入侵植物剑叶金鸡菊种群遗传多样性及克隆多样性研究
2021-01-28曾建军王宁
曾建军,王宁
井冈山大学生命科学学院,江西 吉安 343009
入侵植物因对环境、生物多样性等造成危害而受到广泛关注,入侵机制是入侵生态学一个非常重要的研究内容,其中对繁殖机制的研究有助于揭示外来植物入侵和扩张机理(Burrell et al.,2015;Van et al.,2015)。许多入侵植物同时进行有性繁殖和无性繁殖,其中克隆生长是最主要的无性繁殖方式(Douhovnikoff et al.,2003)。克隆生长不可避免会增加传粉者在同一克隆内访花频率,从而导致雌性适合度(如种子产量)下降(田昊等,2018),尤其是克隆构型为密集型的植物,其交配系统存在异交向自交进化的选择压力(Barrett et al.,2008)。但是,研究表明自然界中仍有许多密集型克隆生长的植物仍保持专性异交特性,克隆植物如何规避克隆生长带来的同株异花授粉的影响,从而为异交提供更多的机会?该问题引起越来越多的生态学家和进化生物学家的关注(Barrett,2015)。克隆生长通过影响克隆大小、种群克隆结构等方面影响花粉传播模式,进而影响个体的有性繁殖成功(田昊等,2018)。对种群遗传多样性和克隆多样性,将有助于了解种群更替动态及繁殖策略(Torimaruet al.,2005;Mandel et al.,2019)。入侵植物在被引入到新的栖息地之后,必须克服一系列的障碍,最重要的一种障碍即为繁殖,其中必定蕴涵着复杂多样的适应与进化信息,对入侵克隆植物繁殖机制的研究,为揭示克隆生长对有性繁殖的影响提供了很好的研究视角(Garcia et al.,2002;Bai et al.,2013)。
剑叶金鸡菊(Coreopsis lanceolata)原产于北美,为菊科金鸡菊属多年生草本克隆植物,1936年作为园林植物引入我国江西庐山(万慧霖等,2008;许媛等,2009)。剑叶金鸡菊具有密集和游击型克隆生长特性,虽然交配系统为专性异交,但克隆生长并未影响到有性繁殖成功,单株种子产量大(曾建军等,2010)。正因为适应性、繁殖能力强,剑叶金鸡菊逃逸为有害野生杂草,被列为我国主要外来入侵物种之一(徐海根等,2004)。江西庐山属海拔直落差大的山地环境,是研究物种入侵与生态环境的一个天然的实验平台(万慧霖等,2008),本研究以江西庐山剑叶金鸡菊不同海拔(低和高海拔)、不同人为干扰程度(干扰大的公路旁绿化带和干扰小的荒地)种群为实验对象,通过AFLP分子标记方法分析种群遗传多样性、克隆多样性和克隆结构,调查不同种群种子结实率,探讨剑叶金鸡菊克隆生长对有性繁殖的影响,以期为揭示剑叶金鸡菊入侵机制提供理论依据。
1 材料与方法
1.1 研究地概况
样地设置于江西庐山,地理坐标在 115°50′—116°10′E,29°28′—29°45′N,总面积为 30500 hm2,具典型中亚热带季风侯特征和山地气候特色,年平均气温16.7 ℃,年平均降水量约1300 mm(杨道德等,2007)。
1.2 试验材料与分析方法
1.2.1 供试材料
选择江西庐山高海拔公路旁绿化带、高海拔荒地和低海拔荒地共3个剑叶金鸡菊种群,各种群的基本情况见表1。将各种群按照50 cm×50 cm划分的连续正方形,记录空间坐标(图1)对每个点上的个体进行定位取样,每株取健康幼嫩叶片,放入装有硅胶的密封袋中干燥保存备用(Li et al.,2009;Jiao et al.,2020)。依照 Douhovnikoff et al.(2003)的方法标记7株不同基因型个体,每基株分别取3株克隆分株的叶片,共21个样,同样的方法干燥保存作为对照组,用于计算 Jaccard’s相似系数阈值(T)。
