苎麻纤维发育相关的3个miRNA鉴定与表达分析
2021-04-01马永红朱四元严理刘头明
马永红,朱四元,严理,刘头明
(中国农业科学院麻类研究所,湖南 长沙 410205)
小RNA(miRNA)是一段长22 bp左右、具有颈环结构的小分子非编码RNA,其在生物体内含量较rRNA、tRNA、mRNA等主要RNA要低得多。miRNA虽然不编码蛋白,但具有转录因子的作用,通过多种酶的剪切加工作用形成成熟的miRNA沉默复合体后,通过碱基互补配对靶向沉默mRNA,抑制靶基因的翻译或靶向降解靶基因,进而调控植物的生长发育[1-7]、逆境胁迫[8-9]等多种生物过程。例如miR319调控靶基因TCP4在拟南芥中的表达,miR319表达下调会使TCP表达量上升,促进了次生壁细胞的合成,筛管增厚[10]。在番茄中,miR319的过表达下调了几个TCPs基因,导致小叶变大,叶片边缘持续生长,而miR319表达水平的降低或TCP表达水平的提高会使番茄叶片变小。在柑橘中,miR397通过调控LAC基因调节细胞壁多糖的二次沉积,调控细胞壁的形成[11]。对于miRNA的研究引起生物学界广泛关注。
苎麻(BoehmerianiveaL.Gaud)是一种多年生草本植物,具有宿根性,纤维含量丰富,早期通过抽取纤维制作麻绳等,是中国主要的纺织纤维作物,具有悠久的种植历史。苎麻作为重要的天然纤维作物,其产量决定因素在于韧皮部,韧皮纤维次生壁的生物合成决定纤维产量。纤维素合成酶是植物纤维合成的关键酶,基于转录组序列,Liu等[12]发现苎麻中36个可能与韧皮纤维发育相关的纤维素合成酶基因。miRNA也可能参与了苎麻的韧皮纤维发育调控。Wang等[13]构建了苎麻纤维伸长、胞壁增厚和端壁溶解阶段miRNA表达谱,并在纤维伸长增厚和端壁溶解阶段识别到298个miRNA,推测他们可能与苎麻的纤维发育相关。在拟南芥、水稻、柑橘等中也有关于miRNA调控纤维发育功能的报道,如在拟南芥中,miR397b通过调控其靶基因AtLAC4的表达进而影响木质素的含量,miR397b过表达株系木质素含量降低,次生壁厚度变薄[14]。在陆地棉中,调控纤维发育的GhLAC4基因与miR397b的碱基互补区域发生了mRNA的切割,也说明了miRNA对棉花纤维发育的调控作用[15]。研究表明,具有较高同源性的基因在不同植物体中可能具有不同的生物学功能,例如mir397a在番茄中主要参与非生物胁迫过程[16],而在梨果肉中主要调控石细胞的发育形成[17]。在苎麻中关于纤维发育的miRNA研究较少,所以本研究拟针对苎麻茎皮进行miRNA测序,并进行表达分析,以研究和发现苎麻中可能与纤维发育相关的miRNA,旨在为阐明miRNA在苎麻纤维发育中的生物学功能奠定基础。
1 材料与方法
1.1 材料处理
苎麻材料为中苎1号,取自中国农业科学院麻类研究所试验基地,所取材料均经过清水冲洗,取叶(中部叶)、5芽(顶端芽)、顶皮(顶端韧皮部)、中皮(中间韧皮部)、茎(中间木质部)、根(根尖)6个部位迅速转入液氮后移至-80℃冰箱中保存备用,以防止RNA降解。
1.2 RNA提取及RNA纯度、产率和完整性检测
将0.5 g苎麻组织在液氮中迅速研磨至粉末状,按照植物RNA提取试剂盒说明书(Takara)的要求提取RNA,-80℃保存。苎麻根部含有大量多糖多酚类物质,在研磨样品后按说明书提取方法二(去除多糖多酚法)进行操作。
取1μL RNA样品,用微量分光光度计(NANO 100,中国)测其A260/A280和A260/A230的比值和RNA浓度。取所得的RNA约2μL,加2μL无RNAase的水和1μL 6×loading buffer后制备1.0%的琼脂凝胶,在1×TBE缓冲液中电泳检测总RNA的完整性。
