利用重组自交系高密度遗传图谱定位水稻芽期耐冷QTL
2020-11-18刘睿琦邹安东吴光亮王鹏李才敬张翔宇程琴王燕宁黄诗颖贺浩华边建民
刘睿琦 ,邹安东,吴光亮,2,王鹏,2,李才敬,2,张翔宇,2,程琴,2,王燕宁,2,黄诗颖,2,贺浩华,2,3,边建民,2,3
(1江西农业大学/作物生理生态与遗传育种教育部重点实验室,南昌 330045;2江西省水稻高水平工程研究中心,南昌 330045;3江西农业大学/作物生理生态与遗传育种江西省重点实验室,南昌 330045)
0 引言
水稻是世界重要的粮食作物之一,水稻高产与稳产对于保障世界粮食安全具有重要作用[1]。水稻起源于热带和亚热带地区,是一种喜温作物,对低温较为敏感。低温冷害是水稻生长的主要限制因子之一,是世界上水稻主产区普遍面临的问题。长期低温胁迫会导致水稻出现死苗、减产及品质下降等现象,对水稻安全生产造成了严重威胁[2-3]。根据冷害发生的时期,可将水稻冷害主要划分为芽期冷害、苗期冷害、孕穗期冷害、开花期冷害和灌浆期冷害[4]。其中,芽期冷害是指水稻从播种至第一片完全叶期间受到低温影响,导致出芽时间延长或烂种,严重影响产量。这个时期又可分为发芽期和芽期,发芽期耐冷性是水稻种子在低温下的萌发能力,而芽期耐冷性是指水稻种子萌发后,在低温下长成绿苗的能力[5]。芽期耐冷性是水稻生长早期重要抗逆指标之一,水稻芽期耐冷性强弱,不仅决定着水稻能否在生长前期早生快发,而且对水稻后期生长起着较为关键的作用[6-7]。因此,寻找水稻芽期耐冷性QTL,对于解析水稻耐冷分子遗传基础、培育耐冷水稻品种具有重要作用。
近年来,随着分子生物学技术发展,水稻耐冷性QTL 定位研究逐渐增多。利用不同水稻遗传群体和不同耐冷性鉴定方法,定位了多个耐冷相关的QTL。其中,以种子发芽率或成苗率作为鉴定指标,定位了至少99 个芽期耐冷性QTL[8]。Miura 等[9]利用‘Nipponbare’/‘Kasalath’回交构建的回交自交系(BIL)群体,检测到5个芽期耐低温QTL,贡献率均达到10%以上。Zhang等[10]和陈玮等[11]利用‘Lemont’/‘特青’构建的RILs群体,通过卷纸法将芽期耐冷的QTL定位在第1、3、7、11 染色体上,其中LOD 值最大的QTL 位于11号染色体,命名为qSCT-11,可用于水稻芽期耐冷性遗传改良。Baruah 等[12]利用RILs 群体把粳稻‘Hokkaido’芽期耐冷性QTL定位在第1、5、11和12等染色体。