1.2.2 试验方法
1.2.2.1 DNA提取
采用的Wilson et al.(2005)方法提取总DNA。
1.2.2.2 AFLP分析
反应总体系为20 μL,包括4 μL模板DNA(50 ng·μL-1),2 μL Hind III/Mse I,1 μL Adapter,2.5 μL 10 mM ATP,2.5 μL 10X Reaction buffer,7 μL AFLP-Water,1 μL T4 Ligase,混匀离心数秒,37 ℃保温5 h,8 ℃保温4 h,4 ℃过夜。预扩增反应体系 25 μL,含 2 μL 模板 DNA,1 μL dNTPs,1 μL Pre-ampmix,2.5 μL 10X PCR buffer,0.5 μL Taq DNA polymease,18 μL水。离心数秒,PCR扩增循环30轮,扩增参数:变性94 ℃ 30 s,56 ℃ 30 s下复性,延伸72 ℃条件下80 s,72 ℃ 5 min。按1∶20稀释预扩增产物,作为选扩模板。选择性扩增体系反应体系25 μL,含预扩增稀释样品2 μL,0.5 μL dNTPS,2.5 μL 10XPCR buffer,1 μL(共 8 种)Mse I引物,1 μL(共 8 种)Hind III引物,0.5 μL Taq酶,17.5 μL H2O。扩增程序如下:第一轮扩增参数:94 ℃,30 s;65 ℃,30 s;72 ℃,80 s。以后每轮循环温度递减0.7 ℃,扩增12轮,然后按94 ℃,30 s;55 ℃,30 s;72 ℃,80 s参数扩增23轮。通过ABI 377测序仪进行AFLP多态性分析。从100个引物中筛选出清晰、不弥散、不模糊且重复性好的条带,阴性对照中无带的引物作为正式扩增引物。扩增使用8个引物序列:Eco-AAC/Mse-CAA;Eco-AAG/Mse-CAC;Eco-ACA/Mse-CAG;Eco-ACT/Mse-CAT;Eco-ACC/Mse-CTA;Eco-ACG/Mse-CTC;Eco-AGC/Mse-CTG;Eco-AGG/Mse-CTT。
2 数据统计与分析
遗传多样性参数计算方法:对照反应产物在凝胶上的对应位置,统计电泳图谱中70—500 bp的条带,“1”为有带,“0”为无带,生成0/1矩阵。采用POPGEN 32软件计算多态位点百分率、有效等位基因数(Effective numbers of alleles,Ne)、等位基因数(Observed number of alleles,Na)、Nei’s遗传多样性指数(Nei’s gene diversity,H)和 Shannon’s信息指数(Shannon’s information index,I)用来估算基因多样性(宋振等,2019)。
采用Douhovnikoff et al.(2003)方法计算两两样本是否来自同一克隆(基株)。计算同一种群两两样本(x和y)间Jaccard’s相似系数Sxy,Sxy=n11/(n11+n10+n01),n11是个体x和y都有的条带数,n10是只在x个体上特异性条带数量,n01是只在y个体上的特异性条带数量。相似系数的计算用SPAGeDi 1.4软件。当两植株间相似系数Sxy大于Jaccard’s相似系数阈值T值时,认为这两株植物属于同一克隆(基株)(Wilson et al.,2005)。
