标准切花菊分枝性状的杂种优势和混合遗传分析
2020-12-02于瑞宁陈发棣房伟民
于瑞宁 马 琦 孙 炜 奥 妮 张 飞 陈发棣 房伟民
(作物遗传与种质创新国家重点实验室/农业农村部景观设计重点实验室/南京农业大学园艺学院,江苏 南京 210095)
菊花(ChrysanthemummorifoliumRamat.)是菊科菊属多年生宿根花卉,是我国传统十大名花之一,花色花型丰富,观赏价值极高,深受广大人民的喜爱。切花菊是世界四大切花之一,包括小花型的多头切花菊和大花型的标准切花菊两种类型。分枝是切花菊的重要园艺性状,分枝数、分枝长度和分枝角度等分枝性状与切花菊的栽培方式和观赏品质紧密相关;特别是在标准切花菊生产中,需要人工抹除侧芽侧蕾,生产成本较高,且易伤及主蕾,影响成花品质[1]。因此,研究切花菊分枝调控机理、选育侧芽侧枝少的标准切花菊品种具有重要意义。
近年来,相关研究已围绕菊花分枝生理机制[2-4]和基因调控[5-7]等方面取得了良好进展,但其遗传特性仍不明确。一般认为,分枝性状属于多基因控制的数量性状[8-9]。菊花分枝性状的数量遗传学研究多集中于盆栽小菊[10]和多头切花小菊[11-13],而关于标准切花菊分枝性状的研究较少[14],遗传机制有待进一步研究。
主基因+多基因混合遗传模型分析方法是经典数量性状研究方法之一,广泛应用于菊花数量性状的遗传研究[14-16]。本研究对标准切花菊两个F1分离群体的总侧芽数、上部一级分枝数、一级分枝粗、一级分枝角度、一级分枝长度和总侧芽数/叶节数6个分枝性状进行杂种优势分析以及主基因+多基因混合遗传分析,探究不同遗传背景下分枝性状的遗传模型和主基因遗传效应,解析标准切花菊分枝遗传机制,以期为标准切花菊的分枝育种提供参考;同时,筛选获得优良的少侧芽F1株系可作为育种中间材料,对少侧芽侧枝标准切花菊品种选育和进一步研究具有重要意义。
