水稻籽粒低镉蛋白LCD互作蛋白的筛选与鉴定
2020-12-04郝小花戴佳利暨文劲黄丹李东屏田连福
郝小花 戴佳利 暨文劲 黄丹 李东屏 田连福
(1.湖南文理学院生命与环境科学学院,常德 415000;2.湖南师范大学作物生命科学学院 作物不育资源创新与利用湖南省重点实验室,长沙 410081)
由于工业化的发展和化肥的施用,耕地普遍存在镉(Cd)污染问题。水稻属于易吸收和积累Cd的作物,在Cd污染稻田中种植水稻会导致大米Cd超标的风险。全世界约有一半的人口以稻米为主食,长期食用Cd超标的大米,对人体健康产生不利影响。培育籽粒Cd低积累的水稻品种是防止大米Cd超标的有效办法之一。
近年来,随着分子生物学、测序技术和生物信息学技术的发展,有关植物在重金属解毒和重金属耐受方面的研究取得较大进展。植物进化出各种机制来维持必需金属离子的生理浓度[1-3],同时最大限度地减少植物体内过多重金属离子对植物细胞造成的损害,从而赋予植株对重金属胁迫的耐受性。当植物暴露于达到毒害作用的重金属浓度时,首先通过限制重金属离子进入外质体、将它们与细胞壁或细胞渗出物结合、降低根细胞对重金属离子的吸收,或通过抑制长距离运输把重金属阻隔在地下组织中[4-6]。其次,已进入细胞的重金属离子,可以通过一系列的存储和解毒策略缓解,包括离子运输、螯合或隔离到液泡中[7]。最后,植物还可以通过激活氧化应激防御机制和与应激相关的蛋白质和信号分子等激活不同的防御或抵抗途径来进行防御,如热休克蛋白、激素和活性氧等[8-9]。水稻中已经鉴定出一些与Cd转运、解毒等相关的基因,包括OsNRAMP1[10]、OsNRAMP2[11]、OsNRAMP3[12]、OsNRAMP5[13]、OsNRAMP6[14]、OsHMA2[15]、OsHMA3[16]、OsHMA9[17]、OsIRT1[18]、OsIRT2[19]、OsZIP1[20]、LCD[21]、OsLCT1[22]、OsPCR1[23]、O s C C X 2[24]、O s A B C G 4 3[25]、O s C A L 1[26]、OsPCS1[27]、OsPCS2[28]、OsPCS15[29]、OsMTP1[30]等,它们在水稻中,参与Cd的解毒、吸收、运输和分配。
