针对HBV preS1、preS2编码链的反基因锁核酸对Hep2.2.15细胞的抗病毒效果
2020-11-17许桂丹韦武均农顺强陈晓昊肖树荣邓益斌
彭 彬,许桂丹,韦武均,农顺强,陈晓昊,肖树荣,邓益斌
1.右江民族医学院附属医院检验科,广西百色 533000;2.广西肝胆疾病临床医学研究中心,广西白色 533000
乙型肝炎是由乙型肝炎病毒(HBV)感染引起的一种传染性疾病,易发展为肝硬化和肝细胞癌[1-2]。目前,全球HBV携带者约有3.5亿,而我国约1.3亿,其中,广西壮族自治区为HBV感染的重灾区之一,在临床上对于乙型肝炎的治疗仍缺乏特效药[3-4]。锁核酸亦称桥核酸,是近年来核酸研究领域的新热点[5],是一种环状核苷酸衍生物,其核糖的2′-O与4′-C缩水形成氧亚甲基桥、硫亚甲基桥或胺亚甲基桥,形似锁状或桥状。研究表明,与其他寡核苷酸相比,锁核酸具有更强大的优势,如热稳定性强[6-7]、抗酶解能力[8-9]、与DNA或RNA相似[10]、杰出的错配辨别力[11]、水溶性好[12]、降解半衰期长[13]等。本研究针对HBV preS1的3023-3037nt、3094-3109nt 2个位点和preS2 的3201-3215nt、3305-3320nt 2个位点分别设计合成锁核酸序列各1条,以Lipo3000介导转染HepG2.2.15细胞株,通过观察HBV的抑制作用,为研究新型抗HBV药物提供依据。
1 材料与方法
1.1材料 材料为HepG2.2.15细胞,其能够在细胞上清液稳定表达HBsAg、HBV DNA等物质,是HepG2细胞在染色体上整合了HBV基因组的肝肿瘤细胞株,购自上海通派生物细胞库,Lipo3000购自美国Invitrogen公司。
1.2仪器与试剂 荧光定量PCR检测HBV DNA试剂盒购自深圳亚能生物技术有限公司;HBV HBsAg定量检测试剂盒购自郑州安图生物工程股份有限公司;化学发光免疫测定仪购自郑州安图生物工程股份有限公司;实时定量PCR仪购自美国ABI公司。HepG2.2.15细胞常规培养于含G418、1%胎牛血清的高糖型DMEM培养基中,并于37 ℃、5% CO2条件下培养,通过荧光定量PCR和免疫化学发光技术测量HBsAg、HBV DNA等指标,观察锁核酸对HBV的抑制作用。
1.3方法
1.3.1锁核酸的设计与合成[4]从NCBI基因库中搜索到HBV ayw亚型全基因组序列(3.2 kb DNA,NC:003977.1,GI:21326584),利用RNA Structure 6.0软件针对preS1编码链的3023-3037nt、3094-3109nt和preS2编码链的3201-3215nt、3305-3320nt 4个位点分别设计1条长度不等的反基因锁核酸序列。3023-3037nt同聚嘌呤区:5′-CA*AA*TGCT*CCCGCT*C-3′(15 pb);3094-3109nt同聚嘌呤区:5′-T*GT*TGTCA*ATAT*GCC-3′(15 pb);3201-3215nt同聚嘌呤区:5′-A*GCA*GGAA*AATA*TAG-3′(15 pb);3305-3320nt同聚嘌呤区:5′-A*AGA*TTGA*CGA*TATG-3′(15 pb)。以上各序列中,标注*号的前一个碱基进行锁核酸修饰。各序列经BLAST排除与人同源后由上海生物工程有限公司合成修饰并纯化。
1.3.2实验分组与细胞转染 本实验设空白对照组与锁核酸-脂质体组。锁核酸-脂质体组包括4个亚组,分别为封闭3023-3037nt同聚嘌呤区(3023-3037nt组)、封闭3094-3109nt同聚嘌呤区(3094-3109nt组)、封闭3201-3215nt同聚嘌呤区(3201-3215nt组)、封闭3305-3320nt同聚嘌呤区(3305-3320nt组)。以1×105个/mL细胞浓度接种于6孔板中,每组随机各设6个复孔,待细胞贴壁达50%~60%后在各组每孔中加入锁核酸-脂质体药物,转染按照脂质体说明书操作,隔3 d收集培养上清液后再次给药,连续给药3次,并收集3、6、9 d的上清液。