1 材料与方法
1.1 试验材料
试验材料为标准切花菊品种精之一世、秦淮春雪、QD3-71和神马,均由中国菊花种质资源保存中心提供。其中,精之一世和秦淮春雪侧芽侧枝数较少,QD3-71和神马侧芽侧枝数较多。2016年秋季通过人工辅助授粉将4个品种配成QD3-71×精之一世(组合Ⅰ)和秦淮春雪×神马(组合Ⅱ)两个杂交组合,2017年春季将所得种子播种并扦插扩繁,分别获得97个和147个F1实生株系。
1.2 性状调查
2018年7月将组合Ⅰ和组合Ⅱ的F1群体及其亲本扦插定植,定植间距为10 cm×10 cm,定植后常规管理。于2018年11月盛花期观测两个组合F1群体及其亲本的6个分枝性状。总侧芽数、上部一级分枝数、一级分枝角度和一级分枝长度的统计参照杨信程等[13]的方法;一级分枝粗为上部一级分枝1/2处的直径,使用精度为0.01 mm的数显游标卡尺测量;总侧芽数/叶节数为总侧芽数与叶节数的比值。每株系单株重复3次,取平均值。
1.3 杂种优势分析
杂种优势用中亲优势(Hm)和中亲优势率(RHm)来表示。中亲值(mid-parents value,MPV)为两亲本某性状的平均值;Hm为F1群体某性状的平均值(Tm)与MPV之差;RHm为中亲优势Hm与MPV的比值,即Hm=Tm-MPV,RHm=(Tm-MPV)/MPV×100%。利用SPSS 25.0软件对F1群体性状值与MPV进行单样本均值t检验,检验中亲优势的显著性。