水稻LCD是一个没有同源基因的单基因。该基因突变导致水稻籽粒中Cd含量大幅降低[21]。lcd突变体在植物早期发育过程中,无论是在平板中还是在水培液中都表现出对Cd的耐受性。Cd含量测定表明,突变体植株的地上部分的Cd含量比野生型显著降低。在Cd污染的土壤中进行田间培养,突变体叶片的Cd含量与野生型差异不显著,但籽粒中Cd含量显著降低。lcd突变体与野生型植株的生物量和种子产量无显著差异。LCD定位于细胞质和细胞核中,主要在根系的维管组织和叶片的韧皮部伴胞细胞中表达[31],可能参与调控Cd的长距离运输。敲除该基因可以限制Cd向地上部,尤其是籽粒的运输,从而降低籽粒中Cd的含量。蛋白结构分析显示,OsLCD蛋白没有富含半胱氨酸,不具有重金属解毒功能,同时也不包含跨膜结构域,没有转运功能,推测LCD蛋白可能通过与其他蛋白相互作用来调节水稻体内Cd的积累和运输。
酵母双杂交(Yeast two hybrid)是一种有效的筛选和鉴定蛋白相互作用的分子生物技术[31-33],在解析蛋白作用的分子机制研究中有广泛的应用。传统的酵母双杂交系统仅限于核蛋白的互作分析,不能进行膜蛋白互作的研究。Stagljar等[34]发展了基于分裂泛素(Split-ubiquitin)的酵母双杂交系统,克服了传统酵母双杂交系统的局限性。DUALmembrane系统是基于分裂泛素介导的膜蛋白酵母双杂交系统,可以进行膜蛋白-膜蛋白、膜蛋白-可溶性蛋白间的互作研究。DUALmembrane膜体系酵母双杂交文库系统已应用在水稻条纹叶枯病毒的传播机制的研究和拟南芥纤维素合成相关蛋白互作的研究中[35-36]。
水稻LCD影响籽粒Cd累积,但其作用的分子机制尚不清晰。本研究拟通过酵母双杂交技术筛选和鉴定与LCD互作的蛋白(调控蛋白或转运蛋白),为阐明LCD参与的Cd离子转运、积累的分子机制奠定基础,为挖掘遗传新资源、培育低镉水稻新品种提供理论指导。
1 材料与方法
1.1 材料
水稻品种为粳稻日本晴(Oryza sativa ssp.japonica cv.Nipponbare);引物、培养基、常规化学试剂等订购于上海生工生物工程有限公司,RNA提取试剂Trizol、反转录酶、X-α-gal、AbA、高保真酶、限制性内切酶等购自Invitrogen公司。
膜体系酵母双杂交系统为Biotech公司DUALmembrane system,酵母菌株为NMY51,文库载体为pPR3-N,诱饵载体为pBT3-N,阳性对照载体为pOst-Nub1。
核体系酵母双杂交系统为Clontech公司Yeast Two-Hybrid System,酵母菌株为AH109,文库载体为pADKT7,诱饵载体为pGBKT7,阳性对照载体为pGBKT7-53和pGADT7-T,阴性对照载体为pGBKT7-Lam。
以日本晴生长一周的根、地上部分,分蘖期的根、叶,开花期的根、叶、节、节间、花均量混匀为材料,分别构建DUALmembrane系统和Yeast Two-Hybrid系统cDNA文库,文库构建由上海欧易生物医学科技有限公司完成。
