度为y轴做散点图,采用数学模型的方法对散点图进行平滑曲线模拟。该曲线与y=100-x这条直线的交点即为所求群落的稳定性交点,即为欧式平方距离,所求交点坐标见表1。一般认为(20,80)坐标点为群落的稳定点,计算出的交点坐标越趋近于(20,80)表明群落就越稳定,反之,越不稳定(郑元润,2000)。

表1 稳定性分析结果Table The analysis result of community stability
数学模型公式为:y=ax2+bx+c
将直线y=100-x带入公式得出:
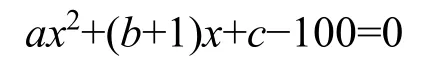
得到坐标为:
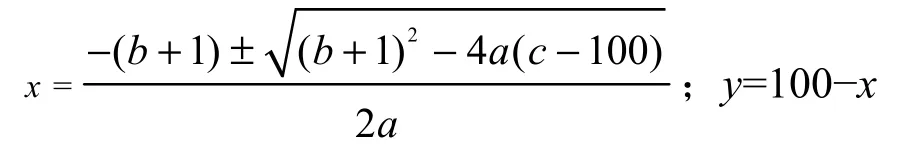
横、纵坐标都为正值的点即为所求稳定性交点。
用Excel 2013软件进行数据的汇总分析与预处理,用单因素方差分析(One-Way ANOVA)和最小显著差异法(LSD)检验不同样地对地上植被指数及土壤理化性质的差异性影响,用Pearson相关性分析定量分析各群落特征值之间的相关关系。
2 结果与分析
2.1 不同生活型植物地上生物量年际变化
由图2可以看出不同样地各年的地上多年生植物生物量(PH)及总生物量(T)均表现为FG>LG>SG>CG样地,一年生植物(AH)生物量表现为SG>LG>FG>CG样地,且丰水年(2014年和2015年)各样地总生物量均高于平水年(2013年),而灌木(S)生物量表现为LG>FG>CG>SG样地。在调查年内3种封育草地(LG、FG、SG)的AH、PH及T地上生物量显著高于(P<0.05)自由放牧草地(CG),且均在2014年最高,而SG样地的灌木生物量在调查年内均最低。且PH地上生物量约占总生物量的一半,AH的生物量比重较小,一般低于20%。
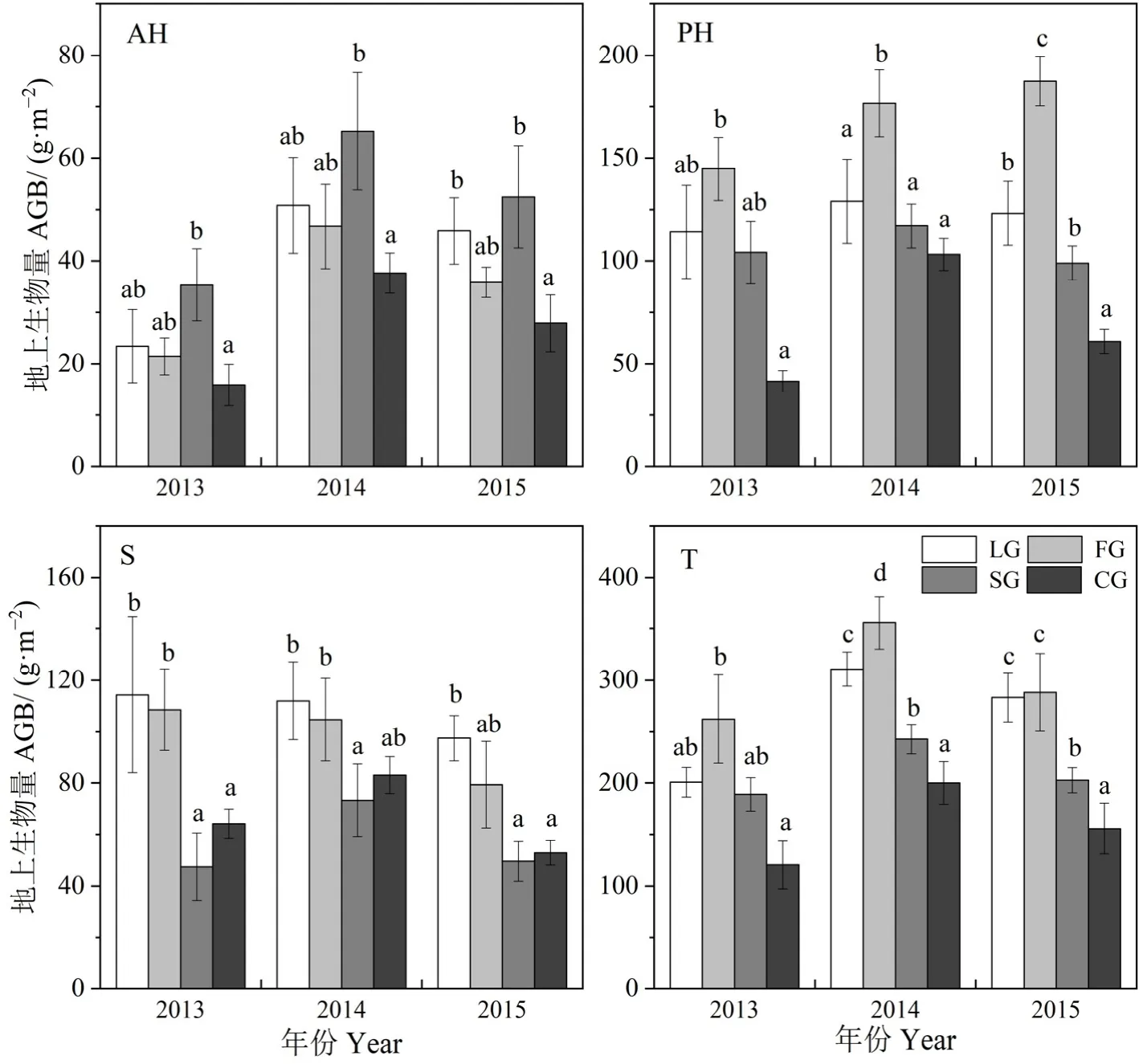
图2 各封育管理方式草地不同生活型植物地上生物量Fig. 2 The aboveground biomass (AGB) of different life-form plants in different fencing regime grasslands
2.2 物种多样性指数
