LncRNA HOTAIR通过Wnt/β-Catenin信号通路调控NSCLC细胞增殖、迁移及侵袭
2021-03-06姚菲菲何爱萍
姚菲菲 王 军 何爱萍
肺癌(lung cancer)是发病率、致死率最高的恶性肿瘤之一,位于全球癌症致死率的首位,其不同类型中非小细胞肺癌(non small cell lung cancer,NSCLC)最为常见,占肺癌总量的85%左右[1]。因缺乏早期诊断的分子标记,多数患者确诊时已发展至中晚期,癌细胞发生了侵袭及转移。因此,开展肺癌发生发展的机制研究,找寻新的诊断及治疗靶点对肺癌的临床治疗具有重要意义。近年来研究发现[2-3],长链非编码RNA(LncRNA)通过多种途径参与基因表达的调控,多种LncRNA在肿瘤中均异常表达,可作为肿瘤潜在治疗靶点及预后标志物。最新研究发现HOX基因家族在胚胎发育过程中发挥重要作用,并通过鉴定分析于12号染色体的HOXC基因位点发现了HOTAIR[4]。据相关研究报道[5-6],HOTAIR在乳腺癌、宫颈癌等众多肿瘤中高表达,下调HOTAIR表达后可显著抑制肿瘤细胞的生长,加快其凋亡速率,且可增加化疗药物的敏感性,提示HOTAIR可作为恶性肿瘤临床诊断及治疗的新靶点。但目前有关HOTAIR在肺癌中的研究相对较少,因此本研究就LncRNA HOTAIR在NSCLC中的表达及作用机制进行了探究,以期为NSCLC的治疗及研究提供新的方向和目标。
1 材料与方法
1.1 实验细胞、试剂、仪器
实验所用人类非小细胞肺癌细胞系A549、H460、H1650及正常肺组织细胞HBE均购自中国科学院上海细胞库,于含10%胎牛血清的培养基中接种,37 ℃、5%CO2培养箱孵育培养。主要试剂及仪器:DMEM细胞培养基、RPMI1640培养基购自美国Gibco公司;胎牛血清(FBS)购自美国Hyclone实验室;胰蛋白酶消化液、LipofectamineTM2000转染试剂购自美国 Invitrogen公司;siRNA由GenePharma公司合成;β-actin购自美国Sigma公司;逆转录试剂盒、PCR Master Mix购自TAKARA有限公司;6/24/96细胞培养孔板、Transwell小室购自美国Corning公司;CCK-8试剂盒购自日本Dojindo公司;酶标仪、PCR仪购自美国 Bio-RAD公司。
1.2 方法
1.2.1 细胞复苏、培养、传代 细胞复苏与培养:于液氮中取出冻存管,37 ℃水浴箱溶解,离心弃上清加入含10%胎牛血清培养基重悬沉淀物,后加入含9 ml细胞培养基的培养皿中,37 ℃、5%CO2培养箱恒温培养,每隔2天更换培养基一次,密切观察细胞生长情况。细胞传代:待细胞生长密度达到80%~90%时取出培养皿,PBS液清洗,加入胰酶,37 ℃、5%CO2培养箱中观察细胞消化状态,见细胞浮立时吸除多余胰酶,加入细胞培养液,离心制成细胞悬液分装至3个培养皿,37 ℃、5%CO2培养箱恒温培养;后行细胞冻存处理,待进行后续实验。