质粒扩繁在大肠杆菌Top10中进行。
1.2 方法
1.2.1 水稻LCD诱饵载体的构建 根据水稻LCD序列设计特异性引物,以cDNA文库为模板,扩增LCD编码区序列,经纯化后,与T载体pMD18-T连接,转化,并提取阳性菌落的质粒。依据LCD的序列和诱饵载体(pBT3-N、pGBKT7)的多克隆位点,选择合适的酶切位点设计构建诱饵载体的引物(表1)。对pMD18-T-LCD和诱饵载体进行酶切、纯化,用T4连接酶连接过夜,转化,筛选阳性克隆,提取阳性质粒进行酶切、测序验证获得正确的诱饵载体。
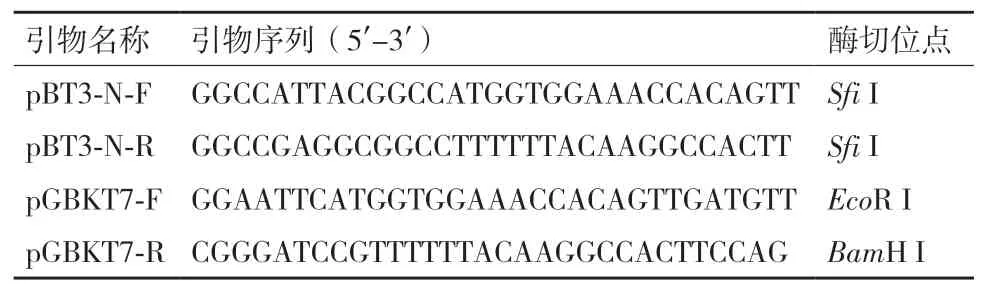
表1 构建诱饵载体的引物序列
1.2.2 重组载体转化酵母细胞 将酵母菌种(NMY51、AH109)在YPDA平板上划线,30℃倒置培养3 d左右,挑取单克隆酵母于50 mLYPDA培养基中,30℃,230-250 r/min振荡培养过夜;测定OD546=0.6-0.8;2 500 r/min离心5 min收集菌体;每个反应管加1.5 μg诱饵质粒,300 μL PEG/LiOAc master Mix(50%PEG 2.4 mL、1 mol/L LiOAc 360 μL和Single-stranded carrier DNA 250 μL),轻轻涡旋混匀,100 μL重悬细胞,涡旋1 min至各组分充分混匀;42℃水浴孵育45 min;500×g 离心5 min,弃上清,用150 μL NaCl(0.9%)溶液重悬菌体;涂不同的营养缺陷平板,30℃倒置培养3-5 d。
1.2.3 文库载体插入片段的扩增和测序 应用文库载体的通用引物(表2),对筛选出的阳性酵母中水稻cDNA文库载体(pPR3-N、pGADT7)的插入片段进行菌落PCR扩增。PCR扩增体系为10×buffer 2 μL、dNTP各0.2 mmol/L、上下游引物各1 μL(10 μmol/L)、Taq 0.5 μL,补充ddH2O至20 μL。PCR产物纯化后送到上海生工生物公司测序。得到的序列在NCBI中用BLAST程序比对查找同源序列并确认基因。
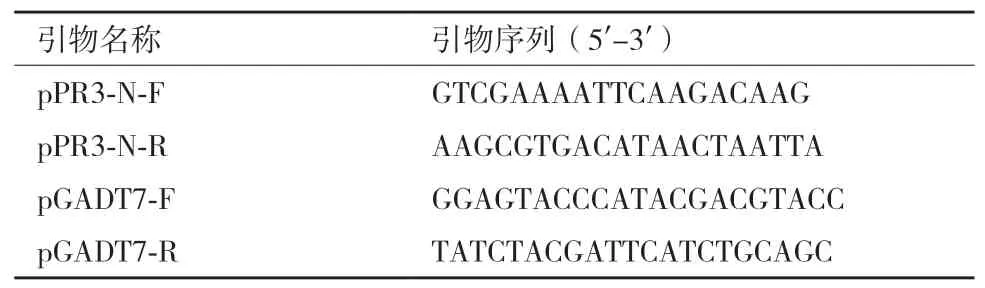
表2 扩增文库载体插入片段的引物序列
2 结果
2.1 DUALmembrane酵母双杂交系统(膜系统)筛选与LCD互作的蛋白
