强冬性区白菜型冬油菜品质和产量相关性状变异分析
2020-11-19郑国强郭敏明魏家萍米文博徐春梅徐明霞曹小东方新玲崔小茹董小云陈其鲜刘自刚
郑国强,郭敏明,魏家萍,米文博,徐春梅,徐明霞,邹 娅,曹小东, 方新玲,崔小茹,董小云,米 超,陈其鲜 刘自刚,
(1.甘肃农业大学农学院/甘肃省干旱生境作物学重点实验室/ 甘肃省作物遗传改良与种质创新重点实验室,兰州 730070; 2.甘肃省武山县农业技术推广中心,甘肃 武山 741300; 3.甘肃省农业技术推广总站,兰州 730070)
油菜为十字花科芸薹属作物[1],油菜籽油是我国主要食用油的来源。油菜是我国种植面积最大、产油量最高、发展潜力最大的油料作物,油菜的大面积种植和稳定发展对我国植物类食用油安全至关重要[2]。我国有冬油菜和春油菜两大产区[3-4],甘肃大部分地区属于典型的春油菜产区,冬季及其寒冷、干旱少雨、植被稀疏土壤沙化严重,生态环境恶劣,不适合冬油菜的种植[5]。刘自刚等[5]选育了强抗寒冬油菜甘油系列品种,使甘肃地区生产冬油菜成为现实。与春作物相比,冬油菜具有增加冬、春季地表植被的覆盖度、有效减少沙尘来源、缓解沙尘危害、改善生态环境条件等优点[6]。同时,冬油菜的含油率和产量等经济性状也远远高于同类春播油料作物[7],而且冬油菜的种植可为部分光照充足地区创造复种条件,有利于改革传统种植制度[8],提高农业复种指数与单位面积经济效益,这对北方地区农业生产具有极其重要的经济和生态意义。
油菜的品质和产量相关性状均属于数量遗传学范畴,控制这些性状微效基因非常多, 这些基因通过显性效应、加性效应、上位性效应以及基因与环境的互作效应在植物体内形成一个非常复杂的调控网络。刘自刚等[5]、李雁等[9]、侯献飞等[10]、张彦红等[11]研究发现,不同生态条件对作物性状表现、产量形成、品质性状等具有很大影响,且不同品种对生态区变化的反应也存在差异。20世纪80年代,黄继英等[12]提出了冬油菜北移种植的设想,并开展了冬油菜耐寒品种的引育、配套栽培技术研发等系列工作,育成/筛选了秦油3号、甘白油菜等10余个较耐寒的甘蓝型油菜品种,研制了“沟播+冬苫”安全越冬栽培技术,成功将甘蓝型冬油菜从黄淮河、关中灌区(北纬35°以南、海拔700 m以下),北移至陕西洛川(北纬35°46′、海拔1 200 m)、山西临汾(北纬36°20′)、河北石家庄(北纬38°00′)等地种植,替代了原来种植的白菜型冬油菜,产量从750 kg·hm-2大幅提升至2 250~3 000 kg·hm-2[12-15]。在全球气候变暖、农膜技术[16]广泛应用、更耐寒甘蓝型冬油菜品种甘杂1号的育成等综合因素作用下,促进了甘蓝型冬油菜北移技术的应用。白菜型冬油菜具有更优异的抗寒性[16],一直是高纬度高海拔地区油菜种植的主要类型。陈娇荣等[17]通过筛选抗寒品种等措施,将白菜型冬油菜引入北方旱寒区更高海拔、更高纬度种植,对季节性风蚀农田有良好覆盖效果[18-19]。开展冬油菜北移后农艺性状、品质和产量性状变化的研究,能够为进一步选育和筛选出适应北移区生态条件、适应机械化生产的高产优质冬油菜品种提供有力依据。本试验利用11个品种(系)在甘肃从南至北的5个试点种植,研究原产区与北移区试点的农艺性状、品质和产量性状的变化规律,以期为冬油菜的北移和选育适应北移区生态条件的新品种提供科学的支撑。