图3结果显示,FG和 SG样地的 Shannon-Wiener指数(SW)、Simpson指数(SP)和均匀度指数(E)显著高于LG和CG样地(P<0.05),而其丰富度指数(R)则显著低于LG和CG样地(P<0.05)。有封育样地(LG、FG、SG)的SW指数显著高于自由放牧样地(CG)(P<0.05)。各个样地中,SG样地的SW、E和SP指数在各个调查年内均最高,而其R指数最低。2014及2015年各样SW指数高于2013年,而其E指数低于2013年,SP指数除在SG样地外与表现为2013年大于2014年和2015年,而R指数除在FG样地均表现为2013年大于2014及2015年。
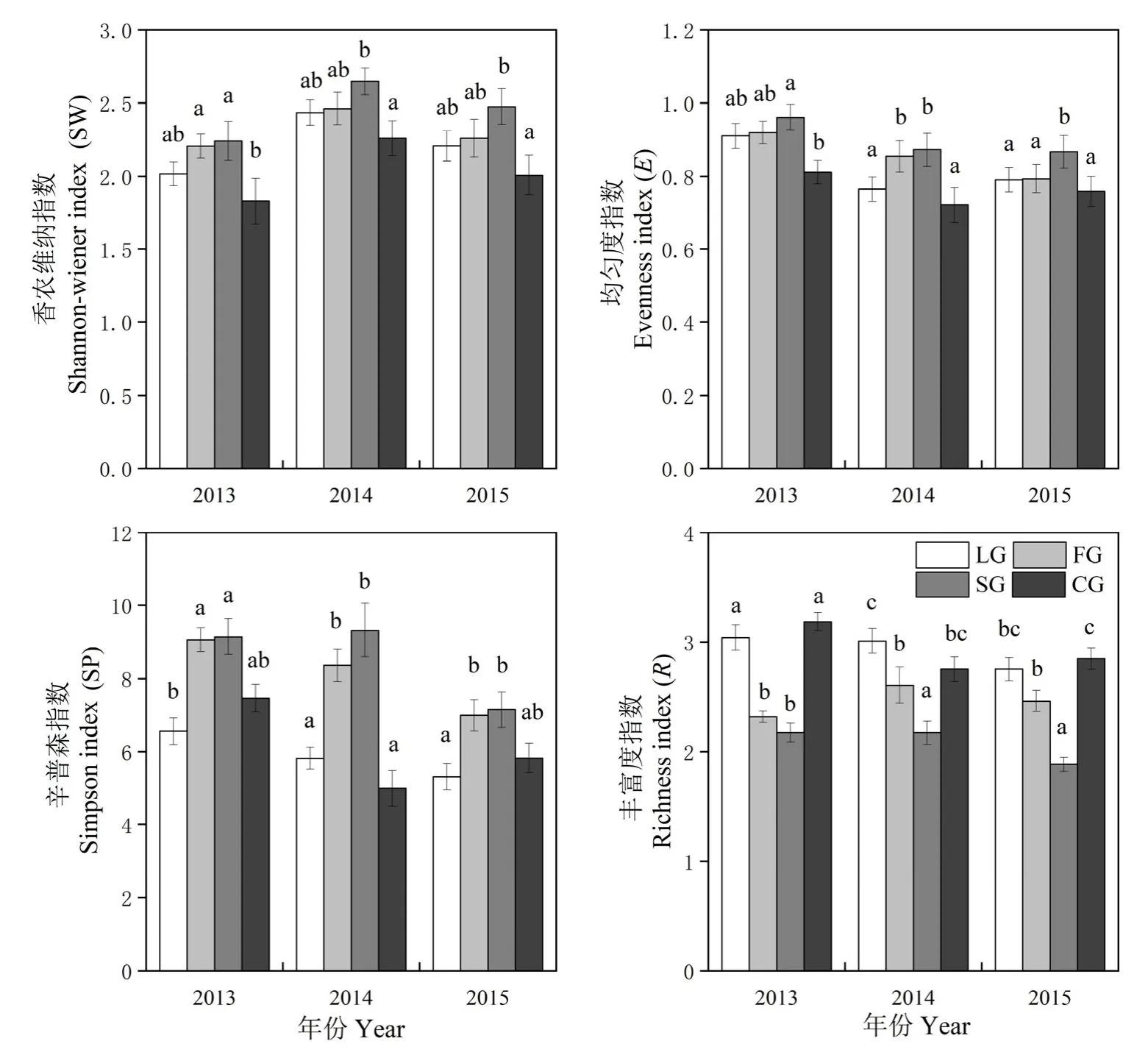
图3 不同封育管理方式草地α多样性指数Fig. 3 The α diversity index in different fencing regime grasslands
2.3 M. Godron稳定性指数
根据 M.·Godron稳定性测定方法,利用 Excel将群落所有植物的倒数百分数同累积相对频度进行曲线模拟,4个不同样地3年的稳定性模拟曲线拟合方程R2>0.99(表1),拟合效果极优,说明该地区样地植被分布规律很好的符合M·Godron稳定性测定方法。通过计算拟合曲线与直线Y=1-X的交点,得出样地的稳定性交点坐标和欧式平方距离(表1),欧式平方距离可直观反映群落稳定性。表1显示,在 2013—2015年欧式平方距离均表现为LG<FG<SG<CG,这表明封育样地(LG、FG和SG)群落稳定性高于放牧样地(CG)。欧式平方距离最大值出现在2015年CG样地,其欧式平方距离为25.96,比2014年增加2.53。最小值出现在2015年LG样地,为21.30。且不同样地群落稳定性对年际间环境变化的响应有所差异,在3调查年内FG的稳定性逐年增强,样地环境正在逐步改善,在2015年其欧式平方距离已同LG样地基本相同,表明经过 13年封育后群落稳定性已接近该地区的峰值,之后继续封育已不能提高群落稳定性,反而使其处于波动变化中。
2.4 各植被特征值的相关性分析