1.3 反转录及qRT-PCR体系的建立
分别取苎麻6个部位RNA 1μg,按照cDNA合成试剂盒(TransgenAT311)上的说明书操作进行反转录。反应体系为 20μL,反应条件:30°C 10min,42°C 20min,95°C 5min,反转录完成后再加无菌水稀释5倍至100μL。反转录后的cDNA置于-80℃冰箱保存。
1.4 miRNA测序以及内参基因的选择
对中苎1号苎麻中韧皮部进行miRNA的提取、文库构建和高通量测序,对miRNA测得的原始数据需进行质量控制,得到高质量序列,为了识别miRNA,将miRNA序列中的优质数据分别采用AASRA[18]和 CMsearch软件[19]与 miRbase[20]和 Rfam[21]数据库进行比对。通过使用 PIPmiR[22]和默认参数,未匹配到这两个数据库的序列被认为是新的miRNAs。与已知miRNA数据库miRBase(v21)中的成熟miRNA序列进行比对,完全相同的序列被认为是已知miRNA。
所选内参基因要具有稳定表达的特性,且与要定量的基因表达量相近,为了获得可靠的qPCR结果,必须为每个研究物种和需要定量的基因选择合适的内参基因[23]。内参基因的稳定性取决于样本和条件,因为苎麻没有完善的qPCR内参设定体系,因此从常用作内参的管家基因中选择18s、ACT、TUB、UBQ4个基因。设计引物,凝胶电泳检测引物设计是否合适,然后进行qPCR检测。
在NCBI中查找测序鉴定到的候选基因的保守序列,用Primer 5.0软件设计引物(表1)。引物的退火温度为53~57°C,引物长度为18~21 bp,扩增产物长度为150~250 bp。引物序列由擎科生物有限公司合成,实时定量PCR使用的引物见下表1。
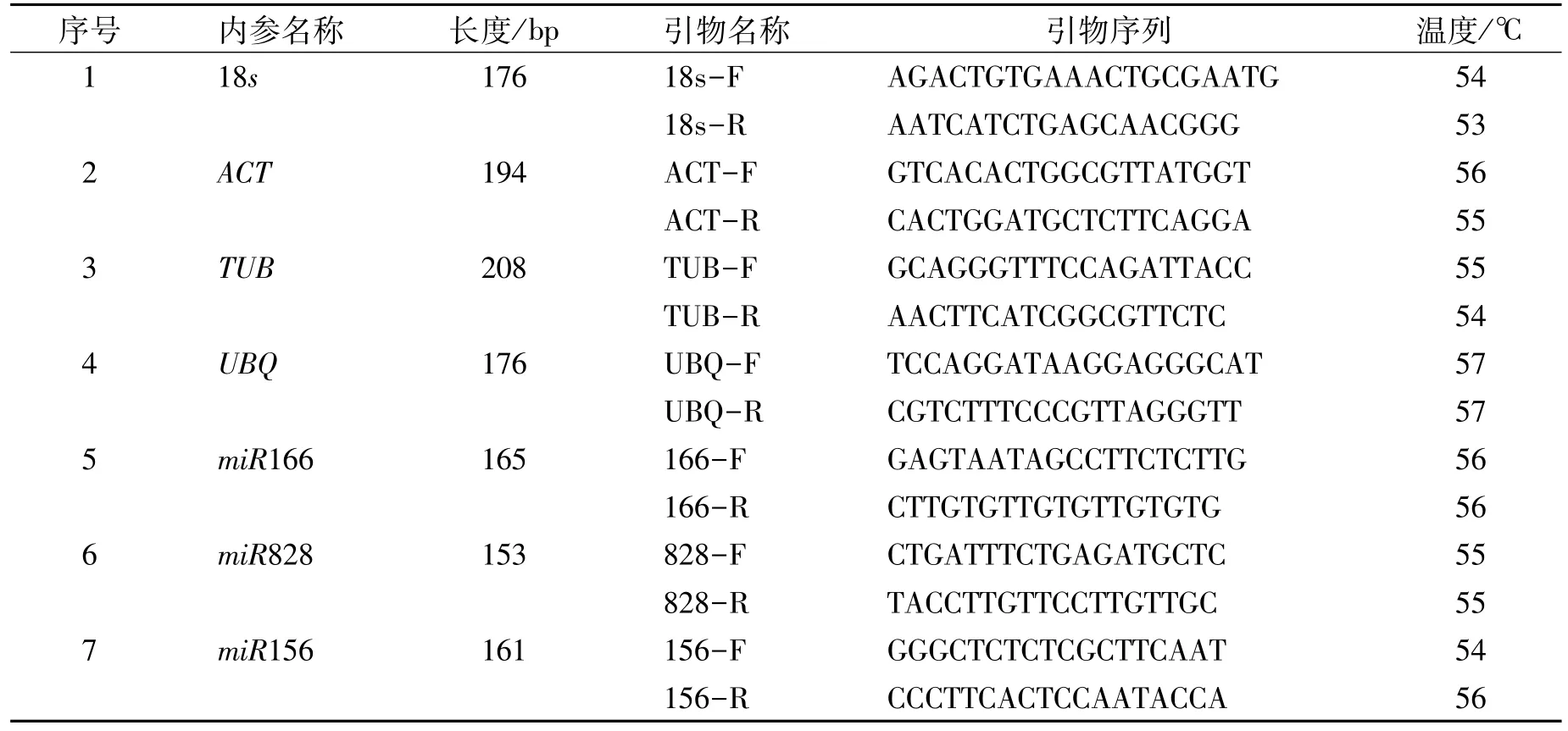
表1 qRT-PCR检测中基因的引物序列Table 1 Primer sequences of genes in qRT-PCR analysis
1.5 纤维发育相关miRNA的靶基因测定
miRNA为具有负调控作用的转录因子,其所调控的靶基因才可能具有实际调控纤维发育的作用,为了进一步阐明miRNA调控纤维发育的生物学机制,取苎麻韧皮部转录本进行测序,构建RNA文库并进行高通量测序,根据miRNA与靶基因碱基互补配对高度的保守性进行具有差异表达的miRNA的靶基因预测。常用的靶基因预测软件有TAPIR[24]和TargetFinder[25],参数设置为默认参数。
1.6 实时荧光定量
PCR试剂采用Transgen公司的荧光定量PCR试剂盒,采用SYBRGreen荧光染料法在CFX96实时荧光定量PCR仪上进行定量。按照说明书进行操作,选取管家基因18s为内参基因。PCR体系按照TB GreenPremix酶(Transgen公司)说明书的要求设置,对6个部位进行3次生物学重复,体系为20μL,扩增程序95℃预变性3 min,40个循环的程序为95℃变性30 s,62℃退火30 s,72℃延伸30 s。
1.7 数据处理
采用Excel进行数据统计和分析,CFX96分析软件制作扩增曲线。用BestKeeper软件分析内参基因表达的稳定性。相对表达量的计算以18s为内参基因,计算Ct值(Ct代表目标扩增产物达到设定阈值所经历的循环数),参照魏敏[26]计算方法,得到RQ值(表达量变化倍数)。即通过计算 ΔCt(ΔCt=Ct目的基因-Ct内参),获得 ΔΔCt(ΔΔCt=ΔCt(实验组)-ΔCt(对照组),再计算RQ值(RQ=2-ΔΔCt),取3次生物学重复的平均值作为该基因在该处理组的表达量,并计算RQ的误差。
2 结果与分析