张露霞等[13]以‘Asominori’/‘IR24’构建的RILs群体和染色体片段置换系为材料,以芽期死苗率为耐冷性评价指标,检测到3个芽期耐冷性QTL,分别命名为qCTBP-5-1、qCTBP-5-2和qCTBP-12,且芽期耐冷等位基因均来自于粳型耐冷亲本‘Asominori’。朱金燕等[14]以‘陆矮4 号’为受体亲本、‘日本晴’供体亲本的单片段代换系(SSSLs)为材料,共检测到8个芽期耐冷QTL,分布在水稻第1、6、8、9 和10 号染色体。Zhao等[15]以‘东乡野生稻’为供体构建的159个片段代换系(CSSLs)为材料,检测到7个耐冷性QTL,并鉴定1株极耐寒株系‘SIL157’。随着研究的深入,QTL间的上位性研究逐渐增多,研究表明上位性效应在水稻耐冷遗传中也起着重要作用[16]。杨洛淼等[17]利用以粳稻品种‘东农422’为母本,耐冷品种‘空育131’为父本,衍生构建的190个F6和F7株系为试验材料,对水稻发芽期、芽期、苗期和孕穗期进行了耐冷鉴定,共检测到33对上位性QTL,其中12对上位性效应为负,22对表现为正,贡献率介于0.41%~13.54%之间,其中有11 对有加性QTL 参与互作。曲婷婷等[18]以水稻籼粳交(‘圭630’/‘02428’)构建的加倍单倍体(DH)群体为材料,检测到8 个控制水稻苗期耐冷性的上位性互作位点,分布在水稻第2、7、8、9和11染色体。Zhang等[10]利用籼粳交(‘Lemont’/‘特青’)构建的包含269 个家系的RILs群体为材料,检测到9个上位性互作位点,每个上位性位点对水稻幼苗耐冷性的影响都很小,但所有互作位点绝对上位效应总和达36%,说明上位效应在幼苗耐冷性遗传控制中的重要性。
然而,水稻耐冷性是受多基因控制的复杂数量性状,传统水稻芽期耐冷性QTL定位多采用籼粳交后代群体,定位的芽期耐冷QTL等位基因较多来自粳型品种。由于遗传分化差异,粳稻耐冷QTL很难在短期内通过分子标记技术转育到籼稻中,来提高籼稻耐冷性,在很大程度上限制了籼稻耐冷性分子育种。因此,挖掘籼稻中耐冷QTL(基因),对于利用分子标记技术改良籼稻耐冷性有重要作用。基于此,本研究利用双季稻区种植较为广泛的超级稻品种‘五丰优T025’双亲‘五丰B’和‘昌恢T025’为材料,构建了包含124 个家系的RILs 群体,并对水稻芽期耐冷性进行QTL 分析,以期挖掘其中芽期耐冷QTL,为籼稻特别是双季稻籼稻芽期耐冷性育种提供参考。
1 材料和方法
1.1 试验材料