1.3.3细胞上清液HBsAg的检测 采用化学发光免疫技术对细胞上清液的HBsAg重复检测3次,取平均检测值。
1.3.4HepG2.2.15细胞上清液HBV DNA的检测 取细胞培养上清液500 μL加入无酶EP管中,离心后留200 μL加入450 μL DNA提取液及4 μL内标溶液,100 ℃恒温处理15 min,按比例取相对应的PCR反应液及Taq酶,充分混匀之后按照每管30 μL分装到PCR八连管中,备用。往上述PCR反应管中加入处理过的待测标本核酸、阴性、强阳性、临界阳性质控品及不同浓度的定量参考品的上清液各20 μL,瞬时离心数秒后放置于荧光定量PCR仪中。扩增条件:92 ℃预变性2 min,93 ℃ 45 s,55 ℃ 60 s共 10个循环,93 ℃ 30 s,55 ℃ 45 s 共30个循环,72 ℃ 5 min共1个循环。
1.3.5细胞代谢活性的检测 采用CCK8比色法检测锁核酸对Hep2.2.15细胞代谢活性的影响,得到各组的吸光度(A)值。
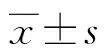

2 结 果
2.1反基因锁核酸对HBV DNA复制的抑制效果 锁核酸-脂质体各亚组加入锁核酸药物后显示,不同亚组的HBV DNA复制在3 d后开始受到抑制,且抑制效果随时间显著升高,锁核酸-脂质体各亚组与空白对照组比较,差异均有统计学意义(均P<0.05);在HBV preS1 的3023-3037nt、3094-3109nt 2个位点中,封闭3023-3037nt同聚嘌呤区的反基因锁核酸对HBV DNA复制的抑制效果最明显。在HBV preS2 的3201-3215nt、3305-3320nt 2个位点中,封闭3305-3320nt同聚嘌呤区的反基因锁核酸对HBV DNA复制的抑制效果最明显。见表1。
2.2反基因锁核酸对HBV DNA复制的抑制率 在HBV preS1 的3023-3037nt、3094-3109nt 2个位点中,封闭3023-3037nt同聚嘌呤区的反基因锁核酸对HBV DNA复制的抑制效果最明显。在preS2 的3201-3215nt、3305-3320nt 2个位点中,封闭3305-3320nt同聚嘌呤区的反基因锁核酸对HBV DNA复制的抑制效果最明显。见表2。

表1 反基因锁核酸对HBV DNA复制的抑制效果

表2 反基因锁核酸对HBV DNA复制的抑制率
2.3反基因锁核酸对HBsAg表达的抑制效果 锁核酸-脂质体各亚组加入锁核酸药物后显示,不同亚组的HBsAg抑制作用在3 d后开始受到抑制,且抑制效果随时间显著升高,锁核酸-脂质体各亚组与空白对照组比较,差异均有统计学意义(均P<0.05)。在HBV preS1的3023-3037nt、3094-3109nt 2个位点中,封闭3023-3037nt同聚嘌呤区的反基因锁核酸对HBsAg的抑制效果最明显。在HBV preS2 的3201-3215nt、3305-3320nt 2个位点中,封闭3305-3320nt同聚嘌呤区的反基因锁核酸对HBsAg的抑制效果最明显。见表3。
2.4反基因锁核酸对HBsAg表达的抑制率 在HBV preS1的 3023-3037nt、3094-3109nt 2个位点中,封闭3023-3037nt同聚嘌呤区的反基因锁核酸对HBsAg的抑制效果最明显。在HBV preS2 的3201-3215nt、3305-3320nt 2个位点中,封闭3305-3320nt同聚嘌呤区的反基因锁核酸对HBsAg的抑制效果最明显。见表4。
