樟叶越桔嫩枝1株内生真菌的鉴定及抑菌活性测定
2020-12-04严冬曾为林罗旭璐陈肖学刘惠民赵平
严冬 曾为林 罗旭璐 陈肖学 刘惠民 赵平,3
(1.西南林业大学西南地区林业生物质资源高效利用国家林业和草原局重点实验室,昆明650224;2.西南林业大学西南山地森林资源保育与利用教育部重点实验室,昆明650224;3.云南森林资源培育与利用协同创新中心,昆明650224)
植物内生真菌是一类在健康植物组织内定殖,但不引起病害症状的真菌,并与宿主建立复杂的相互作用[1-2]。大量研究表明,许多经济林木、药用植物、农作物等植物中均分布着极为丰富的内生真菌,浸染了内生真菌的植物具有生长快速、抗逆性增加、抗病虫能力增强等特性,比未感染内生真菌的植物更具生存的竞争力[3-5]。自从1993年首次发现源自短叶红豆杉(Taxus brevifolia)韧皮部分的1株内生真菌(Taxomyces andreanae)能够生产抗肿瘤活性成分紫杉醇以来,植物内生真菌的研究受到了极大的关注,并成为获取活性天然产物的重要来源之一[6-7]。拟盘多毛孢属(Pestalotiopsis)为半知菌门中一类无性型真菌,是植物内生真菌中最重要的类群之一[8]。Lee等[9]从粗榧(Cephalotaxus sinensis)中分离得到了内生小孢拟盘多毛孢(P.microspora),该内生菌产生的植物毒素对枯草芽孢杆菌(Bacillus subtilis)具有较强的抑制作用。迄今为止,从拟盘多毛孢属内生真菌中发现结构新奇的化合物超过130多种[10-12],在医药、农业和林业领域方面具有潜在的应用价值[13-17]。
樟叶越桔(Vaccinium dunalianum)为杜鹃花科(Ericaceae)越桔属常绿灌木[18],主要分布于云南、贵州、四川、西藏等地[19],具有清热解毒、活络散瘀等功效[20],据记载从明代起就利用其幼嫩叶芽作为茶代用品长期饮用至今。Zhao等[21]发现樟叶越桔富含咖啡酰熊果苷类物质,但其高含量蓄积是否与内生真菌协助增强宿主抗逆性有关尚不明确。越桔属植物内生真菌多集中于菌根真菌的分离及其多样性研究[22-24],而对樟叶越桔内生真菌的研究鲜有报道,仅樊苗苗等[25]发现樟叶越桔叶内生真菌对小麦雪腐病菌(Typhla incarnata)和三七丝核病菌(Verticillium cinnabarium)等的拮抗作用较强,陈肖学等[26]发现樟叶越桔叶内生真菌杂色曲霉(Aspergillus versicolor)的发酵液对棉花枯萎病菌(Fusarium oxysporium sp.vasinfectum)等5种农作物病原真菌具有较强的抑制作用。本研究采用形态鉴别和分子生物学手段对樟叶越桔嫩枝分离得到的1株内生真菌Z-78进行了鉴定,采用平板对峙法评价了其对绵腐卧孔菌(Poria placenta)等3种木腐菌和禾谷镰孢(Fusarium graminearum)等5种植物病原真菌的抑制活性,并进行了Z-78发酵液抑制芸苔链格孢的酸碱稳定性试验,以期为该内生真菌进一步的开发利用提供参考。