1.2.2 引物设计、siRNA序列合成及转染 引物设计:通过美国国立生物信息中心(National Center for Biotechnology Information,NCBI)数据库检索HOTAIR核酸序列。利用引物设计软件Primer5,根据siRNA设计原则进行HOTAIR及内参β-actin的核酸引物序列设计,由GenePharma公司合成,于-20 ℃保存。核酸引物序列如下(表1)。

表1 HOTAIR及内参β-actin核酸引物序列
siRNA序列合成:由GenePharma公司设计、合成3条 HOTAIR-siRNA干扰序列 和1条阴性对照NC-siRNA序列。实验分组:实验组(转染HOTAIR-siRNA),阴性对照组(转染NC-siRNA)及空白对照组(无siRNA转染即without-siRNA)。HOTAIR-siRNA和NC-siRNA核酸序列如下(表2)。

表2 HOTAIR-siRNA和NC-siRNA核酸序列
细胞转染:选用6孔板于转染前一天接种适量待转染细胞,无抗生素培养基培养,显微镜下观察细胞密度达到60%~70%时进行转染。50 μl无血清Opti-MEM培养基加入20 pmol siRNA混匀;1 μl Lipo2000 转染试剂稀释于50 μl无血清Opti-MEM培养基中室温静置5 min;稀释后的Lipo2000和siRNA试剂室温静置20 min形成复合物,加入含有细胞培养基的培养孔板中,37 ℃、5%CO2培养箱孵育6 h,除去复合物,更换含1%青链霉素和10%FBS完全培养基,37 ℃培养48 h。
1.2.3 qRT-PCR反应 取对数生长期细胞,采用Trizol 法逐步获取细胞总RNA。采用紫外分光光度计于260 nm 和280 nm处检测总RNA吸光度值OD260、OD280,吸光度值介于1.8~2.0间说明RNA质量及纯度良好。按照逆转录试剂盒说明书配置10 μl PCR逆转录反应体系反应得到cDNA,并检测其浓度。利用荧光定量反应试剂盒说明书配置qRT-PCR反应体系及反应条件,将逆转录所得cDNA根据反应条件进行PCR扩增,反应条件:预变性(95 ℃,60 s,35个循环),退火(60 ℃,60 s,35个循环),延伸(70 ℃,5 min),实验重复3次,4 ℃保存。绘制溶解曲线,采用2-ΔΔCt法定量分析各细胞中HOTAIR相对表达量,筛选干扰效率最佳的一组HOTAIR-siRNA序列进行后续实验。