1 材料与方法
1.1 材 料
试验材料由甘肃省干旱生境作物学国家重点实验室提供的10个白菜型选育品系(种)和天水农科院选育的Tianyou 4(ck)组成。
1.2 仪器与设备
近红外快速品质分析仪( Foss-NIR System),瑞典FOSS公司。
1.3 方 法
1.3.1试验设计
本试验于2018—2019年,在甘肃的天水、定西、会宁、榆中和武威等5个试验点进行,试验采用完全随机区组设计排列,3次重复。试验小区面积为3.6 m2,行距为20 cm,株距约10 cm,每小区种8行。试验过程总共施肥2次,播种前施20 kg·(667 m2)-1磷酸二铵作为基肥,冬季降雪后在雪面撒施5 kg·(667 m2)-1尿素。
1.3.2成熟期农艺性状的测定
成熟期收获时,在3个重复的其中一个小区中随机取3个植株,测定其株高、分枝部位、一次分枝数、二次分枝数、主花序有效长度、主花序有效角果数、角果长度、角粒数、千粒重、茎粗、根粗、根长[7]。
1.3.3品质性状的测定
考种时获得的油菜籽粒带回实验室,采用近红外分析仪( Foss-NIR System) 测定分析其品质性状,测定指标包括棕榈酸(C 16∶0) 、硬脂酸(C 18∶0) 、油酸(C 18∶1) 、亚油酸(C 18∶2) 、亚麻酸(C 18∶3) 、碳烯酸(C 20∶1) 、廿碳烯酸(C 20∶1)、芥酸(C 22∶1) 、硫苷(Glu)和含油量。

表1 白菜型冬油菜北移区与原产区(天水)农艺性状差异
全株角果数(个);LS为角果长度(cm);Seeds为角粒数(粒);TSW为千粒重(g);SD为茎粗(cm);RD为根粗(cm);RL为根长(cm)。下同。

表2 性状间的相关性分析
1.3.4小区产量的测定
小区内油菜角果70%~80%由绿变黄时(即蜡熟期)进行收割、脱粒、称重,最后折算产量。
1.3.5数据整理与分析
采用Excel软件和SPSS软件对实验数据进行处理与分析。
2 结果与分析
2.1 白菜型油菜农艺性状分析
2.1.1不同试点白菜型油菜植株农艺性状差异分析
与原产区(天水)试点相比,北移区试点不同品种的白菜型冬油菜农艺性状发生了变化,冬油菜北移后株高、分枝部位、主花序长度、主花序角果数、茎粗、根粗等农艺性状均降低或减少,全株角果数除会宁试点外的其余3个试点表现出减少的趋势;一次分枝、二次分枝、角果长度、角粒数、根长等具有明显增加的趋势,千粒重的变化趋势不太明显。
2.1.2不同试点白菜型冬油菜植株农艺性状间的相关性分析