1 材料与方法
1.1 材料
1.1.1 供试菌株 供试内生真菌Z-78从云南省武定县采集的樟叶越桔健康嫩枝中分离纯化得到。供试病原真菌菌株灰葡萄孢(Botrytis cinerea)CGMCC3.3790、腐皮镰孢(Fusarium solani)CGMCC3.2889、禾谷镰孢(F.graminearum)CGMCC3.4733、尖孢镰孢(F.oxysporum)CGMCC3.3633和芸苔链格孢(Alternaria brassicicola)CGMCC3.7805购自中科院微生物研究所菌种保藏中心,密粘褶菌(Gloeophyllum trabeum)CFCC86617、绵腐卧孔菌(Poria placenta)CFCC5608和采绒革盖菌(Coriolus versicolor)CFCC5336购自中国林业微生物保藏管理中心,均保藏于西南林业大学西南山地森林资源保育与利用教育部重点实验室。
1.1.2 培养基PDA培养基用于供试Z-78菌株和病原真菌培养,PDB培养基用于Z-78菌株发酵。
1.2 方法
1.2.1 内生真菌Z-78的鉴定 在超净工作台上将菌株Z-78接种到PDA培养基上,于28℃的恒温培养箱中培养,待菌落长满后观察菌落的形态、菌丝、颜色、大小等,在光学显微镜下观察菌丝及孢子形态。参照陈肖学等[26]的方法,采用Omegabiotek高纯真菌DNA提取试剂盒提取菌株Z-78的DNA,采用真菌通用引物ITS1(5'-AGAAGTCGTAACAAGGTTTCCG TAGG-3')和ITS4(5'-TCCTCCGCTTATTGATATGC-3')进行PCR扩增。扩增体系:DNA模板4.0 μL,ddH2O 6.5 μL,ITS1 1.0 μL,ITS4 1.0 μL,2×Taq PCR maker Mix 12.5 μL。循环体系:第1阶段1循环,94℃ 3 min;第2阶段35循环,94℃ 30 s,51.5℃ 30 s,70℃ 1 min;第3阶段1循环,70℃ 10 min。回收产物经电泳检测后送上海生物工程有限公司进行序列测定,测序结果与GenBank上登录的已知序列在Clustal X1.83软件上进行同源性比对后,采用PAUP 4.0软件构建系统发育树。