1.4 主基因+多基因混合遗传分析
2 结果与分析
2.1 杂交F1群体分枝性状的表型分布与杂种优势
由表1可知,两杂交组合分枝性状的变异系数为10.23%~39.92%,分离较广泛。两组合中一级分枝粗和一级分枝角度的变异较小,变异系数为10.23%~13.81%;总侧芽数、上部一级分枝数、一级分枝长度和总侧芽数/叶节数的变异较大,变异系数为22.79%~39.92%。由偏度、峰度可知,各个分枝性状呈正态分布趋势且连续性较好,符合数量性状分布的典型特征,适合对其进行进一步的遗传分析研究。
由表2可知,在组合Ⅰ中,上部一级分枝数、一级分枝粗和一级分枝长度的Hm均达到极显著水平,RHm分别为14.83%、-9.59%和18.14%,总侧芽数Hm达到显著水平,RHm为8.27%;在组合Ⅱ中,上部一级分枝数和一级分枝角度的Hm达极显著水平,RHm分别为-8.40%和-3.23%,一级分枝长度Hm达到显著水平,RHm为6.48%。从Hm的作用方向来看,在两组合中,一级分枝长度均表现为正向,总侧芽数/叶节数均表现为负向;总侧芽数、上部一级分枝数和一级分枝角度在组合Ⅰ中表现为正向,在组合Ⅱ中表现为负向;一级分枝粗在组合Ⅰ中表现为负向,在组合Ⅱ中表现为正向。总侧芽数、上部一级分枝数在两组合中均表现出一定程度的偏母性遗传。
在两个组合中,F1群体各分枝性状均存在正向或负向超亲个体,存在超亲分离现象。其中,组合Ⅰ的一级分枝长度平均值(11.38 cm)大于高亲本值(10.40 cm),形成了正向超亲优势,超亲优势值为0.98,超亲优势率为9.42%。两个组合的其他分枝性状平均值均介于双亲之间,未形成超亲优势。