‘五丰优T025’是长江中下游种植比较广泛的超级杂交晚稻品种[19]。本研究以‘五丰优T025’的双亲‘五丰B’和‘昌恢T025’为材料,构建了包含124 个家系的RILs群体。
1.2 基因分型和遗传连锁图谱的构建
提取‘五丰B’、‘昌恢T025’和124 个家系的基因组DNA,利用物理法断裂DNA 并构建~300 bp 的文库;利用华大基因MGISEQ-2000设备对构建的文库进行测序;基于测序结果对其中表现多态性标记的SNP进行筛选;将连续多个SNP标记且具有相同的基因型概括为一个bin 标记[20];最后利用获得的bin 标记结合JoinMap 4.2 软件构建了‘五丰B’和‘昌恢T025’杂交衍生的RILs群体高密度连锁图谱。
1.3 芽期耐冷性鉴定
从亲本和RILs 群体每个家系中各挑选籽粒饱满完整且无病虫害的种子40粒,置于垫有滤纸的培养皿中,加少量水于25℃下浸种1~2天,然后挑选长势基本相同的露白种子30 粒,加少量水,放入10℃低温恒温箱进行低温处理7 天。低温处理结束后,从低温恒温箱取出,将材料放入25℃的人工气候室恢复生长7天。恢复后第7 天后测量根数、根长和芽长及幼芽存活率。恢复期间,每天换水保证水分充足。实验重复3次,求其平均数进行分析。
1.4 QTL分析
采用IciMapping V4.2 软件对RILs 群体进行数据分析和QTL定位,LOD的阈值设为2.5,当测定LOD值大于2.5时,则判定该区段存在一个QTL,并同时测定贡献率与加性效应。加性效应正值表明耐冷性的等位基因来自于亲本‘五丰B’,负值则表示来源于亲本‘昌恢T025’;LOD的阈值为5.0 判断是否有互作效应,当测定LOD值大于5.0时,则判定存在互作效应,并同时测定贡献率与上位性效应。QTL 的命名遵循McCouch 等[21]提出的原则。用SPSS22 软件与Microsoft Excel 2017内置公式进行显著性分析。
2 结果和分析
2.1 遗传图谱构建
对测序的数据进行分析,获得高质量的SNP 701,310 个。对获得SNP 进行bin 转换,经过严格筛选,最终获得均匀分布在12条染色体上2578个bin标记,平均每条染色体包含214.8个bin标记。其中,标记最少的是8号染色体,有159个bin标记,标记最多的是1号染色体,有303个bin标记(表1)。进一步利用这些bin标记分析124个家系构建了高密度遗传图谱,该图谱总距离为1762.8 Mb(图1)。其中,2个相邻标记之间的平均距离为0.70 Mb,12 号染色体的平均距离最小,为0.58 Mb,8号染色体上的平均距离最大,为0.92 Mb。