1.2.2 菌株培养 在超净工作台中将已灭菌(121℃、22 min)的PDA培养基倒平板,每平板约20 mL的培养基,待冷却凝固后用接种针挑取冷冻管里面保存的菌丝体接种于平板正中心位置,于28℃恒温培养箱中培养7 d。用5 mm打孔器在培养7 d后的菌落边缘打孔,取带有菌丝的菌饼接种到新的PDA培养基上培养。发酵培养时,培养7 d的Z-78菌株用5 mm的打孔器在菌落的边缘打孔切取直径为5 mm的圆形菌饼,接种于体积为100.0 mL的液体培养基中,在恒温振荡器中振荡培养7-10 d,培养条件28℃、180 r/min。用121℃灭菌后的纱布进行过滤,发酵液用0.22 μm无菌注射器过滤器进行过滤,置于4℃条件下保存。
1.2.3 抑菌活性测试 采用五点对峙培养法[25,27],测定Z-78菌株对供试病原菌的拮抗作用。用二代菌丝进行抑菌实验,每个带有PDA培养基的培养皿中分别放入5个菌饼,培养皿中间接入植物病原真菌,以病原菌为中心十字交叉距病原真菌2.5 cm处的4个点分别接入植物内生真菌,以在培养基中心接入植物病原真菌作为空白对照组,每个处理3个平行,恒温培养,定时观察,从对峙培养的第3天起用十字交叉法测量抑菌带距离和菌落的直径,直到空白长满时停止测量。按公式计算抑菌率:抑菌率=(对照菌落直径-处理菌落直径)/对照菌落直径×100%。参照刘泽星等[28]的方法,将抑菌率<25%标记为“+”,25%≤抑菌率<50%标记为“++”,抑菌率≥50%标记为“+++”,“-”表示没有抑制作用。
1.2.4 内生真菌Z-78发酵液的酸碱稳定性测定 参考陈肖学等[26]和张艳军[29]的方法,对菌株Z-78进行液体发酵,其发酵液分别用NaOH和HCl调至pH为2.0、4.0、6.0、8.0、10.0,静置24 h之后,采用生长速率法[26,30],取4.0 mL处理后的菌株发酵液加到6.0 mL PDA培养基中,以不加发酵液的平板为空白对照,以未做处理发酵液的平板为条件对照,以对环境适应能力较强的芸苔链格孢[31]作为指示菌,28℃条件下培养,分别从培养第2、4、6、8、10天后测量数据,按公式计算抑菌率。抑菌率=(对照组供试菌净生长半径-处理组供试菌净生长半径)/对照组供试菌净生长半径×100%。
2 结果
2.1 内生真菌Z-78的鉴定
观察菌株Z-78的形态与柠檬拟盘多毛孢(P.citrina)相似[32],菌落呈轮纹状均匀的向外扩长,开始生长是白色,一段时间后变为淡黄色,背面呈暗黄色。分生孢子团细小黑点状,散生在菌丝表面,为梭形5细胞,稍弯曲,(16.5-23.1)×(5.6-7.3)μm;中间3个色胞几乎同色,第3色胞浅褐色,上2色胞褐色,分隔处稍溢缩,长6.5-14.5 μm;顶胞无色,梯形,顶端有附属丝2-3根,长10.0-20.0 μm,菌落、菌丝及孢子结构见图1。参考《中国真菌志》[33],上述形态特征与木防己拟盘多毛孢菌(Pestalotiopsis cocculi)基本一致[34]。