2.2 标准切花菊各分枝性状的相关性分析
由Pearson相关分析结果(表3)可知,在15对分枝性状间,共有10对性状表现为极显著相关,2对性状表现为显著相关。其中,总侧芽数、上部一级分枝数、一级分枝长度与总侧芽数/叶节数两两之间呈极显著正相关。一级分枝粗与一级分枝角度之间呈极显著正相关,但与其他性状关系较为复杂:一级分枝粗与一级分枝长度呈极显著正相关,与总侧芽数/叶节数呈显著负相关;一级分枝角度与总侧芽数、上部一级分枝数、总侧芽数/叶节数呈极显著或显著负相关。
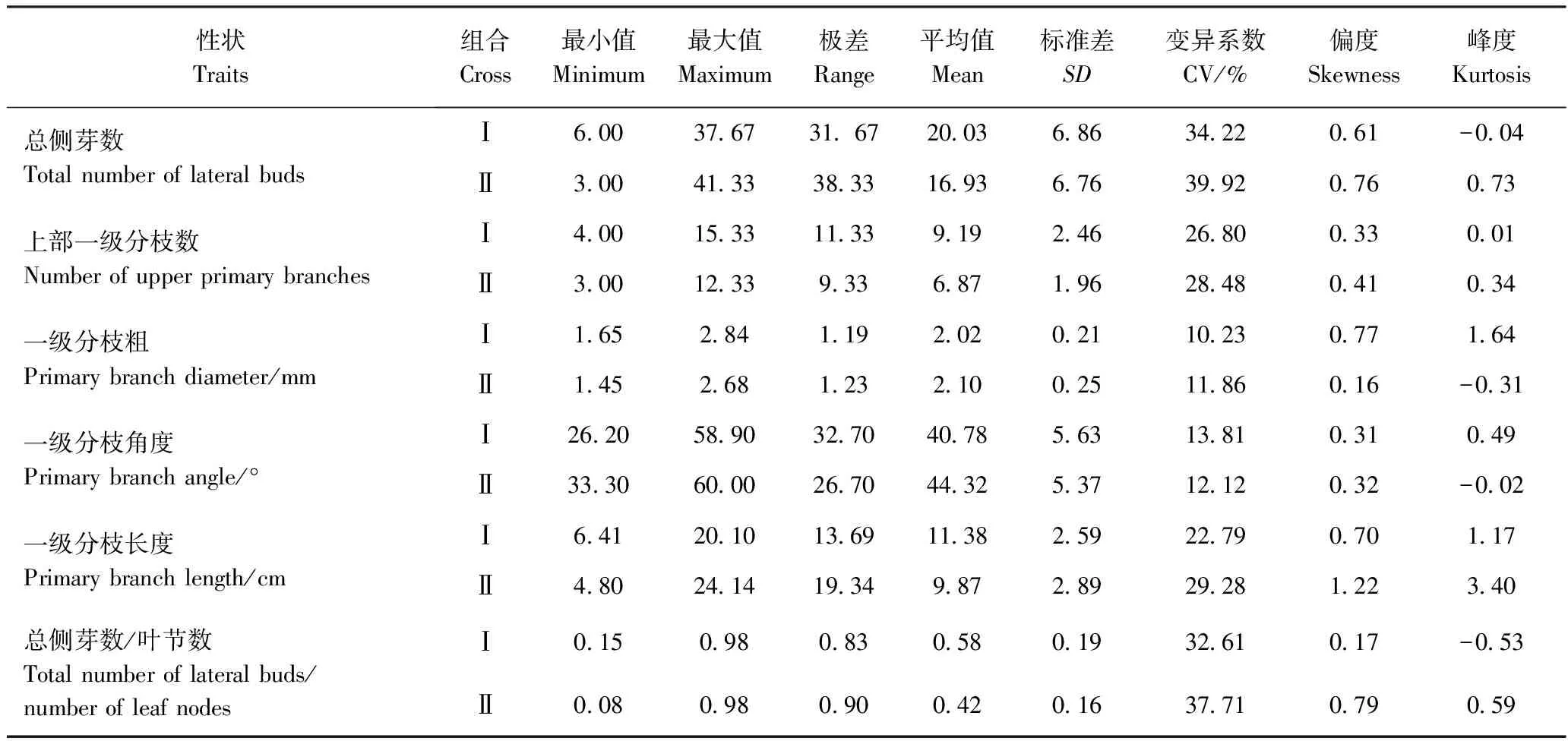
表1 F1群体分枝性状的表型特征值Table 1 Phenotypic characteristics of branching traits in segregating F1 populations
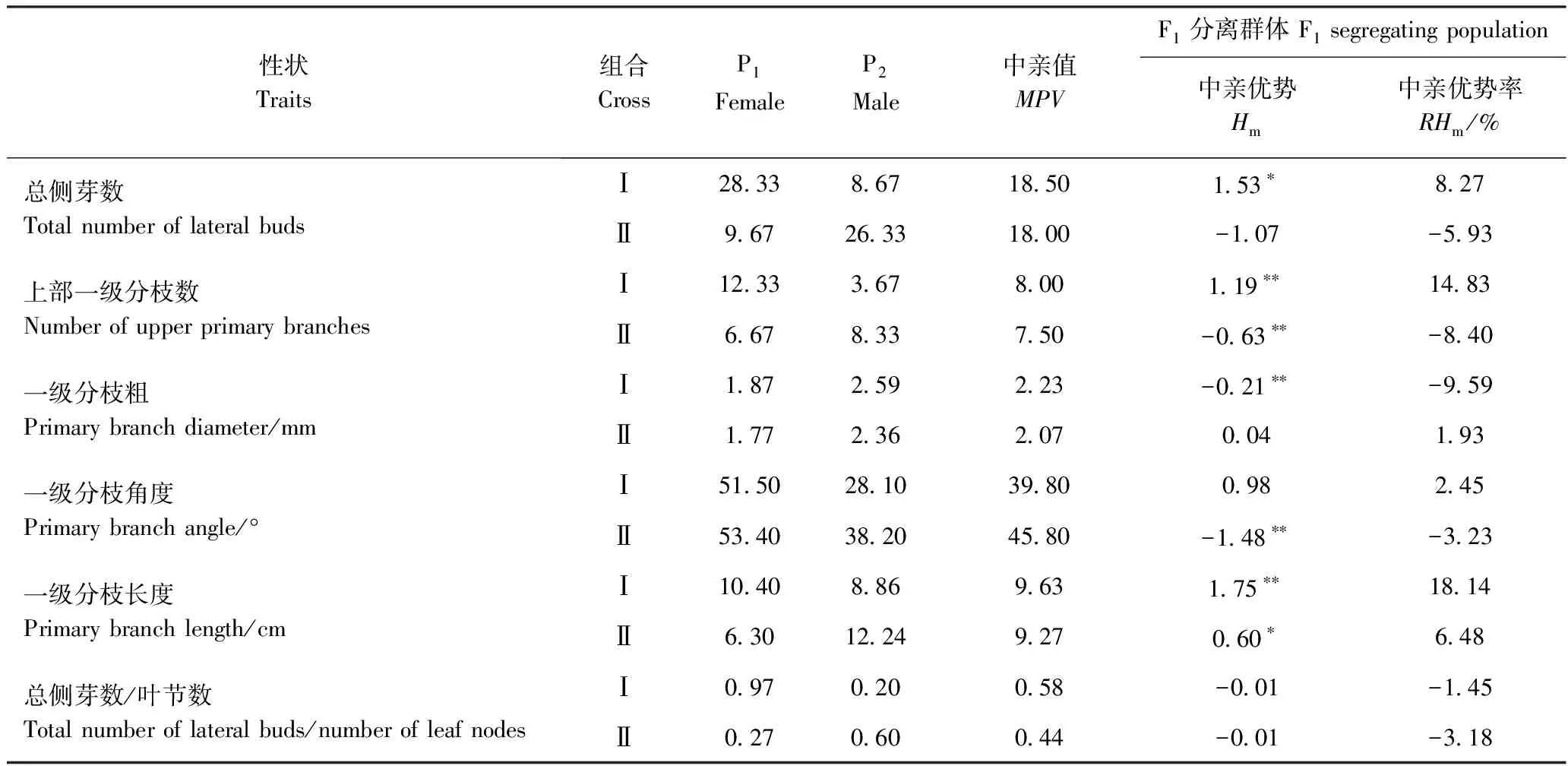
表2 F1群体分枝性状的杂种优势表现Table 2 Heterosis of branching traits in segregating F1 populations
2.3 标准切花菊分枝性状的最适遗传模型与适合性分析

表3 6个分枝性状的Pearson相关性Table 3 Pearson correlation coefficients between six branching traits