采用以下3个参数估算克隆多样性:(1)基因型比例(G/N)=G/N(Dorken et al.,2001;Liet al.,2009);(2)Simpson多样性指数(D);(3)基因型分布均匀度(E)(Pielou,1969;李钧敏等,2009)。

表1 剑叶金鸡菊的不同生境概况Table 1 The conditions of the different habitants of C. lanceolata

图1 种群基株分布图Fig. 1 Distribution of genets in different population
结实率计算方法:选取样地标记 10个植株,每株选取果实5个,计算每株植株结实率以及每个种群的平均结实率。
3 结果与分析
3.1 剑叶金鸡菊不同种群遗传多样性分析
对剑叶金鸡菊种群遗传多样性分析表明(表2),3个种群多态性位点百分率(P)均很高,最低为庐山植物园种群(LBG)89.74%,最高石门涧种群(SMJ)可达 95.13%。Shannon’s指数最大的是石门涧种群(SMJ)为0.51,指数排序为石门涧种群 (SMJ)>牯岭种群 (GL)>庐山植物园种群(LBG)。以Nei’s多样性指数(H)为指标3个种群的排序与Shannon’s指数排序结果一致。
3.2 剑叶金鸡菊不同种群克隆多样性分析
为了辨别种群内不同株是否来自同一克隆,通过对21个样(7个不同基因型基株,每株分别取3个克隆分株)AFLP分析,计算出Jaccard’s相似系数阈值为0.88,即两两植株相似系数大于该阈值即认为来自同一克隆。由表3可知,牯岭(GL)种群的基因型比例和 Simpson多样性指数最低(G/N=0.65,D=0.91),石门涧(SMJ)种群最高(G/N=0.88,D=0.98)。基因型均匀度(E)最高为庐山植物园(LBG)种群0.87,牯岭(GL)和石门涧(SMJ)种群分别为0.53和0.57。3个种群克隆分株间平均距离1.2 m,范围0.93—1.65 m,克隆分株最远距离为1.80 m。
3.3 不同种群剑叶金鸡菊结实率分析
图2结果表明,无论是高海拔还是低海拔种群,或者人工扰动大的公路旁及干扰低的荒地种群,剑叶金鸡菊均有较高结实率,每个种群结实率平均数在64.97%—70.93%之间,不同种群之间的结实率差异不显著。
4 讨论
本研究AFLP分子标记分析结果表明,不同生境剑叶金鸡菊天然种群均有较高遗传多样性。曾建军等(2010)分析了剑叶金鸡菊结实特征,其单株种子产量达到12000粒,种子千粒质量 (1794±18)mg,种子萌发率在60%以上,落地可繁殖。我们推测有性繁殖能力强是剑叶金鸡菊种群遗传多样性高的重要原因。由于天然屏障作用,入侵植物从低海拔向高海拔扩散能力逐渐减低,植物的遗传多样性可能随海拔升高而降低(Alpertet al.,2000;Pauchardet al.,2009)。但是Bustamante et al.(2017)研究发现入侵植物花菱草(Eschscholzia californica)从低海拔到高海拔遗传多样性并未减低的现象。通过分子标记方法显示,人类多次引入是入侵克隆植物Furcraea foetida(L.)引入地种群和入侵地种群均可保持高遗传多样性的重要因素(Barbosa et al.,2019)。剑叶金鸡菊属于多次引入园林植物(万慧霖等,2008),因此我们推断,多次引入是剑叶金鸡菊种群保持较高遗传多样性水平的原因之一。种群遗传多样性高有利于植物有性繁殖成功(Reed et al.,2003;Kettenring et al.,2019),剑叶金鸡菊种群较高的遗传多样性同时也提高了有性繁殖适合度,促进有性繁殖成功,提高入侵能力。