1.2.4 CCK-8法检测细胞增殖能力 于96孔板接种待测细胞悬液,每孔 5×103个,次日分组转染HOTAIR-siRNA及NC-siRNA,每组设3个复孔、5个平行孔,37 ℃、5%CO2培养24 h;采用移液枪向每孔加入CCK-8溶液约10 μl,培育箱继续孵育1 h;于转染后6、12、24、48、72 h采用酶标仪测量各组450 nm处吸光度值。实验重复3次取平均值。
1.2.5 Transwell实验检测细胞侵袭及迁移能力 细胞侵袭:无血清RPMI1640培养基重悬各细胞,饥饿24 h,调整浓度为1×105个/孔,取200 μl细胞悬液接种于Transwell小室上室无血清培养基中;Transwell小室下室加入含20%胎牛血清的RPMI1640培养基,37 ℃恒温孵育48 h;取出Transwell小室,PBS液冲洗,95%乙醇固定,结晶紫染色;显微镜下随机选取5个视野观察拍照计数,实验重复3次取平均值。细胞迁移:试验中调整细胞浓度为5×104个/孔加入Transwell小室上室无血清培养基中,其他方法基本同侵袭检测。
1.2.6 Western blot 检测下调HOTAIR表达后Wnt/β-catenin信号通路相关蛋白表达 使用PMSF蛋白裂解液提取各转染组细胞总蛋白,后采用BCA法于562 nm处检测吸光度值计算样品蛋白浓度。取50 μg蛋白样品,根据Bio-Rad Bis-Tris凝胶系统说明书配制10%SDS-PAGE凝胶体系,于电泳槽内加入电泳缓冲液,将蛋白和蛋白样品预染Marker上样至加样孔进行蛋白电泳,电泳条件浓缩胶(80 V,30 min),分离胶(100 V),待溴酚兰到达凝胶底部时结束电泳。将分离蛋白印迹转至PVDF膜,5%脱脂牛奶中室温封闭1 h,TBST洗膜3次。TBST液稀释抗体,加入一抗4 ℃孵育过夜。TBST洗膜3次,加入PVDF膜与辣根过氧化物酶偶连的二抗室温共孵育1 h,进行抗体标记。采用Bio-Rad ChemiDocTMXRS system凝胶成像系统及Image LabTM软件进行光学信号检测,分析目的条带的相对光密度值。
1.3 统计学处理
2 结果
2.1 不同NSCLC细胞系中LncRNA HOTAIR相对表达情况
为明确NSCLC细胞中LncRNA HOTAIR是否差异表达,采用qRT-PCR法对不同NSCLC细胞系中HOTAIR水平进行检测,并以正常肺组织细胞HBE作为对照。结果显示:与正常HBE细胞对照比较,NSCLC细胞系A549、H1650、H460中HOTAIR均高表达,组间差异具有统计学意义(P<0.01),见图1。