从表2可知,白菜型冬油菜植株农艺性状之间存在明显的相关性,其中茎粗与根粗之间呈极显著正相关;株高与主花序角果数呈显著正相关,株高与角果长度,分枝部位与一次分枝,主花序角果数与角果长度,根粗和角果数呈显著负相关;株高还与主花序长度、全株角果数和分枝部位,一次分枝与全株角果数、根长,角粒数与根长等之间存在不显著的正相关;株高与二次分枝数,茎粗与千粒重等之间存在不显著的负相关。白菜型冬油菜的产量与农艺性状之间存在相关性,其中产量与全株角果数呈极显著正相关,说明角果数对产量的贡献最大,选择高产品种过程中应选择角果数较多的单株;产量还与主花序角果数、主花序长度、千粒重、根粗、茎粗、株高和一次分枝之间呈不显著正相关。
2.1.3农艺性状的差异分析
从表3可知,不同试点种植的白菜型冬油菜农艺性状的变异程度明显不同,分枝部位和二次分枝的变异系数最大,说明这11个品种(品系)的13个农艺性状中分枝部位和二次分枝的变异最为丰富,在不同环境间变异程度最大,表明分枝部位和二次分枝易受环境的影响;角果长度和角粒数的变异系数最小,表明这2个农艺性状在不同环境中较为稳定。在不同试点种植的冬油菜中,同一农艺性状在不同品种的变异性明显不同,如15 PW 16的根长变异系数为0.425,而16 QX 367 HX仅为0.188,Tianyou 4千粒重变异系数为0.068,而17 RLXⅡ38为0.257,表明不同品种对生态环境变化的反应存在明显差异。

表3 品种在不同试点间农艺性状的变异系数

表4 不同生态区白菜型冬油菜棕榈酸含量比较
2.2 白菜型冬油菜籽粒品质性状分析
2.2.1不同试点白菜型冬油菜籽粒棕榈酸含量比较
白菜型冬油菜北移后棕榈酸含量变化显著,从原产区(天水)北移至定西、会宁、榆中、武威试点后,棕榈酸含量表现出明显增加的趋势;方差分析表明,不同生态区白菜型冬油菜棕榈酸平均含量存在显著差异,天水试点的棕榈酸平均含量显著低于其余4个试点,其次是会宁、兰州、定西、武威;与ck相比,在各试点除个别品种外棕榈酸平均含量不存在显著差异(表4)。
2.2.2不同试点白菜型冬油菜籽粒C 18脂肪酸含量比较
白菜型冬油菜C 18脂肪酸主要有4种,即硬脂酸、油酸、亚油酸和亚麻酸。方差分析发现(表5),北移区硬脂酸、亚油酸含量低于原产区,油酸、亚麻酸含量高于原产区;同一试点的不同品种之间C 18脂肪酸含量存在差异,天水试点6个品种硬脂酸含量和8个品种油酸含量低于ck,8个品种的亚油酸含量和5个品种的亚麻酸含量高于对照品种;定西试点有7个品种的硬脂酸、8个品种油酸、6个品种亚油的含量高于对照品种,6个品种亚油酸低于对照品种;会宁试点有6个品种的硬脂酸、9个品种的亚麻酸含量低于对照,9个品种的油酸含量和所有品种的亚油酸含量高于对照品种;榆中试点硬脂酸含量均低于对照品种,5个品种的油酸、7个品种的亚油酸低于对照品种,8个品种的亚麻酸含量高于对照品种;武威试点8个品种的硬脂酸、9个品种的亚油酸含量高于对照品种,8个品种的油酸和9个品种的亚麻酸含量低于对照。
