不同光照强度对四季金花茶光合生理特性的影响
2021-04-07杨运源黄永芳吕铭滔龚勇军
杨运源,邓 征,黄永芳,吕铭滔,龚勇军
(华南农业大学林学与风景园林学院,广东 广州 510642)
四季金花茶(Camellia perpetua)为山茶科山茶属常绿灌木或小乔木,叶片革质偏薄,基部呈圆形、近圆形或宽楔形,种子有短绒毛[1—2]。花朵呈淡黄色或金黄色,观赏价值极高,是上千个茶花品种中稀有的黄色花品种[3],并具有重要的保健作用和营养价值[4]。四季金花茶是目前发现的唯一能四季开花的金花茶,花期主要集中在夏季[5]。野生四季金花茶资源稀缺,主要分布在广西,少数分布在贵州、云南等地[6]。四季金花茶大都生长在温暖湿润、半阴半阳等地域,如山沟、林下、山边等阴凉湿润小环境,属于阴生植物[7]。
不同的光照强度会影响金花茶叶片形状,在低光照强度下金花茶更容易形成短窄型叶片,在高光照强度下则倾向于形成长阔型叶片[8]。四季金花茶种植在一定郁闭度的落叶林下,春夏季可得到林木茂盛枝叶遮挡部分光照,秋冬季林木落叶后有充足的阳光照射[9]。然而,大量四季金花茶原始生长区域植被遭砍伐,导致其适宜的光照环境被破坏,影响四季金花茶种群繁育和保护[10]。本研究设置不同光照强度模拟四季金花茶的生长光照环境,分析其光合生理特性变化,旨在为四季金花茶保育中提供适宜的光照条件作参考。
1 材料与方法
1.1 试验地概况
实验于华南农业大学苗圃园中进行,该地处在广东省广州市天河区(113°18′E、20°06′N),天河区位于北回归线以南,属于亚热带季风气候,夏季高温多雨,冬季温暖少雨。近二十年来,最高气温36.6 ℃,出现在8月,最低温度在1月,年平均温度21.8 ℃。近二十年来年均降雨量逐渐减少,年均降水量1623.6~1899.8 mm,4~9月降水占全年的82%,年平均相对湿度可达79%。
1.2 材料
采用2年生四季金花茶扦插苗,选取生长状况良好、长势相似且无病虫害的苗木进行实验。
1.3 试验设计
四季金花茶扦插苗种植于直径30 cm、高30 cm花盆,栽培基质为黄心土、泥炭土、蛭石按照体积比5:4:1混合配置。利用不同厚度遮阳网遮阴,光照强度分别设 10%光照、20%光照、40%光照和全光照共四个处理。光照强度设置参考柴胜丰等[11—12]及曾祥艳等[13]的方法。每种光照强度处理栽培20株,其他管理措施相同,待苗木适应生长环境45 d后测定相关指标。

1.4 指标测定
1.4.1 光合特性指标
在晴天无风的早上9:00~11:00,每处理随机选取3株生长状况优良的苗木,每株生长点往下第3~5片展开叶为材料,使用Li-6400便携式光合作用测定仪测定净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci),每株测定3片叶,每个样品重复 3次。测定时选用红蓝光叶室,光照强度800 µmol·m-2·s-1,CO2浓度 350 μmol·m-2·s-1,叶室温度 26 ℃[14—16]。
1.4.2 生理特性指标
叶绿素含量测定参照陈建勋等[17]的方法,可溶性蛋白含量测定采用考马斯亮蓝G-250染色法[18],可溶性糖含量测定采用蒽酮法[19];超氧化物歧化酶(SOD)活性测定采用氮蓝四唑光化还原法[20],过氧化物酶(POD)活性测定采用愈创木酚比色法[20],丙二醛(MDA)含量测定采用硫代巴比妥酸法[20]。
1.5 数据处理
使用Microsoft Excel 2010作图,用SPSS 22.0进行数据分析。
2 结果与分析
2.1 不同光照强度下四季金花茶光合特性变化
不同遮光处理下四季金花茶叶片净光合速率总体表现为随光照增强而降低(图1: A)。其中,10%光照、20%光照和 40%光照三个光照处理对四季金花茶净光合速率的影响不显著,全光照处理下四季金花茶净光合速率与其他三个光照处理差异显著(P<0.05),说明全光照处理对四季金花茶光合作用产生抑制。
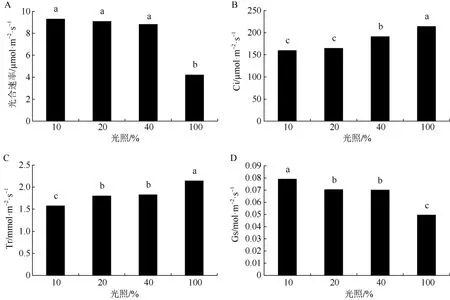
图1 不同光强对四季金花茶光合作用的影响Fig. 1 Effect of different light intensity on photosynthesis of Camellia perpetua