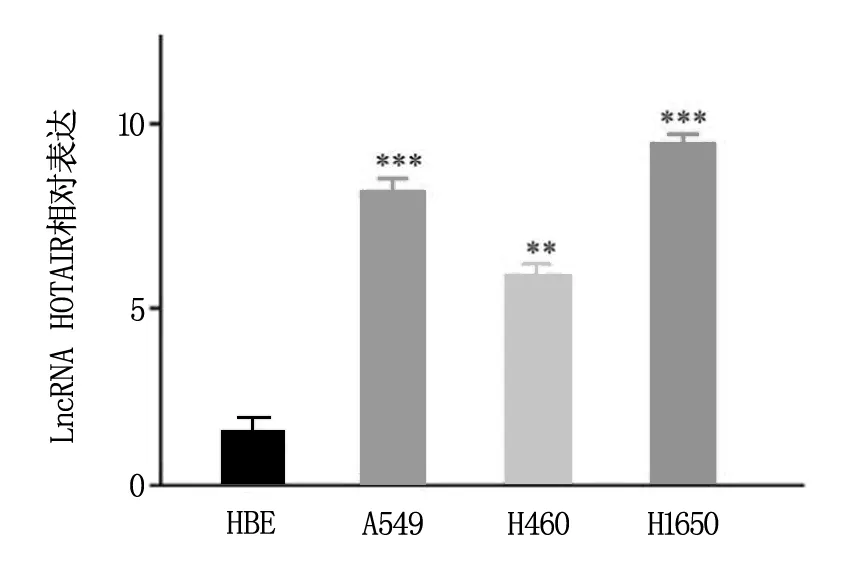
注:与HBE细胞相比,**为P<0.01,***为P<0.001。图1 各NSCLC细胞系中HOTAIR相对表达
2.2 敲降LncRNA HOTAIR的效果
将GenePharma公司设计合成的3条不同HOTAIR-siRNA干扰序列分别转染至A549、H1650、H460细胞系中敲降HOTAIR表达(即实验1、2、3组),以转染NC-siRNA作为阴性对照。经转染72 h后,采用qRT-PCR法对NSCLC各细胞系中HOTAIR表达量进行检测,结果显示:A549、H1650、H460细胞系经转染3条不同HOTAIR-siRNA序列后HOTAIR相对表达均较阴性对照组相比呈现不同程度敲降效果(P<0.01);对比三组细胞系的各组敲降效率,HOTAIR-siRNA3敲降效果均为最佳,选择其进行后续实验。见图2。
2.3 敲降LncRNA HOTAIR基因后各组肺癌细胞系中HOTAIR表达
NSCLC细胞系A549、H1650、H460分别转染HOTAIR-siRNA,敲降HOTAIR基因表达。经转染48 h后取对数生长期各细胞系分别进行qRT-PCR分析,结果显示:敲降HOTAIR基因表达后,3组细胞系中HOTAIR相对表达水平均较阴性对照组及空白对照组明显降低,组间差异具有统计学意义(P<0.05),见图3。
2.4 敲降LncRNA HOTAIR表达对各肺癌细胞增殖能力的影响
采用CCK-8法分别于转染6 h、12 h、24 h、48 h、72 h检测各组细胞OD值并绘制增殖曲线,结果显示:敲降HOTAIR表达,A549、H1650、H460细胞在各转染时间点的增殖能力较阴性对照组和空白对照组相比均受到不同程度抑制,组间差异具有统计学意义(P<0.05),见图4。
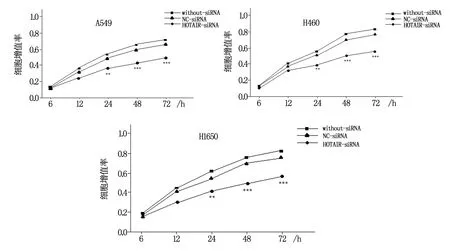
注:与空白对照组和NC-siRNA组比较,**为P<0.01,***为P<0.001。图4 敲降HOTAIR后各细胞增殖能力
2.5 敲降LncRNA HOTAIR表达对各肺癌细胞侵袭/迁移能力的影响
采用Transwell实验检测各组细胞侵袭/迁移能力,结果显示:敲降HOTAIR表达后,A549、H1650、H460细胞的相对侵袭率/迁移率较阴性对照组均明显降低,组间差异具有统计学意义(P<0.01),见图5。
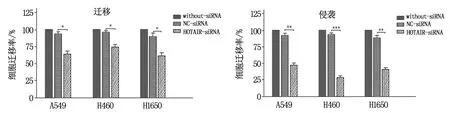
注:与NC-siRNA组比较,*为P<0.05,** 为P<0.01,***为P<0.001。图5 敲降HOTAIR后各细胞迁移/侵袭能力