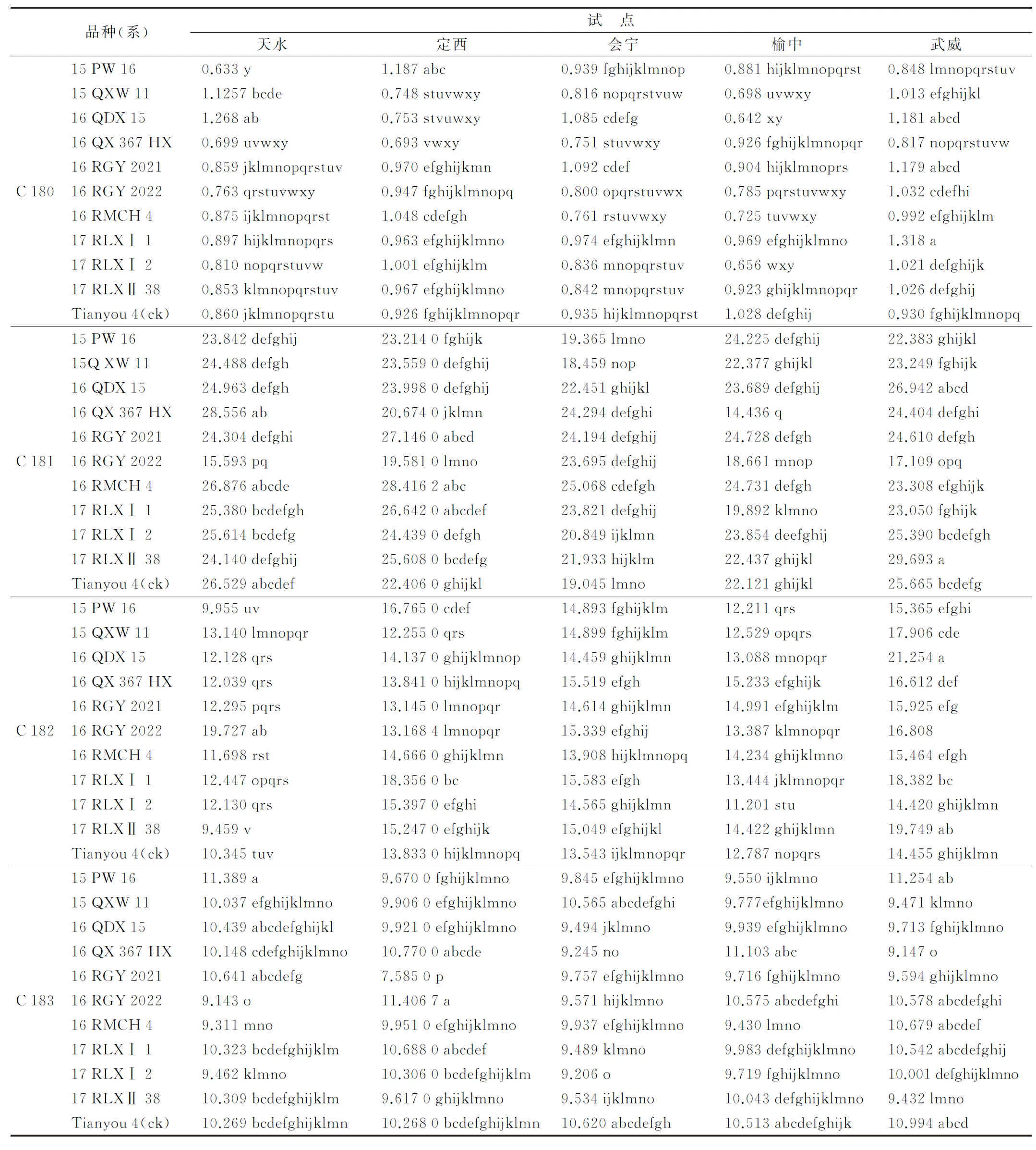
表5 不同试点白菜型冬油菜籽粒C 18脂肪酸含量比较
2.2.3不同试点白菜型冬油菜籽粒廿碳烯酸含量比较
白菜型冬油菜从原产区(天水)北移种植后,廿碳烯酸的平均含量呈现降低的趋势,方差分析表明(表6),天水试点的廿碳烯酸平均含量为12.398%,显著高于北移区试点;不同品种(系)的廿稀酸平均含量也存在显著差异;与对照品种相比发现,天水试点有7个品种廿碳烯酸含量升高,定西试点有6个品种、会宁试点有8个品种、榆中和武威试点廿碳烯酸含量均降低。
2.2.4不同试点白菜型冬油菜籽粒芥酸和硫苷比较
方差分析表明(表7),不同试点种植的白菜型冬油菜品种(系)芥酸含量存在显著差异,与原产区(天水试点)相比,北移区试点的平均芥酸含量呈现降低的规

表6 不同试点白菜型冬油菜籽粒廿碳烯酸含量比较

表7 不同试点白菜型冬油菜籽粒主要品质指标比较

表8 品质性状在不同品种间的变异系数和总变异系数

表9 品质性状在不同试点间的变异系数
律性变化(表4),武威试点的平均芥酸含量最低,为33.986%;硫苷含量在不同试点之间差异显著,会宁试点的平均硫苷含量明显高于其余4点。不同品种(系)之间平均芥酸含量和平均硫苷含量也存在明显的差异,16 RGY 2022的平均芥酸含量最高,为38.900%;16 RMCH 4的平均硫苷含量最高,为100.995%。与原产区相比,除会宁试点外,其余试点表现降低的趋势;与对照品种相比,新选育的部分品种芥酸含量明显较低,其中天水试点有7个品种,定西试点有4个品种,榆中试点有2个品种,武威试点有7个品种芥酸含量差异显著。在太水试点有3个品种,定西试点有3个品种,榆中试点有8个,武威试点有5个品种硫苷含量显著低于对照品种。
2.2.5不同试点白菜型冬油菜籽粒含油量比较
方差分析表明(表7),不同试点种植的白菜型冬油菜含油量存在显著差异,与天水相比,含油量除榆中试点外其余北移区试点呈降低趋势;榆中试点的平均含油量最高,为44.577%,武威试点的平均含油量为37.089%,是5个试点中最低的。在天水试点,16 RGY 2022的平均含油量明显低于其余品种,定西试点17 RLXⅠ 2平均含油量明显低于其余品种,会宁和榆中试点不同品种的含油量之间不存在显著差异,武威试点Tianyou 4平均含油量明显低于其余品种。在定西和武威有部分品种的含油量显著高于对照品种,除个别外的大多数品种的含油量与对照品种间无显著差异。
2.2.6品质性状在不同试点的差异分析
从表8可知,不同试点种植的白菜型冬油菜,不同品质性状的变异程度明显不同,其中棕榈酸和硬脂酸含量的变异系数大于芥酸和硫苷含量的变异系数;不同试点种植的不同白菜型冬油菜品种(系)同一品质性状的变异程度存在差异,其中芥酸在会宁试点不同品种中的变异系数为0.037,而武威试点的变异系数为0.178,表明在武威试点芥酸含量在不同品种之间变化较大。