为进一步了解群落多样性、稳定性和生产力之间的关系,将各个群落特征值指标进行相关性分析。结果显示(表2),欧式平方距离和地上生物量呈显著负相关(P<0.05),相关系数r为0.652,这说明地上生物量越高群落稳定性越强,而与各 α多样性之间相关性不显著(P>0.05)。SW多样性指数与地上生物量、丰富度指数R之间相关性显著(P<0.05),而均匀度指数E与SP多样性指数在0.01水平上极显著相关(P<0.01),Pearson相关系数为0.793。
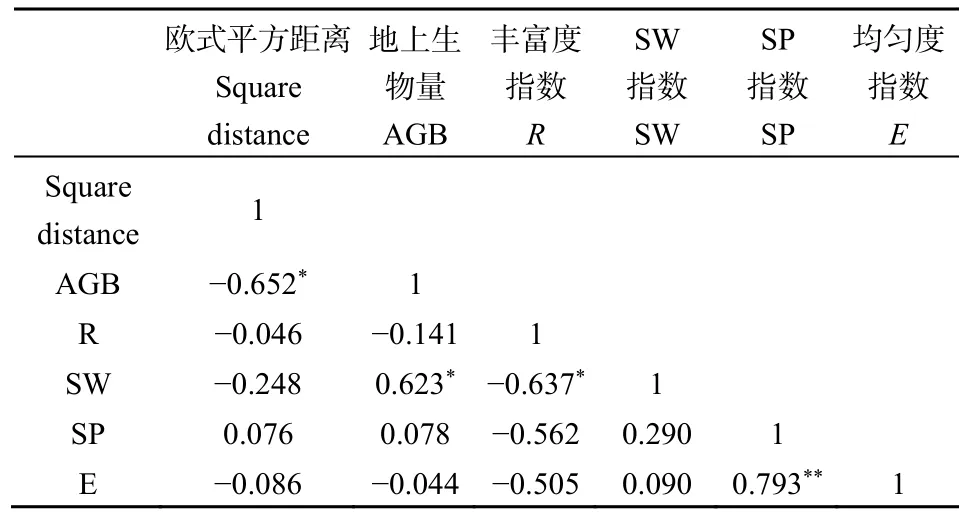
表2 群落特征值相关性分析Table 2 Correlation analysis of different community characteristic
2.5 土壤理化性质
表3结果显示,4个不同样地间土壤含水量、全氮、全磷及有机质含量差异显著(P<0.05),土壤全氮、全磷和有机质均表现为FG>SG>LG>CG,而土壤含水量的表现趋势为LG<FG<SG<CG。其中土壤全氮、全磷和有机质含量分别由 FG样地的115.92 mg·kg-1、234.28 mg·kg-1和 5.48 g·kg-1逐渐减为 CG 样地的 78.13 mg·kg-1、111.41 mg·kg-1、3.48 g·kg-1。而土壤含水量则由LG样地的2.17%逐渐增加到CG样地的4.48%。
3 讨论
围栏封育是退化草场恢复的重要措施之一,它可通过减少物质、能量从土壤-植被生态系统向牲畜的流动过程,改变群落结构,从而加速生态系统恢复(Zhu et al.,2016;刘小丹等,2015)。围封后草地受人畜干扰消除或降低,这为植物快速生长发育创造了有利条件,使得原有生态系统内因牲畜竞争排斥造成的生态位空缺现象逐渐被快速生长的植物占据(Li et al.,2017)。本研究指出草地围栏封育后群落地上生物量、多样性、稳定性等群落特征值显著提高,这与在国内外其它学者的研究一致(聂莹莹等,2020;Liu et al.,2019;Mdeina-Roldan et al.,2012;王黎黎等,2011)。