2.2 芽期耐冷性的表型分析
如表2 所示,10℃低温处理后,‘五丰B’根长和根数显著大于‘昌恢T025’;‘五丰B’种子芽长大于‘昌恢T025’,差异不显著;‘昌恢T025’和‘五丰B’幼苗存活率相同,均为100%。表明在低温胁迫下,‘五丰B’长势要好于‘昌恢T025’。
10℃低温处理后,RILs 群体根长、芽长、根数和存活率均表现出不同程度差异(表2)。RILs群体偏度及峰度均小于1,表明RILs群体根长、芽长和根数表现为近似连续正态分布,并存在明显的超亲分离现象(表2,图2)。表明‘五丰B’和‘昌恢T025’衍生的RILs 群体在芽期10℃低温处理后,幼芽的根长、芽长和根数这3 个性状均表现出典型的数量性状遗传,符合QTL 定位要求。
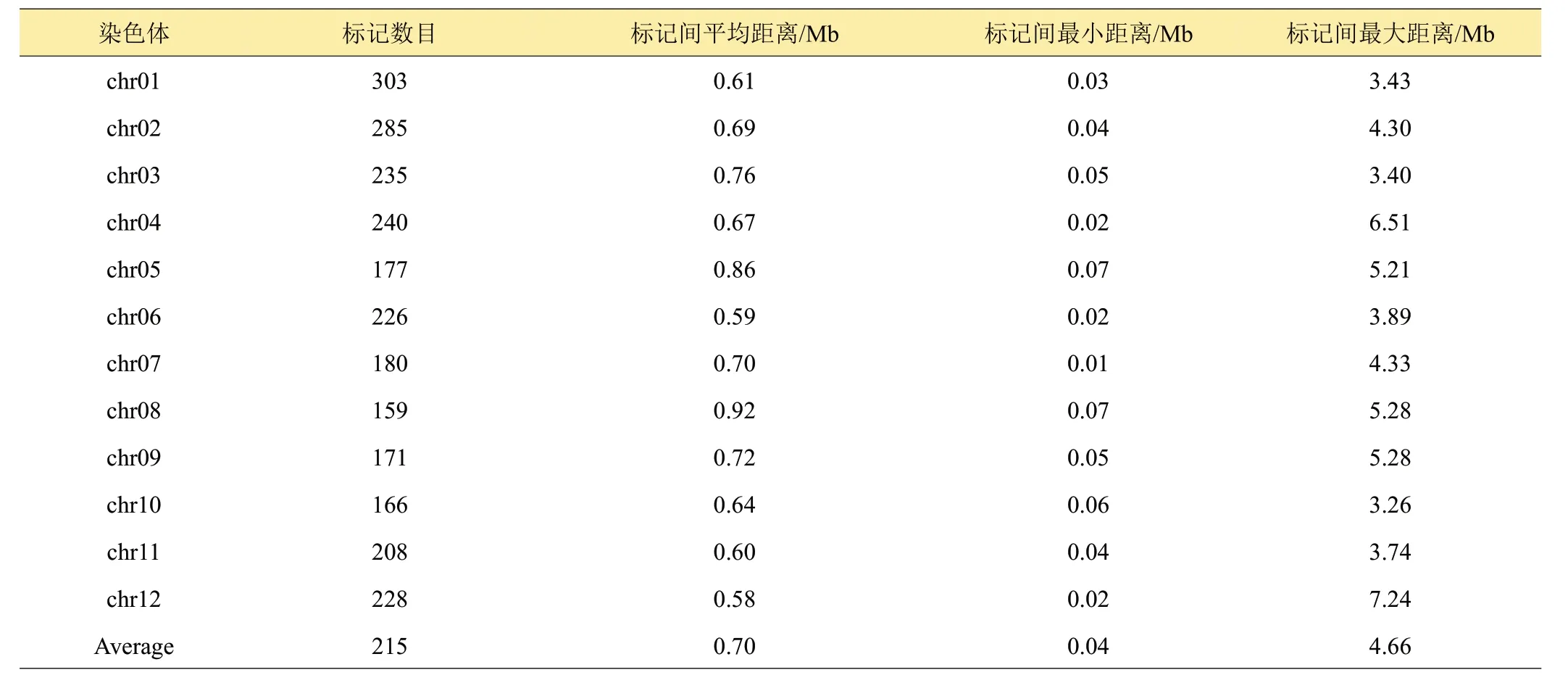
表1 染色体标记信息及遗传距离信息
2.3 相关性分析
利用SPSS 22软件对RILs群体芽期10℃冷处理后的根长、芽长、根数和存活率进行相关性分析。结果显示,根长和芽长在10℃低温处理后存在极显著的正相关,相关系数为0.39;芽长和根数在10℃低温处理后存在极显著的正相关,相关系数为0.23(表3)。表明该RILs 群体在10℃低温处理后,根长和芽长,芽长和根数之间存在联系。
2.4 水稻芽期耐冷性的加性QTL分析
采用QTL Icimapping v4.2 软件进行QTL 分析,共检测到3 个控制芽期耐冷性的QTL,别是qRL1、qRL2和qBL6。其中,qRL1、qRL2控制根长,qBL6控制芽长。这些QTL 分别位于水稻第1、第2 和第6 染色体,LOD值分别为2.98、2.51和5.26,可解释的表型变异率在8.67%~14.04%之间,3个QTL增效等位基因均来自于亲本‘昌恢T025’。本研究未检测到控制存活率和根数加性QTL(表4,图3)。
2.5 水稻芽期耐冷上位性QTL分析
本研究共检测出13 对水稻芽期耐冷上位性互作QTL(表5,图4)。其中以低温处理后水稻芽长作为耐冷鉴定指标,检测到第3染色体和第8染色体之间存在上位性互作,其LOD值为5.21,上位性效应为-0.24,贡献率为21.77%。以低温处理后水稻存活率作为耐冷
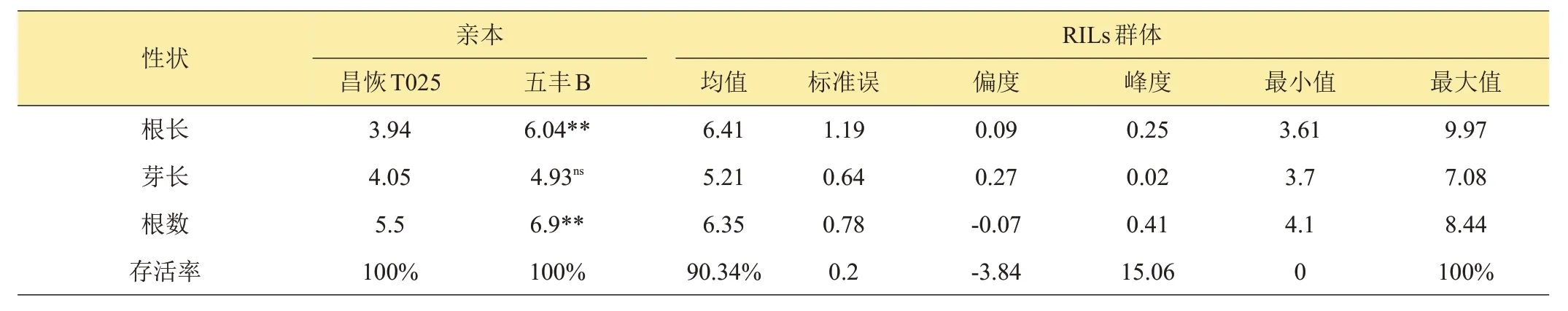
表2 芽期耐冷在亲本和RILs群体中的分布情况