变异性分析显示(表9),种植在不同试点的白菜型冬油菜品种同一品质性状变异系数明显不同,如含油率在16 QX 367 HX中的变异系数最小,为0.063,而在16 RGY 2022中变异较大,变异系数为0.186,表明16 QX 367 HX在不同环境中其含油率较为稳定,不易受环境的影响,而16 RGY 2022的含油率易受环境影响。同一品种不同品质性状的变异系数也不同,表明环境因子对不同品质性状的影响不同。
2.3 不同白菜型冬油菜品种原产区与北移区产量差异分析
与原产区(天水)相比,定西、榆中和武威3个北移区试点所有白菜型冬油菜品种产量均降低,会宁试点除17 RLXⅡ 38和Tianyou 4外的9个品种产量均有所增加(表10)。同一白菜型冬油菜品种在不同试点产量与原产区的产量差异有所不同,如17 RLXⅠ 1在武威试点降低103.44 kg·(667 m2)-1,而会宁试点增加38.35 kg·(667 m2)-1;在同一试点种植的不同白菜型冬油菜品种之间的产量变化也不同,如与原产区相比,定西试点的17 RLXⅡ 38产量降低119.12 kg·(667 m2)-1,15 PW 16的产量减低56.80 kg·(667 m2)-1。在定西试点除16 QX 367 HX、16 RGY 2022、1616 RMCH 4 和17 RLXⅡ 38外的其余品种产量均优于对照品种,会宁试点除17 RLXⅡ 38外的所有品种产量均高于Tianyou 4,榆中试点包括15 PW 16、15 QXW 11、16 QDX 15在内的7个品种北移后减产趋势弱于对照品种,武威试点除17 RLXⅡ 38外的9个品种北移后减产趋势弱于Tianyou 4;综上所述,新选育的大多数品种在强冬区的产量表现优于对照,推测新品种在该区域具有更强的推广潜力。

表10 不同白菜型冬油菜品种原产区与北移区产量差异分析
3 讨 论
对11个品种(系)白菜型冬油菜的14个农艺性状差异性分析表明,在主要农艺性状方面存在较为丰富的变异性,其中分枝部位和二次分枝在不同环境中的变异最为丰富,说明分枝部位和二次分枝易受环境的影响;角果长度和角粒数的变异系数最小,表明2个农艺性状在不同环境中较为稳定;白菜型冬油菜从原产区(天水)北移至定西、会宁、榆中和武威后,其表现出株高降低、分枝部位下降、主花序长度变短、主花序角果数减少、全株角果数除会宁外的其余3个试点也表现出减少的趋势;一次分枝和二次分枝增多,株形变差;角果长度、角粒数、等性状有所改善;这与刘自刚等[5]、刘海卿等[20]的研究结果相符。本研究发现,北移后冬油菜根粗和茎粗协同变小,根长略有增加。因此,以后选择北移引种时应该考虑选择株高较高、主花序长度较长、主花序角果较多、分枝部位较高、分枝数较少、根茎较粗的品种。