干旱绿洲区富士苹果树干边材茎流动态及其对环境因子的响应
2020-09-17马文涛程平李宏张志刚武胜利赵明玉
马文涛,程平,李宏,张志刚,武胜利,赵明玉
(1.新疆师范大学地理科学与旅游学院,乌鲁木齐830054;2.新疆林业科学院,乌鲁木齐830000;3.新疆阿克苏森林生态系统国家定位观测研究站,新疆 阿克苏843000)
阿克苏地区地处新疆环塔里木盆地绿洲带北缘,灌溉便利,日照充足,昼夜温差大,十分适合苹果(Malus domestica Borkh.)种植,是新疆乃至全国重要的优质苹果产区[1]。截至2017 年底,阿克苏地区苹果种植面积达2.13万hm2,产量为59.75万t,产值达32.86 亿元,分别占新疆苹果种植面积、产量、产值的38.7%、65.19%、62.12%,其中红富士苹果是阿克苏地区的地理标志保护产品,因其外观好、肉质细、酸甜适口而享誉疆内外[2],已成为促进区域经济发展、改善生态环境的特色产业。然而,该区是典型的旱作绿洲农耕区,降水量少,且农业用水与生产生活用水矛盾日益突出[3]。因此,水资源供需矛盾已成为该地林果业发展的主要限制因子,而适时适量的水分供给对于果树生长发育至关重要,是果树早实、高产、稳产、优质及长寿的重要条件[4]。为此,研究干旱绿洲区果树蒸腾耗水规律及其影响因子,对于了解果园水分承载力,指导果园科学灌溉具有重要意义。
植物蒸腾作用在植物生命过程中发挥着重要作用,其水分胁迫及需水状况可通过蒸腾速率反映[5-6]。有研究表明,树木边材中99%的茎流量用于植物叶片蒸腾耗水,且茎流速率与气象因子、土壤水分密切相关[7],其中:气象要素决定茎流的瞬间动态,土壤供水决定茎流的整体水平[8]。通过研究树干茎流变化规律及其与环境因子的相互关系,可有效地定量分析树木生长与蒸腾耗水的关系[9-10]。热扩散探针(thermal dissipation probe,TDP)法因具有连续、稳定、准确等优点,已被广泛应用于树木蒸腾耗水的研究[11-12]中。孙习轩对我国北方苹果树需水量与灌溉量的研究发现,不同地区、不同品种的苹果树蒸腾速率不同[13];在黄土干旱塬区[4,9-10,14]、山东丘陵区[15-17]、东北冷凉地区[18]进行的苹果树干茎流的研究也印证了这一结论。但目前对新疆绿洲区苹果树干茎流的研究较少,而且在已有的研究中侧重于探究不同天气条件下茎流变化及瞬时尺度下茎流速率对环境因子的响应模型,而对于生长期内苹果树整个夜间茎流的变化规律及在更大时间尺度上茎流变化是否与其他地区一致,则鲜有报道。