表3 RILs群体芽期10℃低温处理后种子性状的相关性分析
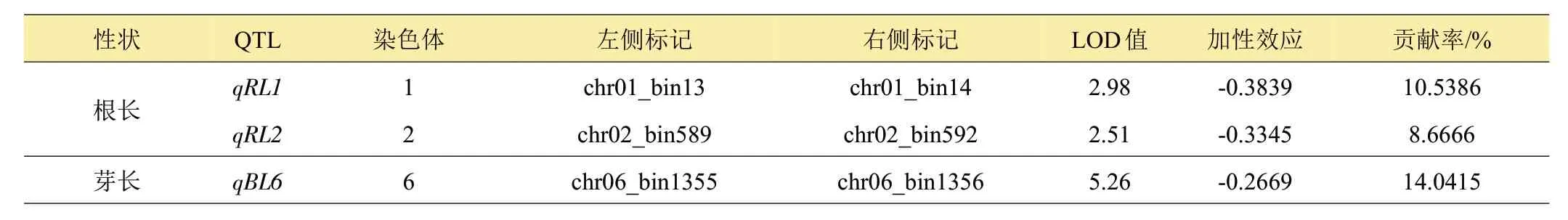
表4 水稻芽期耐冷性的QTL定位及遗传效应
鉴定指标,共检测到12 对上位性互作QTL,LOD 值在29.84~41.24 之间,上位性效应在0.23~0.26 之间,贡献率在4.68%~4.76%之间。以低温处理后水稻根长和根数作为水稻耐冷性鉴定指标,未检测到上位性互作QTL。

表5 水稻芽期耐冷性的上位性效应位点
3 讨论与结论
低温冷害是影响水稻生长发育的重要因素之一。水稻芽期冷害会造成死苗或烂秧,影响水稻生长和成熟,最终导致水稻产量亏损,所以提高水稻芽期耐冷性是水稻高产的必然要求。从国内外研究可知,水稻耐冷性是受多基因控制的复杂数量性状遗传,利用不同的研究群体定位了多个水稻耐冷相关的QTL[8]。但是,芽期耐冷相关QTL 定位的数量相对较少[22-25]。因此,定位更多的影响水稻芽期耐冷QTL对于解析水稻芽期耐冷遗传机理并进行水稻分子育种具有重要作用。
本实验共检测出3 个控制芽期耐冷性QTL,分别位于水稻第1、2和6染色体。通过比较发现,戴陆园[26]以‘昆明小白谷’与‘日本晴’构建NILs 群体定到的生殖期耐冷QTLqRCT6b与本研究定位的qBL6(物理位置:chr6:3.9478 Mb~4.0048 Mb)位置接近。目前暂未发现与qRL1(物理位置:chr1:0.6751 Mb~0.742 Mb)和qRL2(物理位置:chr2:18.1933~18.3025 Mb)相同或接近的水稻耐冷QTL。这2个QTL可能是水稻芽期耐冷新位点,需后续进一步深入探究,结果对于解析水稻芽期耐冷遗传机理并进行水稻芽期耐冷改良具有重要指导作用。此外,本研究未检测到控制存活率和根数加性QTL,很可能是由于控制两个性状的位点效应较小。
一般而言,粳稻品种芽期耐冷性普遍强于籼稻品种[27],通过籼粳交分离群体,也定位了来源于粳稻品种的耐冷QTL[8]。而关于籼稻间耐冷性差异研究较少,限制了笔者对不同籼稻品种耐冷性遗传基础的了解。本实验以2个不同籼稻品种‘五丰B’和‘昌恢T025’进行研究,发现籼稻之间也存在耐冷性的差异,同时检测到了3 个水稻芽期耐冷性QTL,其增效等位基因均来自冷敏型籼稻恢复系‘昌恢T025’。类似的研究结果在前人的实验中也有提及,如张露霞等[13]利用‘Asominori’/‘IR24’构建的RILs和染色体片段置换系为材料,以芽期死苗率为耐冷性评价指标,鉴定出2个来自籼稻品种‘IR24’对芽期耐冷性的增效等位基因。