2.1.1 诱饵蛋白LCD自激活及膜系统酵母筛库体系检测 根据膜系统酵母双杂交的载体选择原则,选择pBT3-N作为诱饵蛋白表达载体进行筛库,诱饵蛋白载体为pBT3-N-LCD。按照表3载体组合检测诱饵蛋白载体pBT3-N-LCD的自激活。结果(图1)表明,转化1的酵母菌可以在SD-Leu平板上正常生长,说明pBT3-N-LCD表达的融合蛋白对NMY51酵母细胞的生长无毒性;转化组合2在SD-Lue-Trp、SD-Lue-Trp-His和SD-Lue-Trp-His-Ade平板上均可以生长,说明pBT3-N-LCD和pOst-Nub1表达的融合蛋白可以相互结合并激活下游报告基因的表达,表明筛库系统可以正常工作;转化组合3在SD-Lue-Trp平板上可以正常生长,在SD-Lue-Trp-His平板上少量生长,在SD-Lue-Trp-His-Ade平板上完全不能生长(图1),说明pBT3-N-LCD和pPR3-N表达的融合蛋白不存在自激活作用。根据酵母生长结果,选择SD-Lue-Trp-His-Ade平板作为后续筛选互作蛋白的筛选条件。
2.1.2 膜系统条件下LCD互作的阳性酵母克隆筛选 将水稻膜系统文库质粒和pBT3-N-LCD质粒共转化酵母NMY51,共标记得到354个阳性克隆。将354个菌落划线至SD-Lue-Trp-His-Ade+X-α-gal+AbA平板上,发现共有284个菌落呈蓝色,说明pBT3-N-LCD与284个蛋白可能存在互作,并激活下游报告基因的表达,不能生长和不能变蓝的菌落视为假阳性,予以舍弃。将第二轮筛选出来的生长良好并呈现蓝色的酵母克隆,再次在SD-Lue-Trp-His-Ade+X-α-gal+AbA平板上筛选(图2),仍然生长良好并呈现蓝色的酵母菌落有228个。通过二、三轮的划线筛选,最终筛选出228个潜在的阳性菌落。
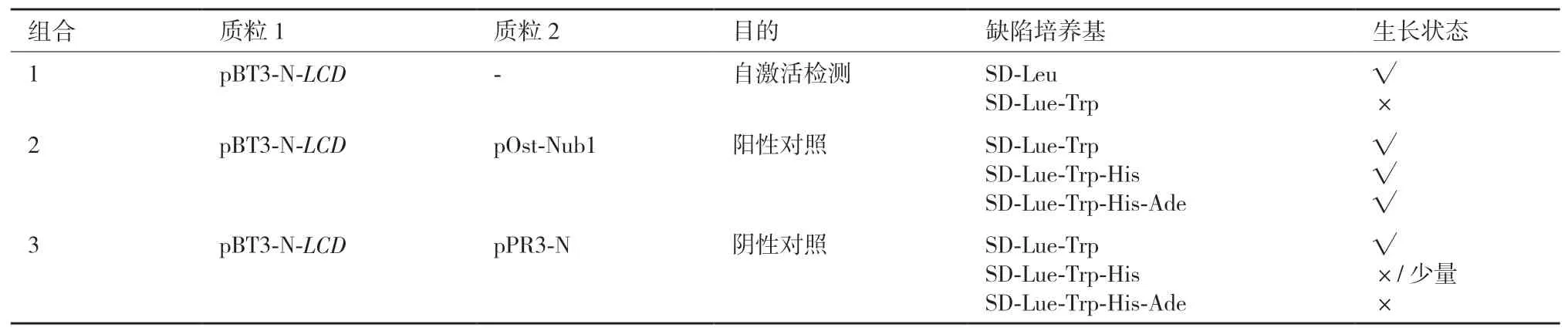
表3 DUALmembrane系统诱饵蛋白LCD自激活体系
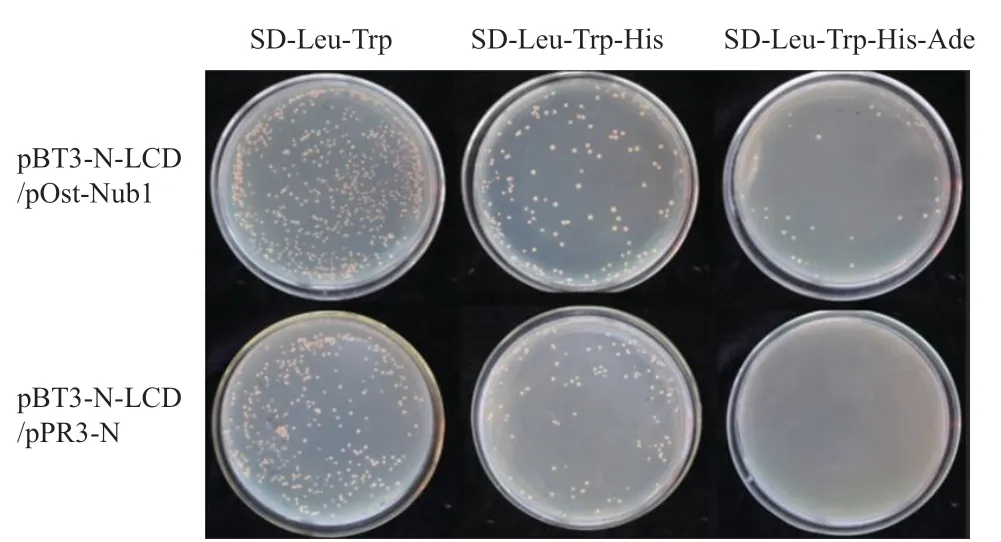
图1 诱饵蛋白LCD在酵母NMY51中的自激活检测

图2 LCD互作的阳性酵母显色筛选
2.1.3 膜系统条件下LCD互作的阳性酵母克隆插入片段的获得与鉴定通过对228个阳性克隆菌落(依次编号为1-228)扩增,得到198个文库质粒pPR3-N中的插入cDNA片段。将198个PCR片段进行测序,去除重复序列,共得到120个测序结果。将测序结果BLAST(http://www.ncbi.nlm.nih.gow/BLAST/)进行基因相似性比较,查找同源基因。进一步去除同源基因中读码框与pPR3-N插入片段读码框不一致的基因,最终共得到22个UniGene(表4)。通过功能预测分析发现,2、11、12、13、19、20与植物耐逆和物质转运相关;1、3、4、7与植物发育响应有关;14、15、16、17、18与细胞的生理代谢有关等。其中11号OsMTP1已有报道,与水稻Cd的积累和耐受性有关。