封育方式是影响植被恢复效果的重要方面,特别是在干旱、半干旱地区围封的效果主要取决于围封时间(Angassa et al.,2010)。王蕾等(2012)通过研究发现封育后短时间内群落各特征指标变化最为显著,之后其波动幅度逐渐减缓,然而由于区域土壤养分、水分等自然资源的有限性,随着封育时间的延长植株个体占据的资源位逐渐减小,植物生长受到影响,导致长期封育后群落各特征指标逐渐降低(Cheng et al.,2016;单贵莲等,2009)。生物量作为衡量群落生产力及潜在生产力的重要参数,可直观反映植被生长状况。本研究指出封育对草地植被恢复有积极的影响,草地围封后短时间内(11—13 a)地上生物量显著提高,长时间封育(22—24 a)降低了地上生物量,这与 Cheng et al.(2016)的研究结果一致。物种多样性可表征植物群落物种丰富程度、组成结构特征、生境差异、发展阶段和种群分布均匀程度等,是研究群落结构水平的重要指标(韩国栋等,1999;宋永昌,200147-48),而群落稳定性可代表植被本身稳定性特征及规律,是衡量群落结构、功能的一个综合特征(高润梅等,2012),两者均具有重要的生态学意义。本研究指出,长时间完全封育样地SW、E、SP指数低于,而其丰富度指数R和群落稳定性指数高于短时间封育样地。相关性研究表明群落稳定性与多样性的相关性不显著,而与地上生物量呈显著相关,这与Sasaki et al.(2011)的研究结果相同。这说明封育后随着地上生产力的逐步改善,群落稳定性逐渐增强,但在封育一定时间后随着种内、种间竞争的加剧,各个群落特征指标及群落多样性指数的下降,群落稳定性也随之发生变化(Liu et al.,2017;单贵莲等,2009)。
退化草地生态系统修复是植被-土壤间相互作用、共同恢复的复杂生态过程(Tao et al.,2013),土壤作为植被生长、发育和繁衍生息赖以生存、发展的基础,土壤性状波动是反映土壤营养环境的优劣、植被退化或恢复的重要指标(Inderjit,2005)。植被恢复过程中植物与土壤之间存在反馈机制,大量研究指出植被的恢复是土壤肥力恢复的前提条件,而土壤的恢复又反作用于植被群落(Liu et al.,2019;Wang et al.,2019)。本研究指出封育草地各土壤养分指标显著高于自由放牧草地,这一方面是因为封育后随着地上植被逐步恢复降低了近地面风速,土壤受风蚀状况降低;而植被对降雨的拦截作用削弱了土壤水蚀,减少了土壤养分流失,加速了土壤性状恢复过程,土壤养分水平逐渐提高(Mekuria et al.,2009;Zhou et al.,2011)。