在不同光照条件下,四季金花茶叶片胞间 CO2浓度总体上随光照增强而升高,即胞间CO2被吸收减少,说明在实验设置的光照强度范围内,随着光照增强四季金花茶光合作用减弱(图1: B)。胞间CO2浓度在全光照处理下显著高于其他三种光照处理(P<0.05),40%光照处理的胞间 CO2浓度显著高于20%光照处理和10%光照处理(P<0.05),但20%光照处理和10%光照处理之间无显著差异。
如图1: C所示,四季金花茶叶片蒸腾速率总体随光照强度增大而加快。全光照处理下四季金花茶蒸腾速率显著高于另三种光照处理(P<0.05),而40%光照处理和20%光照处理之间差异不显著,但两者与10%光照处理差异显著(P<0.05)。
四季金花茶叶片气孔导度大致随着光照增强而递减(图1: D),其最大值在10%光照处理,分析其原因,可能是光照增强导致植物光合作用减弱,消耗CO2减少,因此气孔导度降低,减少空气中CO2进入细胞。10%光照、20%光照和40%光照处理下植株气孔导度显著高于全光照处理(P<0.05)。20%光照处理和 40%光照处理显著低于 10%光照处理(P<0.05),但两者之间无明显差异。
2.2 不同光照强度下四季金花茶的生理特性
如图2: A所示,四季金花茶叶绿素含量在四种光照处理下呈现出不同的变化。在40%光照下生长的四季金花茶叶片叶绿素 a含量最高,为1.35 mg·g-1FW,且与其他三个光照处理差异显著(P<0.05),其他三个处理之间差异不明显。叶绿素b含量以40%光照处理为最高(0.92 mg·g-1FW),20%光照处理的最低(0.03 mg·g-1FW),10%光照和全光照处理之间差异不显著。叶绿素总量最高值出现在40%光照处理,为2.66 mg·g-1FW,最低值在20%光照处理,为0.96 mg·g-1FW,10%光照处理和全光照处理之间差异不明显,其他光照处理之间差异显著(P<0.05)。

图2 不同光强对四季金花茶生理特性的影响Fig. 2 Effects of different light intensity on physiological characteristics of Camellia perpetua
不同光照强度下四季金花茶可溶性蛋白含量总体上随光照强度增大而降低,即10%光照处理>20%光照处理>40%光照处理>全光照处理(图2: B)。10%光照处理下含量最高,为1.42 mg·g-1FW,全光照的含量最低,为0.15 mg·g-1FW。在四个光照强度处理下的四季金花茶可溶性蛋白含量差异明显(P<0.05),说明植株可溶性蛋白含量受光照强度影响较大,对光强反应较敏感。
四季金花茶在四种光照处理下可溶性糖含量出现两个相对高点和两个相对低点,最高值为全光照下的7.54%,最低值10%光照下的3.5%(图2: C)。按照可溶性糖含量高低排序四个处理为全光照>20%光照>40%光照>10%光照,不同处理之间可溶性糖含量差异明显(P<0.05)。
如图2: D所示,不同光照强度处理的四季金花茶超氧化物歧化酶(SOD)活性随光照增强而加大。全光照的四季金花茶叶片 SOD活性最高,为324.27 U·g-1FW,10%光照处理的最低,为280.74 U·g-1FW。全光照和40%光照处理的四季金花茶 SOD活性无显著差异,但两者与 10%光照和20%光照处理差异显著(P<0.05)。
在不同光强下四季金花茶叶片过氧化物酶(POD)活性也不同,总体表现为随光照增强而降低(图2: E)。10%光照和20%光照处理的四季金花茶叶片POD活性显著高于其他两个处理(P<0.05),全光照处理的POD活性最低,为0.56 U·g-1FW。
四季金花茶叶片丙二醛(MDA)含量在10%光照处理下最低,显著低于其他三个处理(P<0.05);当光照高于20%时,MDA含量没有显著变化,可能是超过20%光照后植株已达到光饱和点,过多的光照对植株产生胁迫作用。
3 结论与讨论
光照强度是决定植株生长水平的重要因素[21]。何小燕等[22]发现,在光强过高时,油茶(Camellia oleifera)叶片净光合速率降低,气孔导度降低,以减轻高温高光对植物叶片的伤害。刘慧民等[23]研究发现,绣线菊(Spiraea salicifolia)在高温和强光下叶片蒸气压增加,叶片温度升高,蒸腾速率加快,叶片细胞失水,水势下降,气孔导度下降,净光合速率下降。黄丽英等[24]研究发现,净光合速率与气孔导度成正相关,与胞间CO2浓度成呈负相关。本研究中,四季金花茶在不同光照处理下,净光合速率、胞间CO2浓度、蒸腾速率、气孔导度均发生改变,其中净光合速率与气孔导度随光照增强而下降,两者之间呈现正相关,胞间CO2浓度与蒸腾速率随光照增强而提高,两者之间呈现正相关,与净光合速率和气孔导度呈现负相关,与前人研究基本一致。