2.1.4 LCD与膜系统筛选出的22个候选蛋白的互作验证将筛选到的22个基因克隆,构建重组载体pPR3-N-1-22,并按照表5的转化组合进行自激活检测发现,22个基因中编号15(LOC_Os01g38970)不仅可以在SD-Lue-Trp平板上生长,而且可以在SD-Lue-Trp-His-Ade平板上生长,说明LOC_Os01g38970在酵母中存在自激活现象,考虑其互作可能是假阳性,予以舍弃。选择其余的21个基因进行互作验证。
将pBT3-N-LCD分别与pPR3-N-1-22(15除外)共同转化酵母NMY51,进行一对一互作初步验证。结果发现,16不能在SD-Leu-Trp-His-Ade上生长,其余20个均可在SD-Leu-Trp-His-Ade平板上长出大量白色酵母菌落,说明这20个蛋白在酵母中与LCD存在互作,16舍弃。进一步将20个在SDLeu-Trp-His-Ade平板上长出的白色酵母菌落转移至SD-Leu-Trp-His-Ade+X-α-gal+AbA平板上(图3),发现共16个酵母菌斑可以很好的生长并呈现蓝色,说明这16个蛋白与诱饵蛋白LCD之间存在互作,8、17、18、22不能生长或变蓝予以舍弃。最终得到与LCD的互作蛋白为LOC_Os01g67250、LOC_Os10g34614、LOC_Os07g39900、LOC_Os08g01780、LOC_Os02g35560、LOC_Os06g45120、LOC_Os03g38020、LOC_Os08g42050、LOC_Os12g32380、LOC_Os05g03780、LOC_Os07g44180、LOC_Os09g30418、LOC_Os01g69220、LOC_Os06g08170、LOC_Os11g25980和LOC_Os08g44280,共16个。
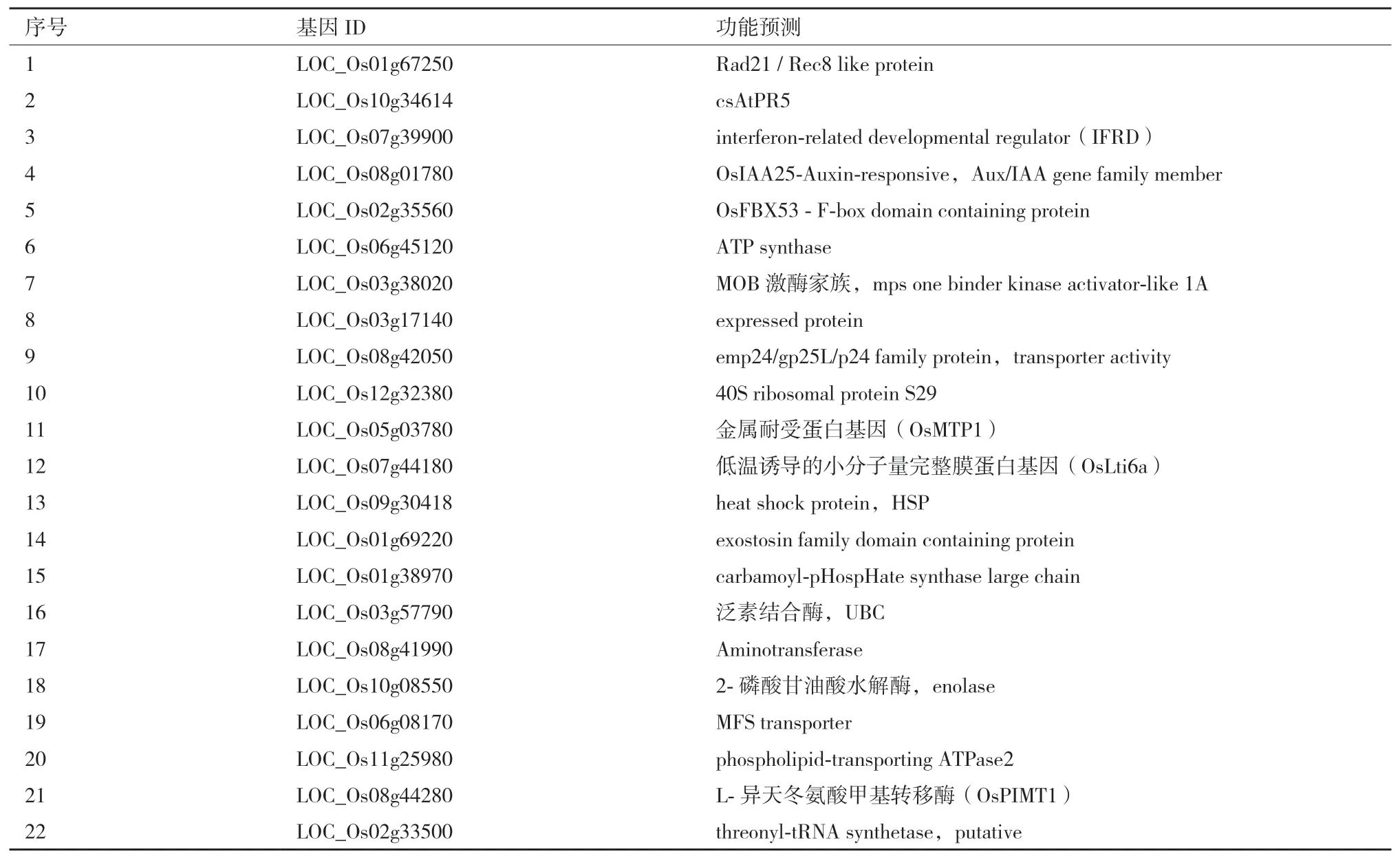
表4 候选蛋白的功能分析

表5 膜系统筛选出的候选互作蛋白的自激活检测
2.2 核系统酵母双杂交筛选与LCD互作的蛋白