由各分枝性状的最适模型及检验结果(表5)可知,在两组合中,一级分枝粗、一级分枝角度都符合A-0模型,无主基因控制,属于微效多基因控制性状;总侧芽数符合A-1模型,由一对表现为加-显效应的主基因控制。上部一级分枝数、一级分枝长度和总侧芽数/叶节数在两组合中差异较大:上部一级分枝数在组合Ⅰ中符合A-0模型,而在组合Ⅱ中符合B-1模型,由两对表现为加-显-上效应的主基因控制;一级分枝长度在组合Ⅰ中符合A-0模型,而在组合Ⅱ中符合A-1模型,由一对表现为加-显效应的主基因控制;总侧芽数/叶节数在组合Ⅰ中符合B-3模型,由两对表现为加性效应的主基因控制,而在组合Ⅱ中符合A-1模型,由一对表现为加-显效应的主基因控制。
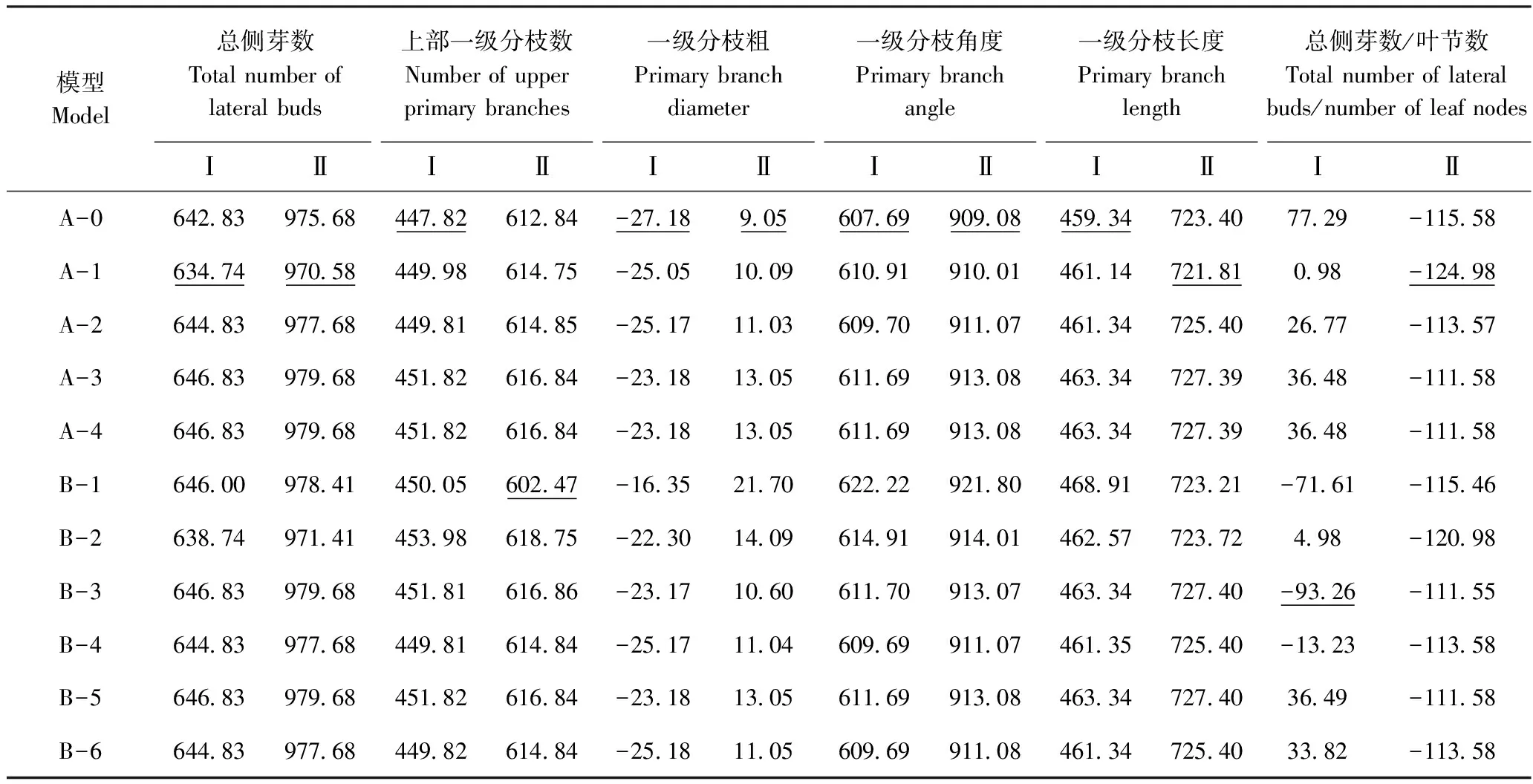
表4 两杂交组合分枝性状分离分析的AIC值Table 4 The AIC values of various genetic models for branching traits of two cross combinations
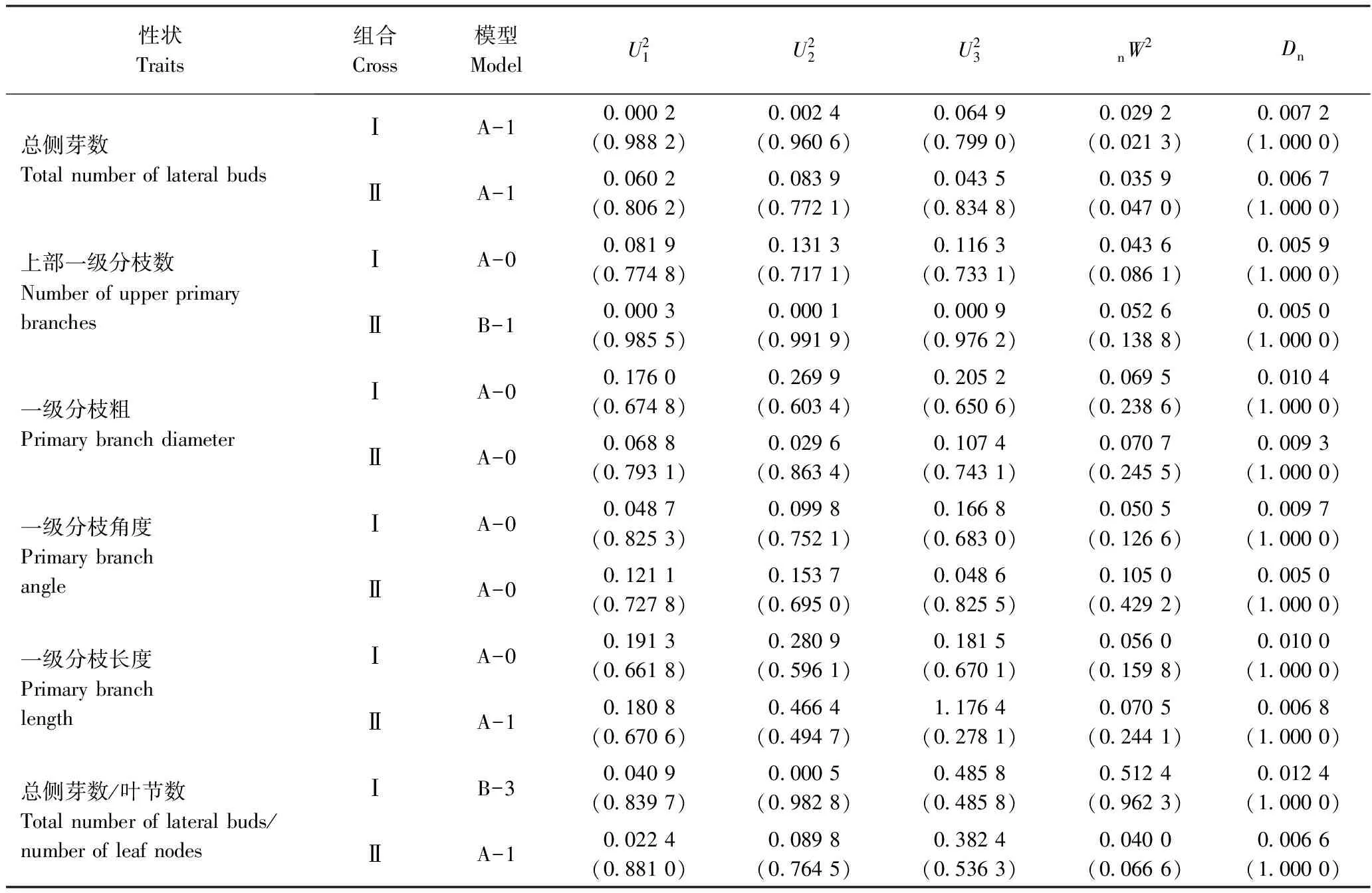
表5 两杂交组合分枝性状入选模型的适合性检验Table 5 Test for goodness-of-fit of selected genetic model for branching traits of two cross combinations
2.4 标准切花菊分枝性状的遗传参数估计