10%、20%和 40%光照处理对四季金花茶净光合速率影响不明显,说明四季金花茶适应光强的范围较广,本实验遮阴设置还未达到抑制净光合速率,在全光照处理下净光合速率显著降低,表明全光照对四季金花茶净光合速率产生抑制作用。胞间CO2浓度、蒸腾速率分别在 40%、20%光照下显著提高,气孔导度在 20%光照中显著降低,表明三者在10%~40%光照中的显著变化不影响净光合速率。四季金花茶是阴生植物,无法在全光照下及时消散剩余光能,受到光氧化作用损害,致使PSⅡ反应中心受到伤害,这可能是四季金花茶光合速率下降的主要原因。
叶绿素含量在一定程度上反映植株光合效率强弱[25],光照强度影响叶绿素的合成和分解,叶绿素在低光照下更易合成,强光容易使叶绿素分解。闫小莉等[26]研究表明,苦丁茶树(Ligustrum robustum)在适当遮阴处理下有利于产生更多的叶绿素a和叶绿素 b,耐阴植物可利用低光照进行光合作用保证正常生长。叶绿素含量随光照强度增加而减少[27—28]。四季金花茶叶绿素a、叶绿素b和叶绿素总量最大值均在40%光照处理,与前人研究有所区别,说明叶绿素含量不仅受光强影响,还受植物外部其他因素或内部因素影响。10%、20%和 40%光照下叶绿素含量的变化对植株净光合速率影响不明显。
可溶性蛋白在植物体内参与大部分代谢活动,包含重要的酶类,可作为植物总代谢的参考指标,在极端环境中能产生新的蛋白[29]。桂克印[30]研究发现,绿萝(Epipremnum aureum)在适宜的光照范围,蛋白质含量随光强增大而升高,当光强过高时,限制了植株代谢活动,蛋白质含量降低。四季金花茶可溶性蛋白含量随光强增大而降低,光照过强可能对其代谢产生抑制,植株无法产生新的蛋白抵抗逆境,此时植株的抗性较低,更容易感染病毒虫害或受其他胁迫影响,降低植株的生命力。因此,在栽植四季金花茶时应尽可能遮阴,减弱光照强度,更有利于其生长发育。
郭峰[31]研究发现,小麦(Triticum aestivum)在遮阴条件下叶片中可溶性糖含量显著下降,可溶性糖含量浓度较高时可能是由于同化物运输能力减弱,导致同化物积累在叶片中抑制暗反应正常进行,从而净光合速率降低。四季金花茶可溶性糖含量在全光照下最高,此时植株受到光抑制,同化物传输受阻,导致可溶性糖含量增加,说明四季金花茶适于低光照环境。
超氧化物歧化酶(SOD)可以在植物受到逆境胁迫时清除其细胞内过多的自由基,是植物重要的保护酶,使细胞能够抵制逆境胁迫。粟春青等[13]研究发现,金花茶(Camellia petelotii)叶片细胞内活性氧增加,SOD活性也增加,说明SOD能够维持活性氧自由基的体内平衡,对保护细胞发挥重要作用。四季金花茶SOD活性随光照增强而提高,表明强光照对四季金花茶产生一定的胁迫,植株通过提高SOD活性消除强光照对细胞的伤害,与前人研究结果相似。在 40%光照和全光照下四季金花茶叶片SOD活性差异不明显,说明40%光照下SOD活性基本达最大值,随着光强的加大无法继续提高SOD活性。
过氧化物酶(POD)活性可反映植物抗逆性强弱,对催化植物细胞中的过氧化物和分解逆境下植物体产生的H2O2等有重要作用。一般POD等植物体内活性氧清除相关酶处在较高水平,当植株受到轻微胁迫时,POD活性增强,当胁迫程度超过一定水平时,细胞代谢失衡,POD活性降低[32]。四季金花茶叶片 POD活性随光照增强而降低,在 40%光照下POD活性显著降低,表明40%光照植株已受到一定的胁迫,继续加大光强可能对植株造成伤害,不利于四季金花茶的生长繁育和种群保护。
丙二醛(MDA)含量可反映植物的抗逆性。田肖箫等[33]研究发现,强光胁迫和弱光胁迫都会破坏闽楠(Phoebe bournei)幼苗叶绿体结构,致使其膜脂产生过氧化作用,进而影响植株光合速率,且 MDA含量随光强增大而提高。四季金花茶叶片 MDA含量在20%光照下接近最大值,与40%光照和全光照无显著差异,说明20%光照对其叶片细胞有一定的伤害,但净光合速率并未明显降低,可能是其他物质与MDA存在拮抗作用。
综上所述,不同光照强度对四季金花茶光合特性和生理特性产生不同的影响,各指标的变化趋势以及变化显著程度均有所差异。根据光合生理指标的动态变化,可见四季金花茶更适合种植在低光照环境中,增强光合作用,更利于植株生长发育,不宜种植在超过40%光照的环境中,应适当做遮阴处理有利于种群保护与人工栽培。