2.2.1 核系统条件下诱饵蛋白自激活检测 将重组质粒pGBKT7-LCD转化酵母AH109,并以转化空载体pGBKT7的AH109作为对照,涂布在SDTrp和SD-Trp-His平板上。结果(图4)表明,转化pGBKT7和pGBKT7-LCD的酵母都能在SD-Trp的平板上正常生长,说明pGBKT7-LCD表达的融合蛋白对AH109酵母细胞的生长无毒性。且转化pGBKT7和pGBKT7-LCD的酵母都不能在SD-Trp-His平板上生长。同时,将pGBKT7-LCD和pGADT7共转化酵母AH109,涂布于SD-Trp-Leu和SD-Trp-Leu-His平板上,结果表明,酵母可以在SD-Trp-Leu平板上生长,但不能在SD-Trp-Leu-His平板上生长。综合转化试验结果,说明pGBKT7-LCD表达的融合蛋白不具有激活报告基因(His)的作用,可用于后续筛库。

图3 LCD与候选蛋白在膜系统酵母中的互作验证
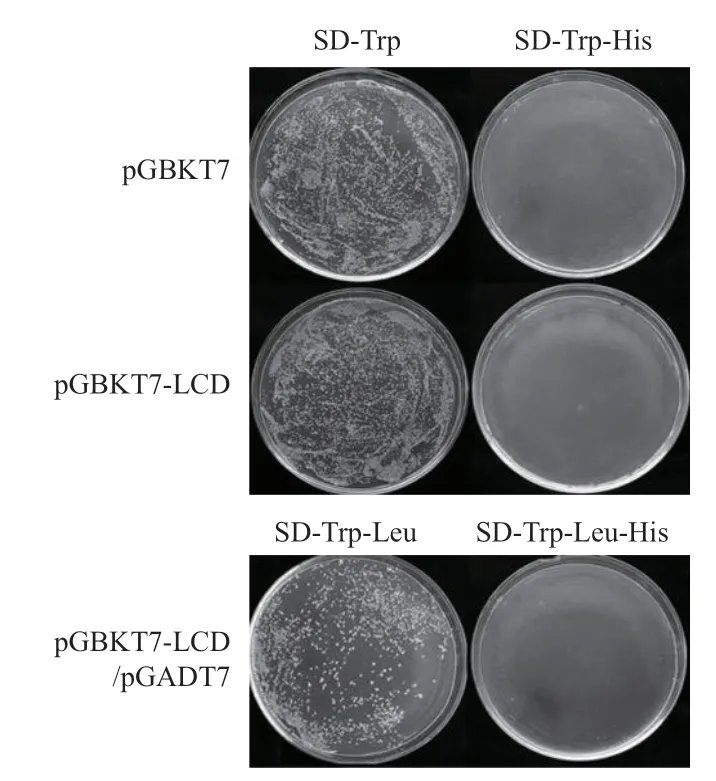
图4 核系统条件下诱饵蛋白LCD在酵母AH109中的自激活检测
2.2.2 pGADT7-prey和pGBKT7-LCD的共转化和互作蛋白筛选将pGADT7-prey和pGBKT7-LCD共转化酵母AH109,涂布于SD-Trp-Leu-His-Ade平板上,共标记51个阳性克隆。将51个菌落划线至SD-Leu-Trp-His-Ade+X-α-gal平板上(图5)发现,共有43个菌落呈蓝色,说明LCD与43个蛋白在酵母中互作,不能生长和不能变蓝的菌落视为没有互作,予以舍弃。
2.2.3 阳性AH109酵母菌中pGADT7插入片段的获得和分析通过对2次筛选出的43个阳性菌落扩增,得到38个文库质粒pGADT7中插入的cDNA片段,5个菌落经多次扩增都不能扩增出片段,予以舍弃。将扩增出的38个PCR片段进行测序,去除重复序列,共得到25序列,将得到的25个测序结果利用NCBI在线比对工具BLAST(http://www,ncbi.nlm.nih.gow/BLAST/)逐一进行基因相似性比较,查找同源基因,去除同源基因读码框与pGADT7插入片段的读码框不一致的序列,最终共得到7个UniGene(表6),编号为1-7。其中,5、6、7与植物抗逆和物质的运输有关;2、3、4与细胞生理代谢相关。

图5 核系统条件下LCD互作的阳性酵母显色筛选
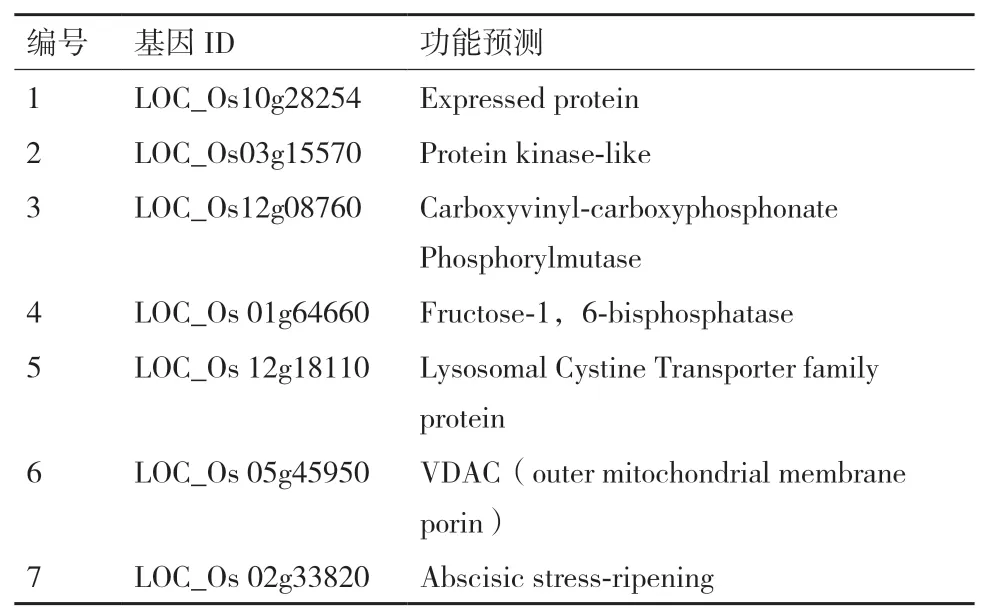
表6 核体系下筛选的LCD候选互作蛋白
2.2.4 AH109酵母中LCD与互作蛋白的验证 克隆筛选到的7个基因CDS全长序列,构建重组质粒pGADT7-1-7,并按照2.1的方法进行自激活检测。发现1、4基因存在自激活现象,予以舍弃,2、3、5、6和7不存在自激活现象,可以进一步进行互作验证。