在NCBI中采用BLAST对Z-78的ITS序列进行比对,用于同源性分析的序列分别是:木防已拟盘多毛孢(P.cocculi)KY703411、弥泽那拟盘多毛孢(P.menezesiana)JN198523[35]、AY687302、芒果拟盘多毛孢(P.mangiferae)KU194426、KX778664、聚多拟盘多毛孢(P.sydowiana)FJ478105、JQ322999、P.diploclisia KM199320[36]、KM199315、杭州拟盘多毛孢(P.hangzhouensis)AY526871、AY526870、小孢拟盘多毛孢(P.microspora)KT372347、KX755256和棒状拟盘多毛孢(P.clavata)KP050596、KC537803,外源序列选择不同科不同属的白座蕉孢壳(Diatrype stigma)KT004563[37]。基于rDNA-ITS构建的系统发育树如图2所示,Z-78与木防已拟盘多毛孢聚为一支,结合上述形态特征及分子鉴定结果,菌株Z-78鉴定为拟盘多毛孢属木防已拟盘多毛孢[Pestalotiopsis cocculi Vuill(Guba)G.C.Zhao &N.Li]。

图1 菌株Z-78的正面(a)、背面(b)生长状况、菌丝(c)和产孢结构(d)
2.2 内生真菌Z-78对病原菌的拮抗作用
由表1可知,内生真菌Z-78对8种不同供试病原菌呈现出不同程度的抑菌活性,抑菌率在50%-72.22%之间,表明菌株Z-78对供试病原菌具有较强拮抗能力,其拮抗效果见图3所示。在3种木腐菌中,Z-78对密粘褶菌的抑菌活性最强(图3-A),对绵腐卧孔菌(图3-B)次之,对采绒革盖菌(图3-C)相对较弱,抑菌率分别为72.22%、70.39%和57.84%。Z-78对5种常见农作物病原真菌的抑菌活性差异不大,按强弱顺序依次为尖孢镰孢(图3-D)>禾谷镰孢(图3-E)>芸苔链格孢(图3-F)>腐皮镰孢(图3-G)>灰葡萄孢(图3-H),抑菌率分别为61.76%、61.11%、61.00%、52.35%和50.00%。
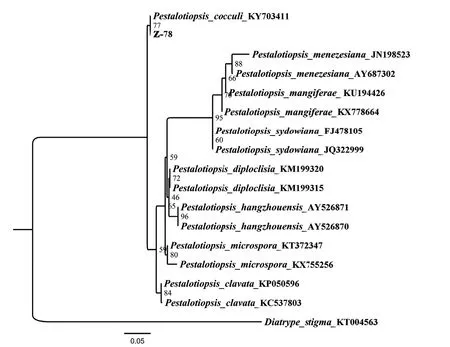
图2 基于内生真菌Z-78 rDNA-ITS序列的系统发育分析
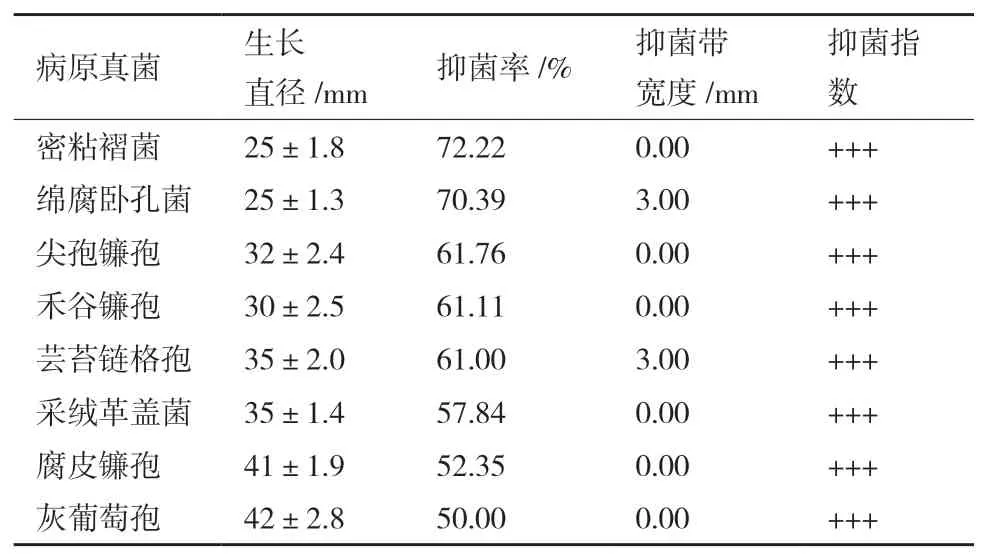
表1 内生真菌Z-78对8种植物病原菌的抑菌率
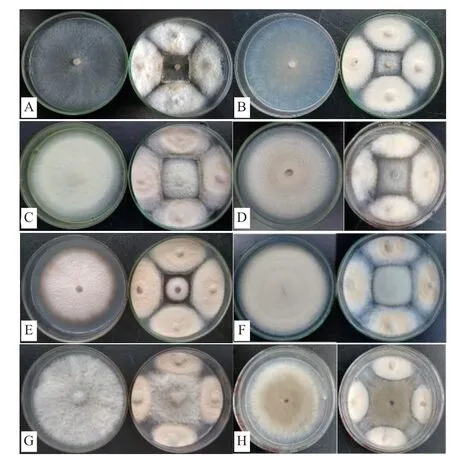
图3 内生真菌Z-78对8种植物病原菌的拮抗效果