2.6 HOTAIR对Wnt/β-Catenin信号通路的影响
为明确HOTAIR是否通过诱导Wnt/β-Catenin信号转导通路参与NSCLC的发生,本研究采用Western blot法对Wnt/β-Catenin信号通路的重要蛋白β-catenin、c-Myc水平进行检测,探究了敲降HOTAIR基因表达对各NSCLC细胞系中β-catenin、c-Myc表达水平的影响,并根据蛋白条带强度进行量化分析。结果显示:与阴性对照组及空白对照组相比,敲降HOTAIR基因表达后,A549、H1650、H460细胞系的β-catenin、c-Myc表达水平均明显减弱(P<0.05)。敲降细胞内HOTAIR表达可抑制β-catenin、c-Myc表达,HOTAIR表达水平能够调控Wnt/β-Catenin信号通路的活性。见图6。
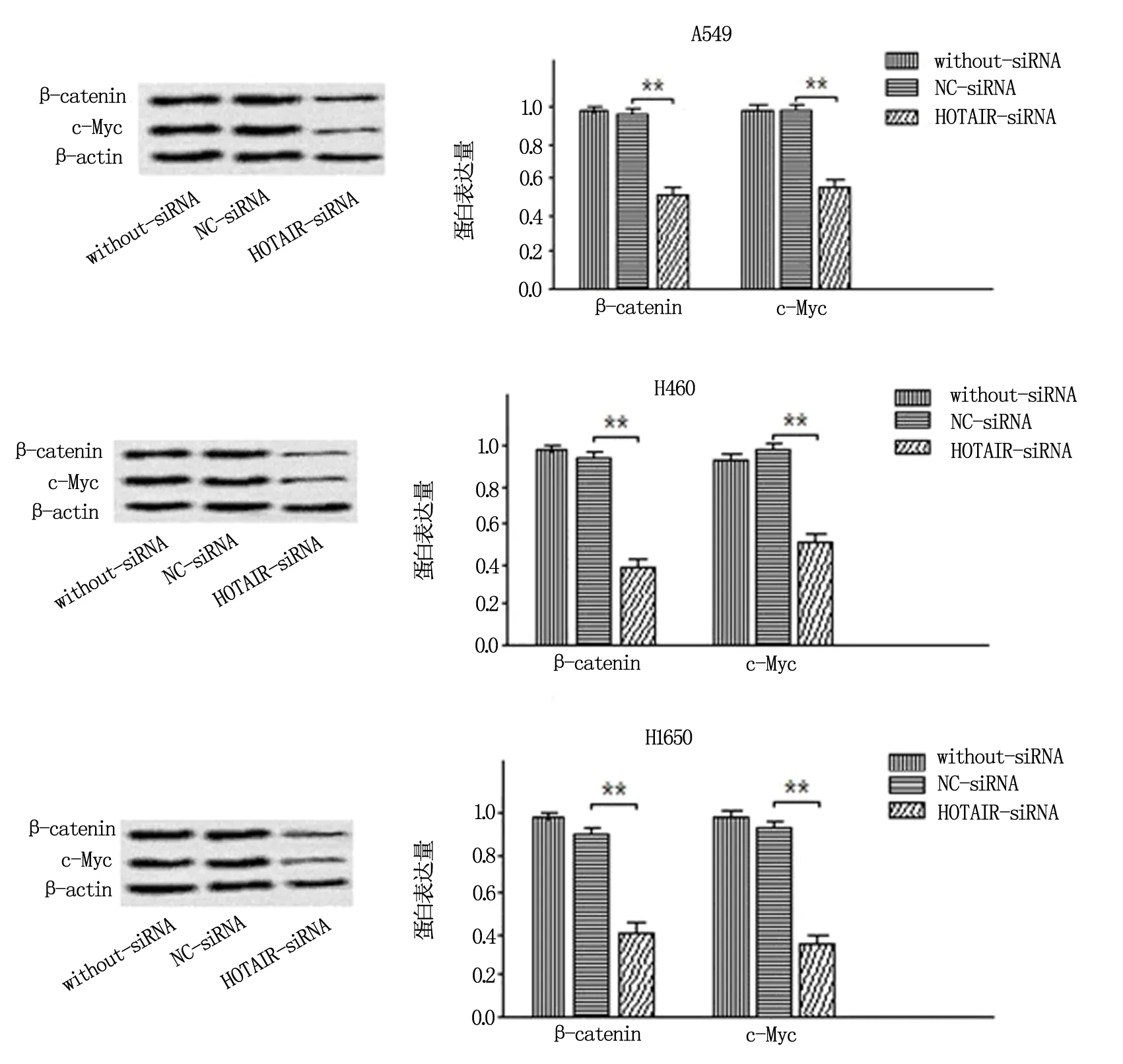
注:与NC-siRNA组比较,** 为P<0.01。图6 敲低HOTAIR表达对Wnt/β-Catenin信号通路中相关蛋白的影响
3 讨论
肺癌作为最常见的恶性肿瘤,其发病是多因素共同导致的结果,包括原癌基因、癌基因突变、LncRNA表达失调及DNA甲基化状态改变等。近年来,随着基因测序技术的发展及人类基因库数据的不断完善,基因测序显示人类基因组中仅有<3%的基因可编码为蛋白,其余均转录为非编码RNA(ncRNA )。研究指出[7],ncRNA中长链非编码RNA(LncRNA)积极参与各细胞中信号转导通路的传递,是调节多种细胞进程的重要因子。据有关报道[8],LncRNA的异常表达或失调会导致肿瘤的发生发展,在肿瘤中发挥促癌或抑癌作用,可在转录水平、转录后水平及表观遗传学水平等多方面对肿瘤进行调控[9],为肿瘤的预防及治疗提供了新的研究方向及治疗靶点。
HOTAIR是HOX的转录反义RNA,是第一个被识别具有反式转录调控作用,且与恶性肿瘤相关的 lncRNA,位于基因组 12q13上HOXC 基因位点[10]。研究表明[11],HOTAIR主要与多梳蛋白抑制复合物2(polycomb repressive complex 2,PRC2)相互作用,调节染色体重排,进而促进肿瘤的发生。Li等发现[12],HOTAIR在肝癌组织中高表达,通过抑制抑癌基因表达促进肝癌发生;反之下调HOTAIR表达则可抑制肝癌细胞的生长迁移。Pan等发现[13],胃癌组织中HOTAIR过表达,对胃癌淋巴结转移、EMT进展及血管侵犯具有促进作用,HOTAIR过表达是造成胃癌患者预后不良的主要标志物。Zhang等研究报道[14],人神经胶质瘤中,HOTAIR可通过互补序列下调miR-126,影响神经胶质瘤的进展。