将pGBKT7-LCD分别与pGADT7-2、pGADT7-3、pGADT7-5、pGADT7-6、pGADT7-7共同转化酵母AH109,发现2、3、5、6、7均可以在SD-Leu-Trp-His-Ade长出白色酵母菌落,说明2、3、5、6、7与LCD在酵母中存在互作。进一步将在SD-Leu-Trp-His-Ade营养缺陷板上长出的白色酵母菌落转移至SD-Leu-Trp-His-Ade+X-α-gal板,发现5个编号的转化菌株均能生长并呈现蓝色(图6)。因此,最终筛选得到的互作蛋白为LOC_Os03g15570、LOC_Os12g08760、LOC_Os 12g18110、LOC_Os 05g45950和LOC_Os 02g33820。

图6 LCD与候选蛋白在AH109酵母中的互作验证
3 讨论
近年来,已经有20多个介导水稻Cd积累的相关基因被鉴定,除LCD之外,大多数基因都直接参与Cd的吸收与转运过程,主要是一些转运Fe、Mn、Zn、Ca等离子的运输蛋白,如Nramp5介导水稻Mn/Cd的吸收[37];OsHMA3在根部细胞的液泡膜上,把Zn/Cd转运到液泡中解毒,维持水稻中Zn的稳态[16,38];OsCCX2在水稻节部调节Ca/Cd的分配[24]。除了运输蛋白,一些富含半胱氨酸的蛋白或多肽,能螯合Cd和解Cd等重金属毒害,如植物螯合肽(PC)及其谷胱甘肽(GSH)等前体、金属硫蛋白(MT)等。这些蛋白和多肽的合成和转运,对水稻对Cd的分配以及Cd毒害耐性有重要影响,如OsLCT1很可能能转运PC的前体,调控水稻GSH稳态,在As/Cd解毒中起重要作用[22]。防卫素蛋白如CAL1等也富含半胱氨酸,能螯合胞质中的Cd并分泌到木薄壁细胞外,促进Cd通过导管从根部到叶片的长距离运输[26]。此外,还有一些调控基因如转录因子、miRNA等,在调控水稻吸收、运输、分配Cd的过程中起作用,最终影响Cd在水稻不同部位的分配和积累[39]。而LCD定位于细胞核和细胞质中,既不是离子运输蛋白,也不是富含半胱氨酸的蛋白,也没有典型的结构域,推测LCD可能通过与其他蛋白互作,调控某个(或某些)Cd转运蛋白的转运活性,但目前对其介导Cd转运的机制还不清晰。