由表6可知,在组合Ⅰ中,控制总侧芽数的一对主基因的加性效应为6.193,显性效应为-6.096,表现为负向部分显性,主基因遗传率为57.39%;控制总侧芽数/叶节数的两对主基因的加性效应分别为0.127和0.457,主基因遗传率为98.84%。在组合Ⅱ中,控制总侧芽数的一对主基因的加性效应为5.494,显性效应为-5.484,表现为负向部分显性,主基因遗传率为46.73%;控制上部一级分枝数的两对主基因的加性效应分别为2.810和1.146,显性效应为-1.398和-0.305, 均表现为负向部分显性,两对主基因之间的加-加上位效应、加-显上位效应、显-加上位效应和显-显上位效应分别为0.320、-1.130、1.190和1.467,主基因遗传率为93.04%;控制一级分枝长度的一对主基因的加性效应为2.082,显性效应为-2.082,表现为负向部分显性,主基因遗传率为36.84%;控制总侧芽数/叶节数的一对主基因的加性效应为0.140,显性效应为-0.140,表现为负向部分显性,主基因遗传率为52.11%。
2.5 优选F1株系的获得
对两个杂交组合的244个F1株系利用总侧芽数和上部一级分枝数进行初步筛选,选出10株总侧芽数少于10且上部一级分枝数少于5的优选株系(表7)。可通过分枝性状稳定性鉴定、复选成为分枝较少的标准切花菊新品种,或作为“桥梁材料”进行回交,进一步研究分枝性状的遗传特性。
3 讨论
杂种优势利用是观赏植物杂交育种的有效手段[20-22]。在栽培菊花中,遗传背景复杂,基因组高度杂合,后代变异分离情况也较为复杂[23-24]。菊花的花器[25]、绿心[18]、侧芽数和侧枝数[11, 13-14]等性状表现出负向中亲优势,杂种优势呈下降趋势;在菊花的花色[26-27]、耐寒性[28]等性状的遗传中还存在偏母性遗传。