此外,除发挥促癌/抑癌作用外,HOTAIR也参与肿瘤治疗后顺铂耐药;沉默HOTAIR表达可增加恶性肿瘤细胞对化疗药物的敏感性,改善患者预后,并指出HOTAIR具有预测肿瘤化疗耐药的特质[15]。因此本研究就LncRNA HOTAIR对NSCLC细胞生物学行为及其可能作用机制进行了探究,结果发现NSCLC细胞系中 HOTAIR异常高表达;转染敲降HOTAIR表达后NSCLC细胞增殖、迁移及侵袭能力均受到显著抑制。提示HOTAIR 具有类癌基因的作用,可促进癌细胞的增殖、迁移及侵袭。
基因参与调控肿瘤的发生发展通过细胞信号通路进行信息传导,进而影响细胞生物学功能,而LncRNA主要通过:①作为信号传导分子参与信号通路传导,调控下游基因转录;②直接与蛋白结合阻断分子作用和信号通路,调控下游基因;③与蛋白结合,将复合物定位至特定DNA序列;④直接或间接结合多个相关转录因子,实现多种信号通路间信息整合等机制调节细胞功能[16]。研究发现[17],Wnt信号传导通路是肺癌细胞中唯一保持高活性的信号通路,肿瘤的发生发展与Wnt通路密切相关;经典Wnt/β-Catenin 信号通路异常激活,诱导转录因子的表达,是导致肿瘤发生的主要驱动力。此外,Wnt信号通路是1个复杂的调控空网络,其关键蛋白发生突变,导致信号异常活化,能够诱导肿瘤的发生[18]。Ding等[19]研究发现,肺癌组织中Wnt1蛋白含量上调,Wnt1高表达与异常高表达的c-Myc、β-Catenin等癌基因相关,Wnt1高表达肺癌细胞呈现较强的增殖、迁移、侵袭能力和抗肿瘤细胞凋亡。Wang等[20]发现,抑制癌细胞中β-Catenin转录活性,阻碍Wnt/β-Catenin通路,进而可抑制c-Myc等癌基因表达及细胞增殖。同时Fu等也研究发现[21],HOTAIR与PRC2联合调控H3K27三甲基化,改变下游PTEN、WIF1等基因表达,对Wnt、AKt等信号通路发挥作用,参与调控肿瘤的生长、凋亡等过程。因此深入探究Wnt/β-Catenin信号通路与LncRNA HOTAIR间的潜在调控关系及机制,可提供新的肺癌治疗靶点。本研究探究发现敲降HOTAIR表达后,各NSCLC细胞系中Wnt通路相关蛋白β-catenin、c-Myc表达水平明显减弱,提示敲降HOTAIR表达可抑制Wnt/β-Catenin信号通路相关蛋白的表达,抑制Wnt/β-Catenin信号通路的活性,进而抑制NSCLC的发生。
综上,LncRNA HOTAIR在NSCLC细胞系中高表达,敲降细胞内HOTAIR表达可抑制NSCLC细胞增殖、迁移及侵袭,其机制可能与Wnt/β-Catenin信号通路活性受抑制有关。