从表1中可以看出,多数对峙结果中的抑菌带宽度均为0 mm。例如,在菌株Z-78与腐皮镰孢的对峙培养中(图3-G),两者菌落之间没有产生抑菌带,但菌株Z-78菌落随着生长时间增加,逐渐覆盖腐皮镰孢菌落,导致腐皮镰孢的生长受到了抑制。而有些病原菌株的菌落生长速度缓慢(图3-E),菌株Z-78可能释放一些代谢产物到培养基中,或产生一些挥发性抑菌气体,使得禾谷镰孢生长受制。并形成较宽的抑菌带。从图4可以看出菌株Z-78对8种植物病原菌的拮抗作用随着时间的推移逐渐增强,当抑制率达到顶峰时趋于一个相对平稳的状态,说明抑菌效果相对稳定。同时,也发现菌株Z-78对8种植物病原菌在不同培养时间也呈现出不同程度的抑菌活性。
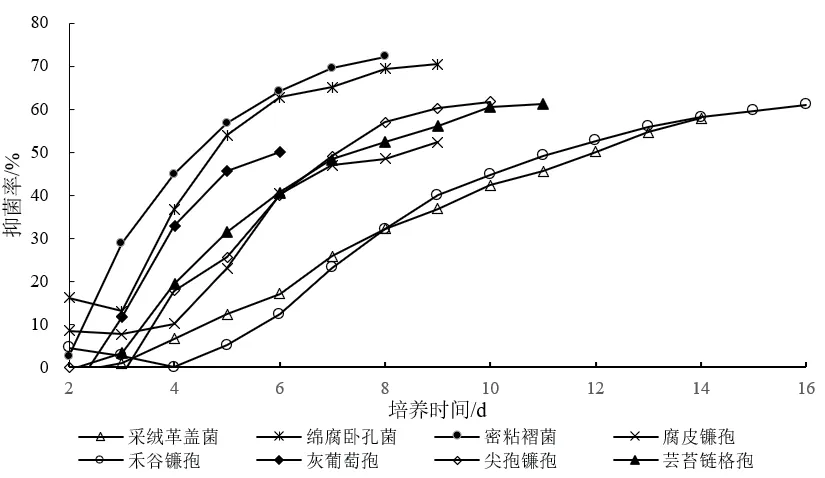
图4 内生真菌Z-78对8种植物病原真菌的拮抗动态变化
2.3 内生真菌Z-78发酵液的酸碱稳定性试验结果
内生真菌Z-78不同pH发酵液在不同培养时间下对芸苔链格孢的抑制作用,如表2所示。从表2可知,内生真菌Z-78不同pH发酵液在不同培养时间下对芸苔链格孢的抑制率均有不同程度的变化但差异不大,且变化趋势一致,即抑菌率从开始逐渐增大,到第8天达到最大值后逐渐减小。Z-78发酵液的初始pH值是5.0-6.0,当pH 6.0时,其对芸苔链格孢的抑菌率为55%,五点对峙培养法所得的抑菌率为61%(表1),说明两种方法所得的抑菌率较为接近。当发酵液pH达到10.0时,第8天的抑制率达到最大值为60%,pH 6.0以下时的最大抑制率为57%,表明其抑制作用波动不大。从培养第10天的抑菌率来看,除pH 8.0外其他pH发酵液的抑菌率均大于50%,显示出Z-78发酵液对芸苔链格孢具有较强的抑制作用。在添加了4.0 mL pH值为2.0、4.0、6.0、8.0、10.0的无菌蒸馏水的平板对照组中,病原真菌的菌落生长未受到明显的影响,表明Z-78发酵液对芸苔链格孢的抗菌活性较为稳定,不会受到酸碱性的影响。
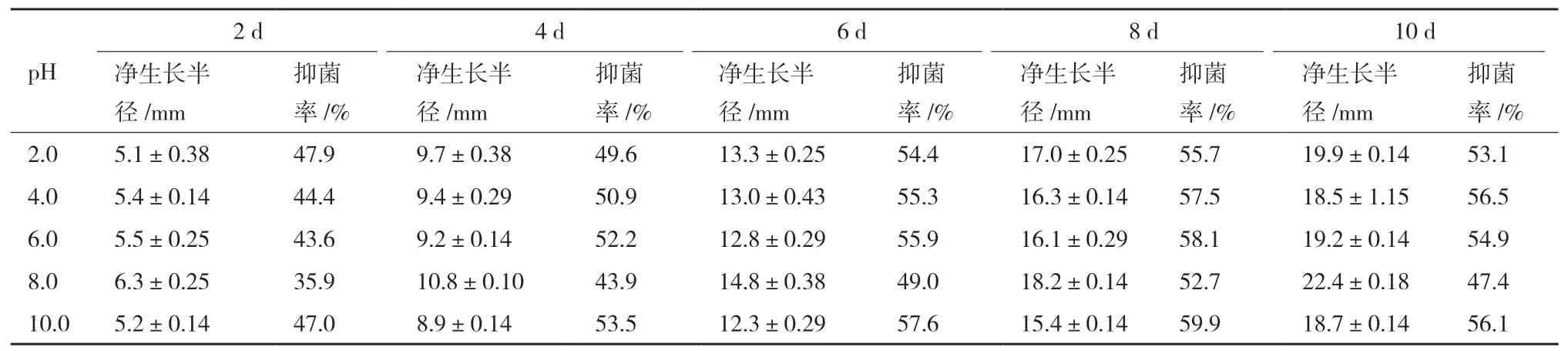
表2 不同pH条件下Z-78发酵液对芸苔链格孢的抑制作用
3 讨论
研究结果表明,大约有30%的内生真菌能够生产抗菌活性化合物,可对多种植物病原真菌具有专一拮抗或广谱抗性[38],其拮抗机制通常包括分泌抑菌物质、产生抑菌挥发性气体、竞争作用、重寄生作用等[39]。拟盘多毛孢是植物内生真菌的重要类群,次生代谢产物种类丰富,具有抗癌、抗菌等生理活性,具有良好的应用开发前景[8,13]。易晓华等[40]报道源自除虫菊(Pyrethryum cinerariifolium)叶拟盘多毛孢属内生真菌Y1菌株对苹果炭疽病菌(Glomerella cingulata)、灰葡萄孢、玉米大斑病菌(Exserohilum turcicum)等6种植物病原真菌的抑制作用达到85%以上。