表4 反基因锁核酸对HBsAg表达的抑制率
2.5CCK8比色法检测锁核酸对Hep2.2.15细胞代谢的影响 用药9 d后,3023-3037nt组、3094-3109nt组、3201-3215nt组、3305-3320nt组的A值分别为2.209±0.006、2.211±0.007、2.208±0.009、2.210±0.011,空白对照组的A值为2.207±0.008,锁核酸-脂质体组各亚组与空白对照组比较,差异均无统计学意义(P>0.05)。
3 讨 论
HBV是由约3.2 kb碱基对组成的相对松弛不完全双链DNA环状分子,共有4个开放读码区,即S、C、P、X区,其中,S、C区序列是保守区,也是基因治疗的理想靶位[14-16]。在编码HBsAg的S基因5′末端有编码能力的核苷酸序列,称为preS区,可分为preS1区和preS2区,对应编码的是preS1蛋白和preS2蛋白。preS1蛋白位于表面抗原大分子蛋白的N末端,由109~119个氨基酸组成,preS2基因起始于第3 172位核苷酸,终止于第833位核苷酸。
本研究分别针对HBV preS1的3023-3037nt、3094-3109nt 2个位点和preS2 的3201-3215nt、3305-3320nt 2个位点设计锁核酸序列,通过Lipo3000脂质体转染入HepG2.2.15细胞内,通过检测空白对照组和锁核酸-脂质体组的HBV DNA复制和HBsAg表达等来评价不同位点锁核酸的疗效,实验结果显示,preS1的3023-3037nt位点每隔3 d连续3次对应给药之后,对HBsAg表达的3次抑制率分别为(36.17±3.26)%、(49.62±4.36)%、(61.94±4.11)%,对HBV-DNA复制的3次抑制率分别为(26.96±3.63)%、(38.31±4.03)%、(56.08±3.22)%;3094-3109nt对HBsAg表达的3次抑制率分别为(28.75±4.22)%、(37.07±4.31)%、(40.22±4.21)%,对HBV-DNA复制的3次抑制率分别为(22.16±4.22)%、(37.26±5.07)%、(40.24±4.54)%;3201-3215nt对HBsAg表达的3次抑制率分别为(43.28±4.36)%、(48.50±3.75)%、(68.14±5.34)%,对HBV-DNA复制的3次抑制率分别为(25.66±4.38)%、(27.14±3.24)%、(46.61±3.40)%;3305-3320nt对HBsAg表达的3次抑制率分别为(50.38±6.17)%、(57.42±4.19)%、(72.93±4.14)%;对HBV-DNA复制的3次抑制率分别为(48.68±3.42)%、(52.96±3.61)%、(61.79±3.40)%。由此可看出,preS1编码链的3023-3037nt的抑制效果较好,preS2编码链的3305-3320nt的抑制效果较好,而相对于preS1的3023-3037nt,preS2的3305-3320nt抑制效果较高,可更有效地通过HepG2.2.15细胞的核孔进入到细胞核内,与基因组cccDNA的编码链相结合并形成三链杂交分子,在细胞核的复制和转录水平发挥抗病毒效果,具体的机制还有待进一步的研究,此外,通过CCK8比色法证实锁核酸-脂质体各亚组对细胞的代谢活性无明显毒性作用。
综上所述,通过探究preS1 的3023-3037nt、3094-3109nt 2个位点和preS2 3201-3215nt、3305-3320nt 2个位点,采用实时定量PCR、化学发光免疫分析技术观察HBV DNA、HBsAg等抗病毒指标,这为preS1、preS2的靶位提供了理论和实验依据,也为HBV的药物治疗提供了有效的靶位。