基于此,本文以新疆南部阿克苏地区温宿绿洲为研究区域,对在该区域内广泛种植的富士苹果树树干茎流及环境因子进行连续动态监测,并分析不同尺度下树干茎流变化规律及其对环境因子的响应模型,实现对单木蒸腾耗水的预测,进而指导果园科学灌溉。
1 材料与方法
1.1 试验地概况
试验地位于阿克苏地区温宿县境内的新疆林业科学院佳木试验站,地理位置为41°15′N,80°31′E,海拔1 103 m,浅层地下水位9.3 m;属于温带大陆性荒漠气候区,年平均气温10~11 ℃,年降雨量60~80 mm,≥10 ℃年积温2 916~3 200 ℃,年日照时数2 780 h,无霜期190~215 d,春夏季特别干燥,秋冬季冷凉。果园土壤质地为砂壤土,0~80 cm土层平均容重为1.38 g/cm3,田间持水量为29.01%(以体积含水量表征),pH 为8.44,含有机质10.65 g/kg、有效磷22.54 mg/kg、速效氮50.21 mg/kg、速效钾367 mg/kg。
1.2 试验材料
试验地为8年生盛果期的富士(长富2号)苹果园,株行距4 m×5 m,东西向栽植,树形为疏散分层型;在果园中央选择3株生长健康、长势一致、无病虫害、无结疤的苹果树作为试验样株,平均地径(距离地面30 cm)为16.0 cm,平均树高为4.6 m,平均冠幅为4.1 m(东西)×4.7 m(南北)。灌溉方式为漫灌,灌溉量3 150 m3/hm2,年灌溉7 次,灌溉定额为2.205×104m3/hm2。采用基肥+追肥方式施肥:秋季施基肥油渣和羊粪,各0.5~1.0 kg/株;夏季追肥按m(磷酸二铵)∶m(尿素)=1∶1的比例混匀施用,约0.5 kg/株。施肥方法为沟施。定期对果园进行病虫害防治,并清除杂草。果园管理措施在该区域具有代表性。
1.3 试验方法
试验于2019年5—9月进行,采用热扩散探针法,通过检测插入树干边材的1对有热电偶的探针温差来计算茎流速率。在选定的3株苹果树干上各安装1个由美国Dynamax公司生产的FLGS-TDP插针式探针,型号为TDP-30,长30 mm,针头直径1.2 mm。茎流计工作电压用数据采集器CR1000(Campbell Scientific公司,美国)和PC400软件来调节和检测热电偶,系统设置为每30 min 记录一次数据,连续记录2个探针间的温差。试验前期调查发现,截断的3株地径16 cm的枯死苹果树的边材与心材具有明显的颜色差异,因此,用破坏性取样方法(打生长锥)测定其距离地面35 cm 处的样株边材厚度,从而计算边材面积。树干边材茎流速率(vsap)及茎流量(体积流量,Fs)由以下公式[11]计算:
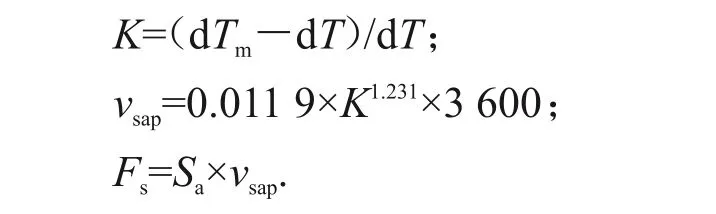
式中:K 为Grainer 定义的一个量纲一的量的“茎流指数”,用于消除茎流速率为0时的温差;dTm为无茎流时的温差最大值,℃;dT 为瞬时温差值,℃;vsap为茎流速率,cm/h;Fs为茎流量,g/h;Sa为边材面积,cm2。其中,dTm和dT为茎流仪系统自动检测结果。
环境因子测定:气象数据由HOBO自动气象站(Onset公司,美国)观测所得(架设高度2 m)。测定指标包括太阳辐射(solar radiation, Rs)、大气温度(air temperature,Ta)、空气相对湿度(relative humidity,RH)、风速(wind speed,Sw)等,每30 min记录一次数据。采用EM50数据采集器(Decagon公司,美国)测定土壤温度及其体积含水量,探头深度为10、30、50、70 cm,每天定时采集一次数据。水汽压差(water vapor pressure deficit,VPD)计算公式[10]为:
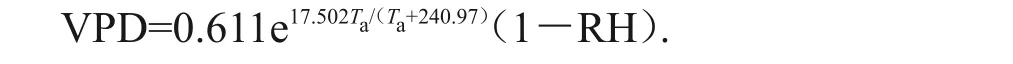
式中:Ta为大气温度,℃;RH 为空气相对湿度,%;e为常数,取值2.718。
1.4 数据处理
试验期间,由意外停电、更换损坏探针导致的茎流数据缺失,采用平均日变化法进行插补,并利用Excel 2013、SPSS 23.0 和Origin 2017 进行数据分析和作图。
2 结果与分析
2.1 富士苹果树干茎流变化特征
2.1.1 苹果树干茎流季节变化
选取在不同季节典型晴天的苹果树干茎流速率的测定结果,分析其在不同季节的茎流变化特征。结果(图1A~B)表明:夏季(6—8月)和秋季(9月),苹果树干茎流日变化主要呈双峰型曲线,在春季(5 月)以单峰型为主,偶有微弱的双峰型曲线。受环境因子影响,反映树木蒸腾耗水特性的树干茎流的启动时间、峰值、持续时间、进入低谷时间等时空变化特征[19],在不同季节间差异很大。