李靖等[41]发现石楠锈孢锈菌(Aecidium pourthiaea)重寄生拟盘多毛孢(Pestalotiopsis sp.)对辣椒炭疽病菌(Sclerotinia sclerotiorum)等10种常见植物病原真菌具有一定的抑制作用,李向楠等[42]也发现源自云南热区的多种该属内生真菌显示出广谱抗菌活性,表明不同来源拟盘多毛孢属内生真菌具有较为显著的抑菌活性。Li等[43]报道源自欧洲红豆杉(Taxus baccata)、落羽杉(Taxodium distichum)、佛罗里达榧树(Torreya taxifolia)和石斛(Dendrobium sp.)的内生小孢拟盘多毛孢(P.microspora),以及从澳大利亚瓦勒迈杉(Wollemia nobilis)分离的斑污拟盘多毛孢(P.maculans)的液体发酵液含有的环己烷类物质,对腐皮镰孢、古巴镰孢(Fusarium cubense)、麦根腐长孺孢(Helminthosporium sativum)、蒂腐色二孢(Diplodia natelensis)等病原真菌具有抗菌作用。源自红树林海莲(Bruguiera sexangula)枝的污斑拟盘多毛孢(P.foedan)中的单萜内酯类对灰葡萄孢和烟草疫霉菌(Phytophthora nicotianae)[44],源自铁皮石斛幼苗的该属DO14菌株中的吡喃酮类物质对白色念珠菌(Candida albicans)、新型隐球菌(Cryptococcus neoformans)、红色毛藓菌(Trichophyton rubrum)和烟曲霉(Aspergillus fumigatus)[45],源自茶(Camellia sinensis)枝的无花果拟盘多毛孢(P.fici)中的吡喃酮类物质对禾谷镰孢[46],源自芒果(Mangifera indica)的芒果拟盘多毛孢(P.mangiferae)中的酚类物质对白色念珠菌[47]显示有较强的抗菌活性,表明不同来源该属内生真菌的抑菌活性物质基础及其抑菌机制值得进一步深入研究。
4 结论
本研究对樟叶越桔嫩枝1株内生真菌Z-78进行了形态鉴别和分子鉴定,将其确定为木防已拟盘多毛孢[Pestalotiopsis cocculi(Guba)G.C.Zhao & N.Li]。采用五点对峙法评价其对密粘褶菌等3种木腐菌、尖孢镰孢等5种常见植物病原真菌的拮抗活性,结果表明菌株Z-78对8种供试病原菌的生长均具有较强的抑制作用,其中对密粘褶菌的抑菌活性最强,并发现Z-78发酵液在酸碱性条件下对芸苔链格孢的抑菌活性较为稳定,研究结果将为樟叶越桔枝内生真菌木防已拟盘多毛孢中抑菌活性物质进一步的分离鉴定和开发利用提供了参考。