本研究以水稻一周的根、地上部分,分蘖期的根、叶,开花期的根、叶、节、节间、花均量混匀为材料,分别构建DUALmembrane系统和Yeast Two-Hybrid系统cDNA文库,并以LCD为诱饵,初步筛选到了21个与其互作的重要蛋白,分别属于转运蛋白、逆境响应相关蛋白、生长发育相关蛋白、蛋白激酶、生理代谢相关等及一些未知功能蛋白。与LCD互作的转运蛋白中,有MTP、MFS(Metal facilitator superfamily)等转运蛋白家族的成员,如LOC_Os05g03780、LOC_Os06g08170、LOC_Os08g42050等,推测水稻受到Cd毒害之后,LCD可能通过与相关转运蛋白相互作用,调控转运蛋白的转运活性,从而影响Cd在水稻中的分配和积累。其中,LOC_Os05g03780(OsMTP1)已经被报道参与Cd的运输。OsMTP1属于阳离子扩散促进剂(CDF)蛋白家族,广泛存在于细菌、真菌、植物和动物体内。在酵母或烟草中异源表达OsMTP1能够增强Cd积累和对Cd的耐受性。dsRNAi水稻转基因幼苗对重金属敏感性改变,同时成熟水稻不同器官中金属的积累量也发生了变化[30,40]。最近,一个MFS成员OsCd1已被鉴定,参与水稻Cd吸收与籽粒Cd积累[40]。LCD互作蛋白中,LOC_Os06g08170是一个MFS新成员,其是否参与水稻的Cd吸收与转运有待进一步的研究。
蛋白激酶、热激蛋白、ABA相关蛋白等在植物生物与非生物逆境胁迫反应中都起着重要的作用,如Cd能通过诱导活性氧(ROS)的产生,激活2个玉米蛋白激酶(ZmMPK3和ZmMPK6),来响应Cd胁迫[41]。早前也有研究发现水稻的这两个蛋白激酶(OsMPK3和OsMPK6)受Cd诱导激活,且该激活需要钙依赖的蛋白激酶(CDPK)和磷脂酰肌醇-3激酶(PI3kinase)的参与,增强植物的耐Cd能力[42]。另外的研究发现,异源表达一个水稻热激蛋白(OsMSR3),能够增加拟南芥耐Cd、Cu毒害的能力[43-44]。与LCD互作的蛋白中,也有蛋白激酶(LOC_Os03g38020和LOC_Os03g15570)、热激蛋白(LOC_Os09g30418)等,其与LCD相互作用的具体机制还需进行深入研究。
由于酵母双杂存在一定的局限性,本研究获得的互作蛋白还是一个初步结果,下一步还需采用其他方法,对初筛出的21个蛋白作进一步互作分析,如利用双分子荧光互补、免疫共沉淀等方法对LCD和21个候选蛋白之间的互作进行鉴定。此外,还需要构建基因敲除突变体等遗传材料,对候选互作蛋白在Cd积累中的遗传功能开展研究,多方面探讨LCD参与的Cd离子转运、积累机制。
4 结论
利用水稻膜系统和核系统酵母双杂交cDNA文库,筛选和鉴定了21个与LCD相互作用的蛋白,参与植物对逆境响应、离子转运等途径,推测LCD与其中某些蛋白共同作用调节水稻Cd积累。