在实际杂交育种过程中,应合理选配父母本,结合具体的F1分离情况综合判断,充分利用杂种优势。在本研究中,总侧芽数和上部一级分枝数在组合Ⅰ中表现为正向中亲优势,而在组合Ⅱ中表现为负向中亲优势,这种差异可能是杂交的父母本遗传背景不同导致的,但总侧芽数和上部一级分枝数在两组合中均表现为偏母性遗传。因此,以培育少侧芽侧枝标准切花菊品种为杂交目标,需要选择总侧芽数、上部一级分枝数少的品种作为母本。

表6 标准切花菊4个分枝性状的遗传参数估计值Table 6 Estimation of genetic parameters for four branching traits of standard cut-chrysanthemum
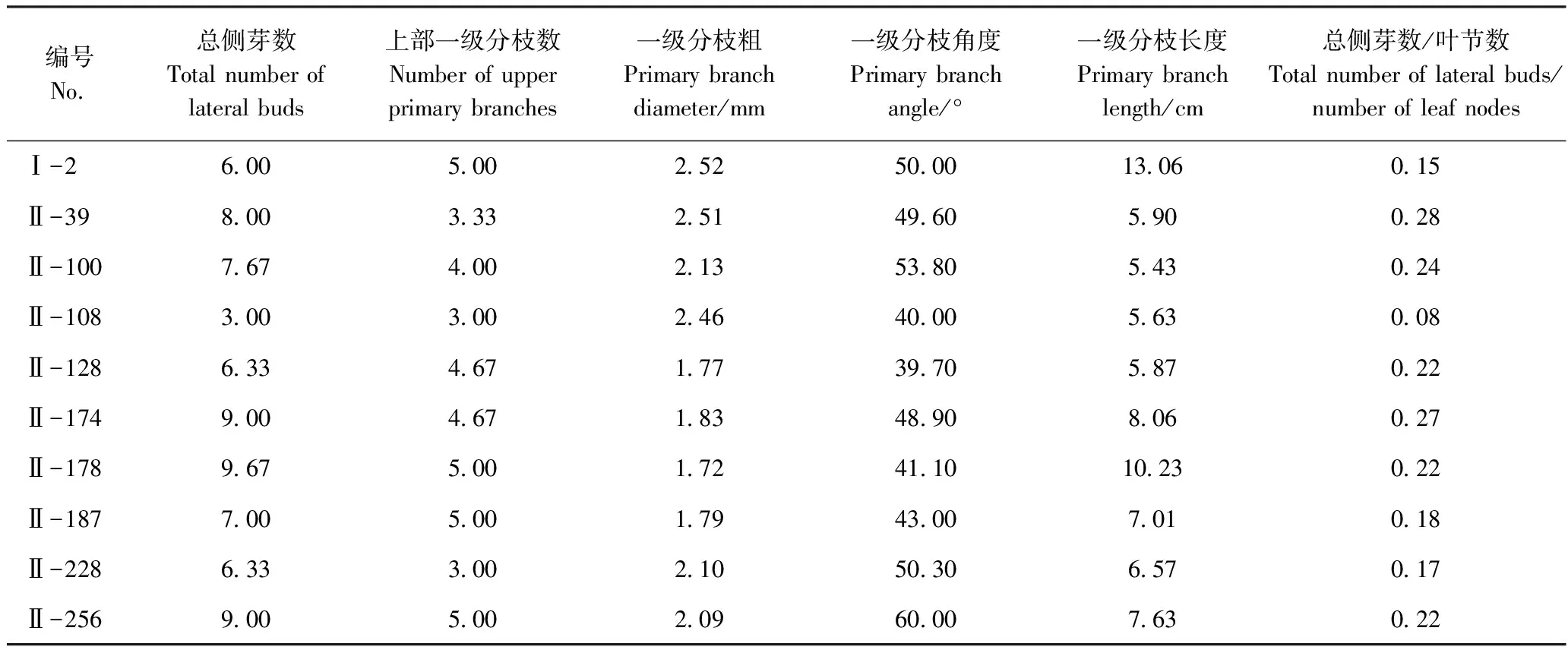
表7 优选F1株系的分枝性状Table 7 Branching traits of screened F1 strains
主基因及其效应检测是植物数量性状遗传研究的重要内容,可为后续遗传改良奠定重要基础[29-33]。在本研究中,总侧芽数在两组合中均检测到一对表现为加-显效应的主基因,上部一级分枝数和一级分枝长度在组合Ⅱ中分别检测到两对表现为加-显-上效应的主基因和一对表现为加-显效应的主基因,总侧芽数/叶节数在组合Ⅰ和组合Ⅱ中分别检测到两对表现为加性效应的主基因和一对表现为加-显效应的主基因。上部一级分枝数、一级分枝长度、总侧芽数/叶节数在不同的群体中表现出不同的主基因效应,这可能是由于在不同遗传群体中杂交亲本遗传背景不同造成的,在切花小菊[13]、甘蓝型油菜[34]、玉米[35]等植物的分枝性状遗传中也存在这种现象。