2.1 苎麻中的miRNA测序
通过对苎麻韧皮部miRNA的测序,在苎麻韧皮部共计378个miRNA中,将测序序列进行注释,共鉴定出176个已知miRNA和202个新miRNA,为了研究miRNA在苎麻不同部位是否具有表达差异,选定了3个在模式生物中同源且与纤维发育相关的常见miRNA进行了实时定量PCR检验,分别为miR828、miR166和miR156。
2.2 纤维发育相关miRNA的靶基因预测
通过与拟南芥同源序列对比发现有许多和拟南芥一致的靶标,如HDZIP基因、MYB基因、NAC基因等,被miRNA(分别为miR166、miR828、miR156)靶向调控,具体见表2。而拟南芥中的HDZIP基因、MYB基因、NAC基因具有调控纤维发育的功能,由于miRNA具有反向调控靶基因表达的作用,通过抑制靶基因的表达调控植物生长发育。在拟南芥纤维发育过程中,茎顶端miRNA基因的表达量高,从而抑制靶基因的程度也高,在茎中部,miRNA基因表达量降低,使靶基因表达量上升,从而引起次生壁细胞发生增厚或增多,茎秆增粗,纤维量增加[27]。推测在苎麻中miRNA表达也具有这种规律性。

表2 miRNA靶基因功能注释Table 2 Functional annotation ofmiRNA target genes
2.3 苎麻内参基因的分析
对4对候选内参基因18s、ACT、TUB和UBQ进行测试,Ct值在6~37,表达丰度分布范围较广(图1)。由此可初步判断,18s和ACT表达较为稳定,TUB和UBQ表达差异较大。通过比较两个基因的变异系数(coefficient of variation,CV)和平均标准偏差(standard deviation,SD)来确定基因的稳定性[28]。一般稳定的内参基因具有较低的SD值和CV值。而SD的临界值为1.5,若SD>1.5,则表明稳定性较差[29]。但是在对目标基因进行校准时,不仅要求所用内参基因的表达水平稳定,而且要求其稳定的表达,与目标基因的表达量相近,表达量过高或过低,都可能导致定量结果出现明显偏差[30]。由表3可知,18s在不同部位中表达量较为稳定,为了保证试验的准确性,选定18s作为标准内参,取相对变量。
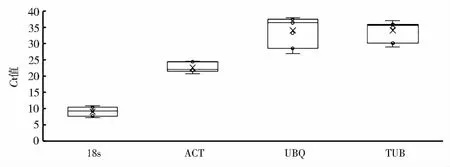
图1 苎麻内参基因的Ct值分布Fig.1 Distribution of Ct values of reference genes in ramie

表3 4个候选内参基因在苎麻不同部位的表达稳定性Table 3 The expression stability of 4 candidate internal reference genes in different tissues of ramie
2.4 miRNA中miR156、miR166和miR828的表达分析
对miR156、miR166和miR828进行定量分析,得到溶解度曲线见图2,采用表达量变化倍数(RQ值)对3个mirRNA基因在中苎1号不同部位的表达情况进行分析。以中苎1号根的表达量作为对照,设定表达量为1。由图3可知,miR828、miR166和miR156在顶皮和芽中表达量较高,miR828在芽中表达量最高,miR166其次,miR156最低。miR828、miR166和miR156在顶端韧皮部中表达量高,miR828表达量最高,miR156其次,miR166最低。miR828、miR166和miR156在苎麻的叶、茎和根中表达量均较低。
图2 苎麻中miR828、miR166和miR156扩增曲线Fig.2 Amplification curves of miR828,miR166 and miR156 in ramie