如图1、表1所示:从茎流启动时间来看,夏季7月茎流启动时间为09:00—09:30,比春季5月茎流启动时间早0.5 h,比秋季9 月早1.5 h。从到达茎流峰值来看,秋季茎流到达峰值的时间比较集中,时间为13:00—13:30,峰值在17.78~24.15 cm/h 之间,而春季和夏季茎流到达峰值的时间比较分散,峰值分别在16.28~18.84 和26.93~29.67 cm/h 之间。从茎流持续时间来看,春、秋季的持续时间分别为10和9.5 h,夏季持续时间最长(12.5 h),比春季多2.5 h,比秋季多3 h。从茎流速率来看,夏季日变化格型宽、幅度大,茎流活跃,且7 月日均茎流速率最大,为10.78 cm/h,春季5月最小,为7.95 cm/h。
2.1.2 苹果树干茎流夜间变化
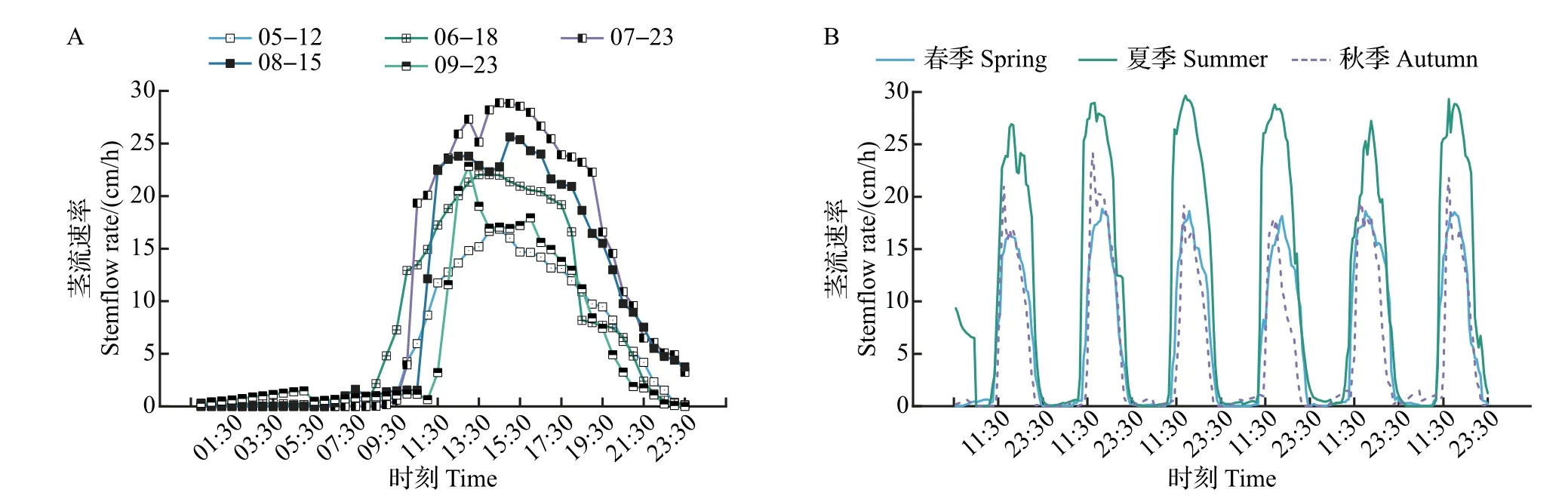
图1 富士苹果树干茎流速率不同月份日变化与不同季节连日变化Fig.1 Diurnal variation and daily changes of stemflow rate of red Fuji apple in different months and seasons
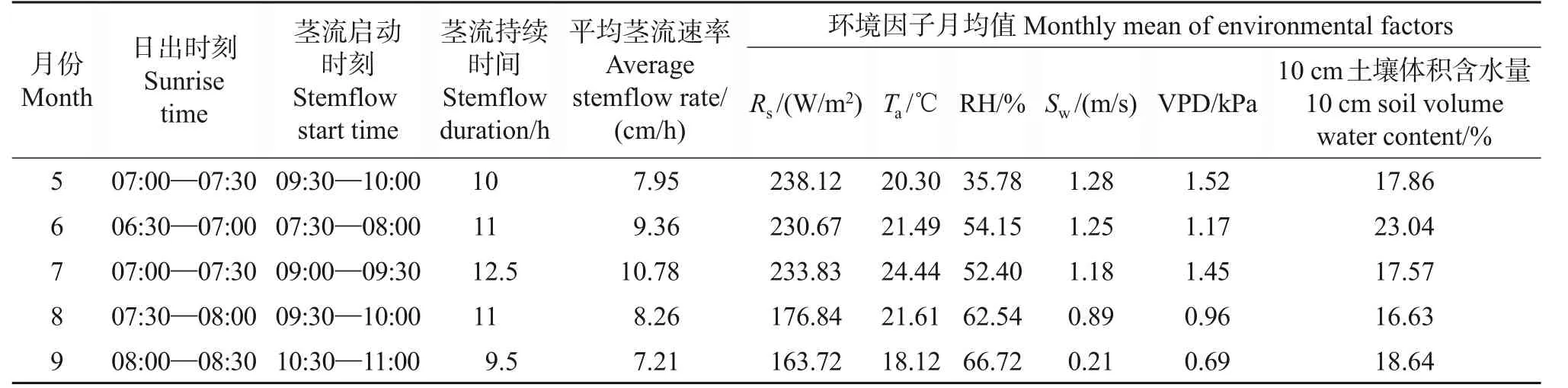
表1 富士苹果树干茎流速率月际动态变化Table 1 Monthly dynamic changes of stemflow rate of red Fuji apple
试验将一天中太阳辐射值小于或等于0时的树干茎流定义为夜间茎流。不同生长期昼夜茎流时间划分见表2。通过对树干茎流的监测,发现夜间茎流速率变化稳定,接近或等于0,其累积茎流量在单日茎流总量中占有一定的比例。图2 显示:生长期内典型晴天日温度在日落后到次日日出前呈现缓慢递减的趋势;空气相对湿度缓慢递增,并在日出前后达到最大;茎流速率变化趋势与夜间大气温度变化基本一致,与空气相对湿度变化趋势相反,且在日落后到次日00:30 茎流变化幅度大,在00:30到日出前变化较稳定,其值趋于0。说明树干茎流在日落后并未完全停止,而是处在缓慢下降的过程中,此时,树干仍以较低的茎流水平保持吸水状态。