另一方面是随着地面植被的大量生长,植物对空气降尘拦截量增加,改变了土壤机械组成;加之地下根系对土壤结构的破坏作用,削弱了土壤板结,促进了土壤养分循环与运输(Wang et al.,2019),引起土壤质地逐步发生变化,造成土壤有机质等含量升高(Zhou et al.,2011)。此外,植被的生长发育将大量营养物质固定在植物组织中,减少了营养物质外流,并以凋落物、菌根、死根或根系分泌物渗出物的形式进入土壤,随着其逐步分解促进土壤发育和演变(Prietzel et al.,2012)。以上这种植物-土壤间复杂交互作用机理最终造成封育后土壤全氮、全磷和有机质含量显著提高。
水分是植物生长发育的必要条件(张继义等,2010),特别在气候干旱的西北内陆地区由于日照强烈,水分蒸发量较大,土壤水分含量对植物的作用尤为显著。有研究表明,多年封育后随着植被逐渐恢复,植物的吸水及耗水特性致使土壤水分条件逐步恶化(张继义等,2010)。此外随着草地封禁时间延长,由于缺乏干扰枯落物逐渐积累及土壤结皮逐步发育,在吸收利用有限水分的同时,也阻碍了降水的下渗,减少了土壤水分下渗量(刘小丹等,2015;张定海等,2017)。土壤水分的降低影响了土壤与植物间物质能量循环,土壤养分无法得到有效更新,抑制了植物的生长发育,不利于植物群落繁殖更新,最终导致群落多样性和生产力的下降(苗福泓,2015;杨婧等,2014;张定海等,2017)。而秋冬季放牧过程中牲畜的踩踏活动破坏了地表生物结皮,促进了降水的入渗作用,且牲畜的采食、排便过程使得能量流动加快,导致土壤全氮、全磷含量和有机质含量降低,土壤养分条件的改变反作用于群落结构(Li et al.,2016;Harris et al.,2007),致使群落特征值发生变化。
4 结论
围栏封禁是一种近自然恢复的生态修复手段,其短期目的是恢复退化植被,长远目标是通过人为干扰维持草地生态系统平衡。本研究发现不同封育方式对草地群落结构和土壤产生差异化影响不同,长时间完全封育不利于生物多样性保护,引起草地地上生物量、群落稳定性、土壤理化性质降低等问题。而生长季封育草地其地上生物量、群落多样性、群落稳定性及土壤理化指标均高于长期完全封育草地,因此建议在草地封禁适度时间后可对其进行适度冬季放牧利用,在维持草地生态系统稳定性的同时,也可达到牧民增收的目的。