北移后纬度和海拔升高,冬季最低气温更低,不利于油菜等作物的安全越冬,低温下油菜体内基因表达组发生重大变化,BnICE1[21]、ELI[22],CBF[23]、Cu/Zn-SOD[24]、APX[25]、BnMKK2[26]、BnHMGB2[27]、β-1,3-葡聚糖酶基因[28]等抗寒相关基因被表达,使得白菜型冬油菜适应冷胁迫和提高抗寒性,北移后农艺性状呈现有规律的变化是由于白菜型冬油菜抵御严寒的形态学结果。白菜型冬油菜植株不同农艺性状之间存在相关性,其中茎直径与根直径间呈极显著正相关,这与植物在生长过程中地上部分与地下部分相互促进和制约的理论相符[29];株高与主花序角果数之间存在显著正相关,且主花序角果数与产量之间呈不显著的正相关,因此主花序角果数可作为高产育种过程中的主要选择依据;分枝部位与一次分枝之间存在显著负相关,说明分支部位与一次分枝数之间相互抑制,分枝部位低的品种一次分枝较多,这一依据也可作为白菜型冬油菜未来选择育种的依据。
白菜型冬油菜的棕榈酸、硬脂酸、油酸、亚油酸、亚麻酸、廿碳烯酸、硫苷、芥酸和含油量在不同试点间差异显著;与原产区相比,北移区的硬脂酸、亚油酸、廿碳烯酸、芥酸和含油量均呈降低趋势,硫苷除会宁试点外表现降低趋势,棕榈酸、油酸、亚麻酸呈上升趋势。与对照品种相比,个别品种在品质方面具有更好表现,更加有利于食用油安全保证。Zhao J Y等研究认为,油菜脂肪酸生物合成先形成油酸,沿碳链延长方向合成廿碳烯酸、芥酸等;沿碳链去饱和方向合成亚油酸、亚麻酸[30]。油菜品质性状表现均受微效多基因修饰,在杂交分离群体中呈连续分布特征,环境因子对表型有重要影响[31]。推测各试点的纬度、海拔、温度、降雨量和光照等环境因子的不同是造成品质性状不同的主要原因。因此,在选择杂交亲本改善作物品质性状过程中剔除环境对其性状的影响显得格外重要。在不同的品种之间大部分品质性状差异显著,其主要由于不同品种间控制品质性状的微效多基因的遗传背景和调控不同。因此,可以采用品种间杂交等方式实现优良微效多基因的聚集,从而改善冬油菜品种品质性状表现。
与原产区(天水)相比,白菜型冬油菜北移区除会宁试点外产量均降低。与对照品种(Tianyou 4)相比,新选育的大多数品种在强冬区的产量更高、适应性更强、表现更好,更适合在强冬区推广种植,正好能够填补强冬区冬油品种单一的困境。通过相关性分析表明,白菜型冬油菜的产量性状与农艺性状之间存在相关性,其中产量与全株角果数之间呈极显著正相关,说明北移过程中产量的降低主要是由于全株角果数的减少造成的,这与徐进鸿等[32]、蒙祖庆等[33]、王毅等[34]的研究结果相符;产量还与主花序角果数、主花序长度、千粒重、根粗、茎粗、株高和一次分枝之间呈不显著正相关。因此,在选择适合北移区种植的冬油菜高产品种过程中,应加强对全株角果数、主花序角果数和千粒重等农艺性状的选择。另外,冬油菜北移种植后不仅仅体现在经济价值上,更重要的是其生态价值。因此,冬油菜北移仍具有巨大的发展潜力。