王棋[28]以‘泸恢99’和‘沈农265’杂交衍生的重组自交系为材料,以低温条件下水稻发芽率和苗期叶片赤枯度为耐冷性鉴定指标,检测到的qLTG-5和qSCT-9,其耐冷增效等位基因均来源于籼稻品种‘泸恢99’。这些结果表明籼稻中也蕴藏着耐冷基因(QTL),因此,挖掘籼稻耐冷基因(QTL)对于利用分子标记技术进行耐冷基因(QTL)聚合育种提高籼稻耐冷性具有重要作用。‘昌恢T025’作为江西等双季稻区比较有代表性的籼稻恢复系品种,可以利用‘昌恢T025’作为中间材料,进行连锁分子标记,通过分子标记辅助选择技术将这些芽期耐低温的主效QTL转入其他籼稻品种中,提高籼稻品种的芽期耐低温能力。此外,尽管本研究通过构建的高密度遗传图谱已将3个控制水稻芽期耐冷性的QTL定位在很小的染色体区间,但是由于重组自交系群体背景复杂,很难在短期内将其中耐冷QTL分离克隆,因此有必要构建近等基因系来分离并克隆这些水稻芽期耐冷性QTL,为解析籼稻耐冷遗传机理和后期利用这些QTL 进行分子标记辅助选择育种奠定基础。
上位性互作是数量性状遗传的重要组成部分,上位性互作对解析复杂性状的遗传基础具有十分重要的意义[29-32]。本实验以低温处理后的芽长为耐冷鉴定指标,检测到第3染色体与第8染色体之间存在上位性互作,其贡献值最大,为重组型大于亲本性互作。此外,每条染色体上都检测到一对低温胁迫下影响水稻芽期存活率上位性互作位点,并且位点相邻。而未检测到与根长和根数相关的上位性互作位点。表明在水稻芽期,上位性互作位点可能在调控水稻低温胁迫起了重要作用。关于芽期耐冷上位性效应研究涉及较少,所以发掘上位性互作新位点并分析位点间上位性互作和估算其效应,可以进一步了解芽期耐冷的遗传基础及为辅助育种提供依据。
本研究以双季超级稻品种‘五丰优T025’双亲‘五丰B’和‘昌恢T025’杂交衍生的重组自交系群体为材料,对水稻幼芽耐冷性进行了分析,结果显示水稻芽期耐冷性是由数量性状位点控制的。结合高密度连锁图谱,检测到3 个控制芽期耐冷性主效QTL,其中qBL6可能与已报道的基因是同一基因或等位基因,而qRL1和qRL2所在染色体位置目前尚未有耐冷QTL的报道,可能是控制芽期耐冷的新位点。这些耐冷QTL 定位对于后续克隆籼稻耐冷基因解析籼稻耐冷遗传机制奠定了基础。此外,本研究检测到13对影响水稻芽期耐冷上位性互作QTL,说明上位性互作效应在调控水稻芽期耐冷过程中起着重要作用。