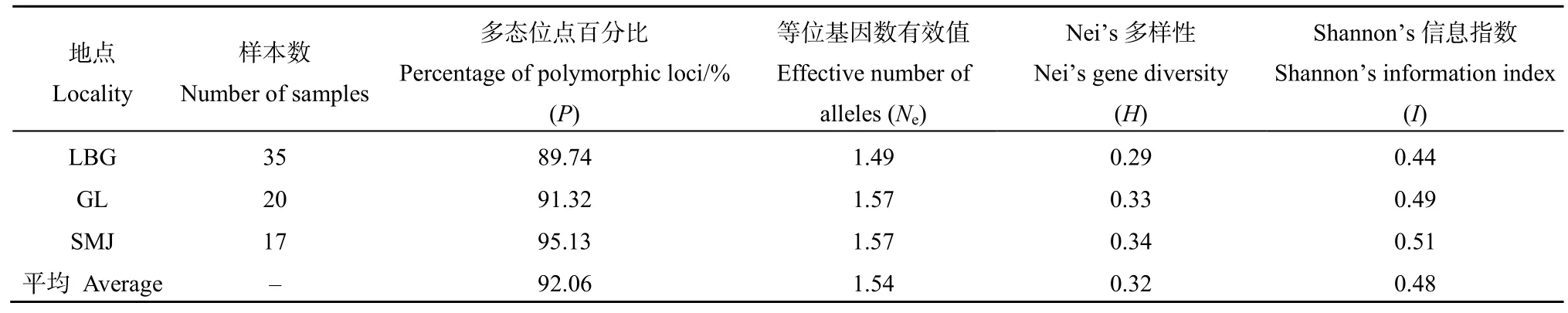
表2 剑叶金鸡菊不同种群遗传多样性分析Table 2 Statistical analysis of genetic variation of C. lanceolata in different population

表3 剑叶金鸡菊不同种群克隆多样性分析Table 3 Statistical analysis of Clonal diversity of C. lanceolata in different population

图2 剑叶金鸡菊不同种群结实率Fig. 2 Setting rate of C. lanceolata in different population
克隆植物种群的基因型比例G/N的平均值约为0.44(Honnayet al.,2008),本研究中剑叶金鸡菊3个种群基因型G/N平均值为0.77,较高G/N值表明剑叶金鸡菊种群内植株体大多来自不同基株,说明游击型生长对提高剑叶金鸡菊种群不同基因型混合度及拓殖种群的作用有限。有性繁殖是克隆植物种群保持较高基因型比例的机制(Bona et al.,2019;Hedrén et al.,2019),剑叶金鸡菊种群较高的克隆多样性进一步表明有性繁殖是该物种种群更替非常重要的繁殖策略。Albert et al.(2008)认为,种群克隆多样性水平与异交率正相关关系,例如欧洲越桔(Vaccinium myrtillus)克隆多样性高的种群异交率也更高。本研究中庐山3个剑叶金鸡菊自然种群果实平均结实率均达64.97%以上,与人工模拟基因型均匀混合种群平均结实率64%基本一致(曾建军等,2014),进一步说明剑叶金鸡菊的克隆生长并未影响异株异花传粉,推测这与其种群克隆多样性高密切相关。
花粉扩散距离与克隆大小之间的关系也影响异交率。Sinclair et al.(2014)研究了克隆植物波喜荡(Posidonia australis)的传粉过程,通过水为媒介,花粉扩散距离可达178 m,远远大于其克隆平均大小30.8 m,因此保证了充足的异株异花花粉数量。在本研究中,剑叶金鸡菊种群内克隆分株间平均距离较小为1.2 m,最远距离仅1.8 m。曾建军等(2014)研究了剑叶金鸡菊主要传粉昆虫的一次最长飞行距离虽然只有60 cm,但传粉昆虫在访花过程中有多个飞行回合,访问的基株数最大达到6株,总飞行最远距离超过 1.8 m。而且我们也观察到与其它伴生植物比较,剑叶金鸡菊的克隆生长产生了更大的花展示,吸引了数量众多的传粉昆虫,提高了异株花粉资源输出,因此保障了异交交配系统的繁殖成功。
5 结论
江西庐山剑叶金鸡菊不同海拔(低和高海拔)、不同干扰程度(干扰大的公路旁绿化带和干扰小的荒地)种群均有较高遗传多样性,有性繁殖能力强和多次引入是该克隆植物保持种群高遗传多样性的重要原因。种群较高的遗传多样性同时也提高了剑叶金鸡菊有性繁殖适合度,促进有性繁殖成功。种群克隆多样性高减低了剑叶金鸡菊同株异花授粉几率,推测剑叶金鸡菊的克隆生长产生了更大的花展示,通过吸引数量众多的传粉昆虫,提高异株花粉资源输出,因此保障了剑叶金鸡菊专性异交交配系统有性繁殖成功。