图3 候选基因在中苎1号不同部位的相对表达量Fig.3 The relative expression levels of candidate genes at different sites of Zhonghu No.1
2.5 纤维发育相关miRNA鉴定
将miRNA在顶皮和中皮两个部位进行定量表达见图4,由图4可以看出,3个miRNA在顶皮中的表达量要显著高于中皮,也进一步证明其与苎麻纤维发育相关。

图4 miRNA在顶皮和中皮的表达差异Fig.4 Expression differences of miRNA in the apical and mesothelial
苎麻顶端韧皮部与中间韧皮部中的纤维发育差异明显,中皮纤维处于生长期,次生壁处于正在加厚或增厚结束时期,而顶皮纤维处于刚开始生长期,纤维发育相关基因的表达必定活跃。
3 结论与讨论
本研究在苎麻中共检测到378个miRNA,其中包括176个已知miRNA和202个新miRNA。Wang等[31]对苎麻纤维发育茎皮中miRNA的表达谱进行了表征,得到51个具有差异表达的miRNA。可见,本研究中识别的miRNA数量上更加全面和丰富。在所选的4个管家基因中,18s在苎麻各个部位都能够稳定表达,建议在苎麻研究中作为内参基因。在测定的378个miRNA中选取的3个miRNA经实时定量PCR鉴定,在韧皮部顶端和中部具有差异表达。而顶端韧皮和中部韧皮主要存在纤维发育方面的差距。在模式植物拟南芥中miRNA具有调控纤维发育的作用,如miR319可以通过靶向调控TCP4转录因子激活次生壁的生物合成。miR319过表达导致茎中TCP4丰度降低和次生壁的形成,表明miR319介导TCP4发育过程中次生细胞的生物合成[32]。据此,推测苎麻中miRNA具有调控纤维发育的作用。
在拟南芥的纤维组织中,Yang等[33]发现MYB26过表达会引起木质素的异位沉积,导致表皮纤维组织次生增厚。并发现在两个NAC结构域基因NAC次生壁促因子1(NST1)和NST2的表达也发生了变化,这两个基因与花药内膜的次生增厚有关。NAC基因作为MYB26基因的上游基因,共同调控了纤维组织次生壁的增厚。拟南芥基因组包含47个HDZIP基因,具有调控纤维发育的作用。薛宇[34]比较不同发育时期棉纤维的表达谱深度测序数据,发现大量HD-Zip家族基因可能参与棉纤维发育过程。这说明MYB、NAC和HDZIP在纤维发育中具有重要调控作用。
在本研究中,通过生物信息学预测miR166、miR828和miR156分别靶向调控MYB、NAC和HDZIP基因。通过对比拟南芥同源序列发现,具有纤维发育相关的HDZIP基因、MYB基因、NAC基因的同源基因被miRNA(分别为miR166、miR828、miR156)靶向调控。一般来说,如果预测的靶基因具有调控苎麻纤维发育的作用,那么茎顶端miRNA基因的表达量高,从而抑制靶基因的程度也高,在茎中部,miRNA基因表达量降低,会使靶基因表达量上升,从而会引起次生壁细胞发生增厚或增多,茎秆增粗,导致苎麻纤维量增加。而HDZIP基因、MYB基因[35]和NAC基因在拟南芥等模式生物中对纤维发育的调控作用已被证实。MYB作为一类转录因子广泛参与木质部中木质素、细胞壁的合成。这说明miR166、miR828、miR156可能通过调控MYB、NAC和HDZIP基因来进一步参与到苎麻纤维发育的生物网络中去。本研究只对miRNA在苎麻不同部位的表达进行了定量分析,表明miRNA可能通过调控相关基因表达来调控苎麻纤维发育。对于以上推测需要对预测靶基因的表达量开展进一步的研究。因此,本研究为探究miRNA在苎麻纤维发育调控中的功能奠定了基础。