其中,总侧芽数/叶节数主基因遗传率为52.11%~98.84%,遗传率较高,受环境影响较小,表现较为稳定,可在早期选择中适当提高选择标准[36]。在组合Ⅱ中发现,总侧芽数、上部一级分枝数、总侧芽数/叶节数的负向中亲优势主要来源于主基因的显性效应,可通过轮回选择和修饰回交聚集增效基因来改良[37]。这些主基因效应的发现将有助于进一步了解标准切花菊分枝性状的遗传基础,对标准切花菊分枝性状的QTL定位研究和通过分子育种缩短育种周期具有重要意义。
性状间的相关性在植物育种中有重要意义。在育种选择过程中,可以根据性状间的相关性,利用测量相对简便的性状进行选择,提高选种效率[38]。对于标准切花菊而言,总侧芽数、上部一级分枝数和总侧芽数/叶节数对其生产成本与成花品质尤为重要。本研究分析各分枝性状的相关性发现,总侧芽数、上部一级分枝数、一级分枝长度和总侧芽数/叶节数两两之间呈极显著正相关。因此,本研究通过总侧芽数、上部一级分枝数对F1株系进行初步筛选,得到10株优选株系,以培育侧芽侧枝较少的标准切花菊新品种,或作为中间材料进行下一步的分枝遗传特性研究。
4 结论
本研究明确了两个标准切花菊杂交组合中分枝性状的杂种优势,发现总侧芽数与上部一级分枝数在两个组合中均存在偏母性遗传效应,并在总侧芽数、上部一级分枝数、一级分枝长度、总侧芽数/叶节数4个分枝性状中检测到主基因效应,为少侧芽侧枝标准切花菊育种的亲本选配及后续QTL定位研究提供参考依据。同时,筛选获得优良的少侧芽F1株系可进一步鉴定、复选成为标准切花菊新品种,或作为育种中间材料继续进行分枝性状的遗传和育种研究。