将富士苹果树在不同生长期的昼夜茎流量分别进行累积统计,以了解昼夜间茎流的分布特点。结果(表3)表明:单日茎流量大小为7月>8月>6月>5月>9月,即7月茎流量最大(52.50 kg/d),9月最小(28.79 kg/d),前者单日茎流量约为后者的1.82 倍。夜间茎流量表现为9月>5月>8月>7月>6月,即9 月夜间茎流量最大,为2.96 kg/d,占比为10.28%。说明夜间茎流在果树单日茎流中占有一定的比例,在计算果树蒸腾耗水时不能忽略夜间茎流量。
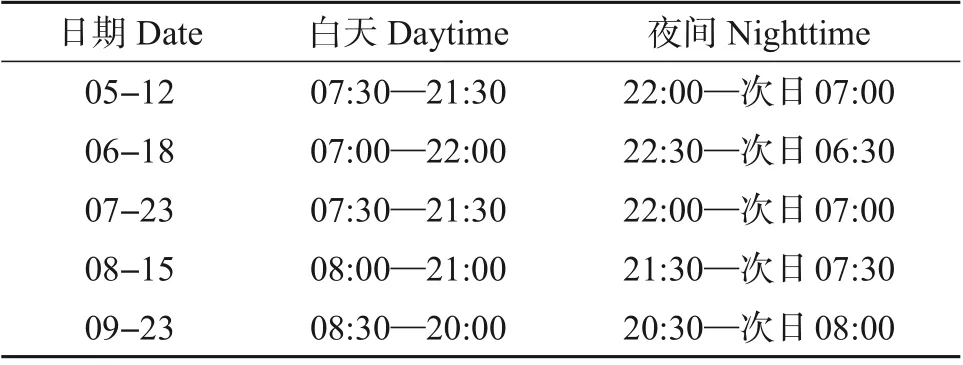
表2 昼夜茎流时间划分表Table 2 Daytime and nighttime allocation table of stemflow
2.2 不同季节富士苹果树干茎流速率变化与气象因子的相关性
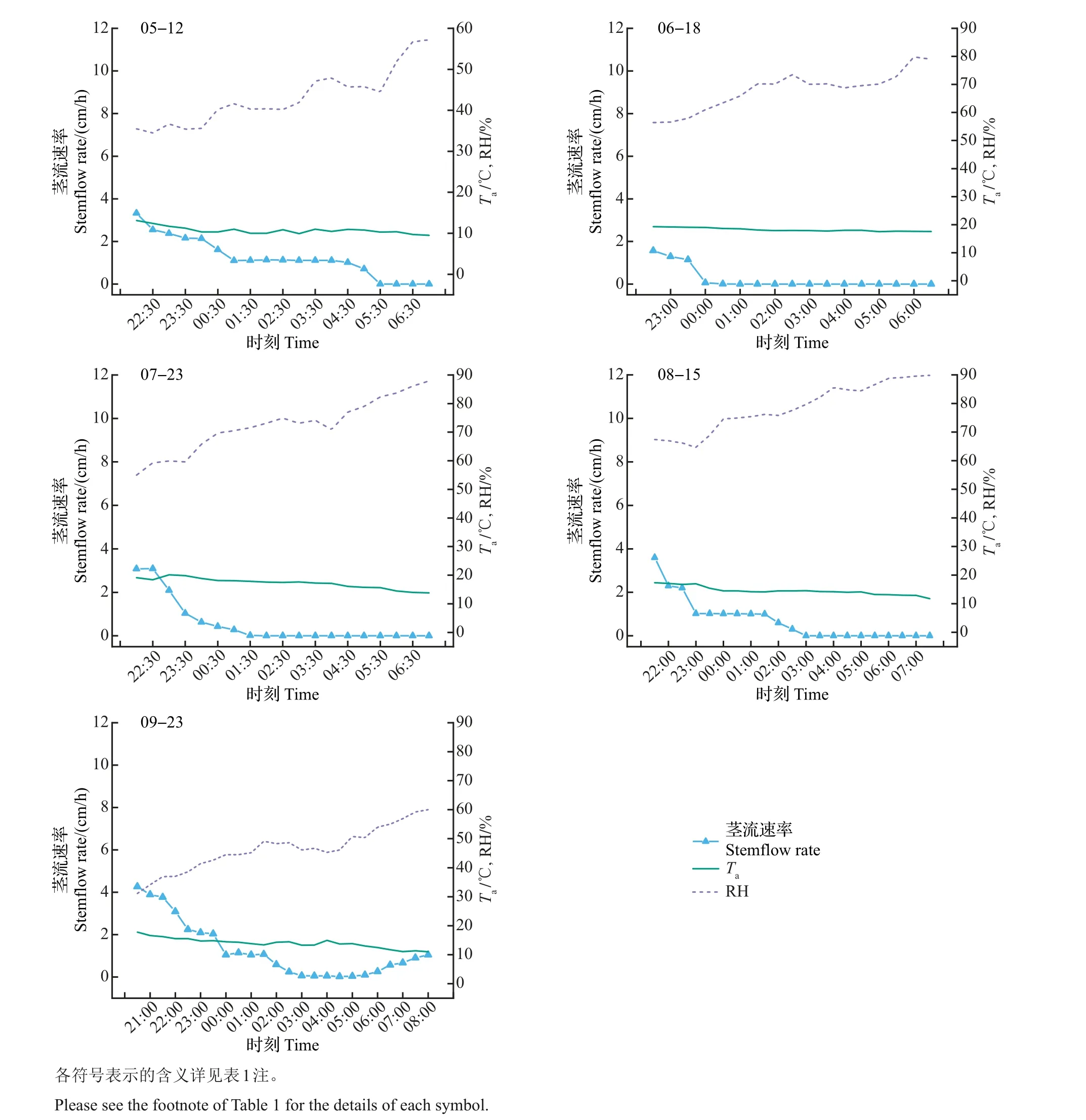
图2 生长期内夜间茎流速率与主要环境因子变化Fig.2 Variation of stemflow rate and main environmental factors at nighttime during the growth period of red Fuji apple
由于试验地地下水埋深较深,土壤含水量在瞬时尺度上变化较小且季节变化不显著,因此,树干茎流速率日变化主要与气象因子有关。基于5月、7月、9月内连续6个晴天的茎流速率与气象数据值进行相关分析,结果(图3、表4)表明:生长期内茎流速率与太阳辐射、大气温度、水汽压差、风速具有相同的变化趋势,均表现为显著正相关,与空气相对湿度的变化曲线相反,表现为显著负相关。春季影响茎流速率日变化的主要因子是水汽压差、太阳辐射、大气温度、风速,空气相对湿度的影响最小;影响夏季茎流速率日变化的主要因子是太阳辐射和大气温度,空气相对湿度影响最小;秋季对茎流速率影响较大的是太阳辐射、大气温度、空气相对湿度、水汽压差,风速影响最小,通径系数仅为0.008。可见,太阳辐射、大气温度、水汽压差是影响树干瞬时尺度上茎流速率的关键因子。
2.3 富士苹果树干日均茎流速率与环境因子的相关性
更大时间尺度上树木的茎流变化受众多因子的影响,如树体生理结构和功能,气象和土壤环境因子等。在以日为单元的时间尺度上,土壤温度和土壤含水量对树干茎流速率的影响较小,但在季节尺度上土壤温度和土壤含水量与日均茎流速率显著相关[20]。因此,在较大时间尺度上,应考虑土壤温度、土壤含水量对树木蒸腾量的影响。

表3 昼夜树干茎流量占比Table 3 Daytime and nighttime stemflow ratio
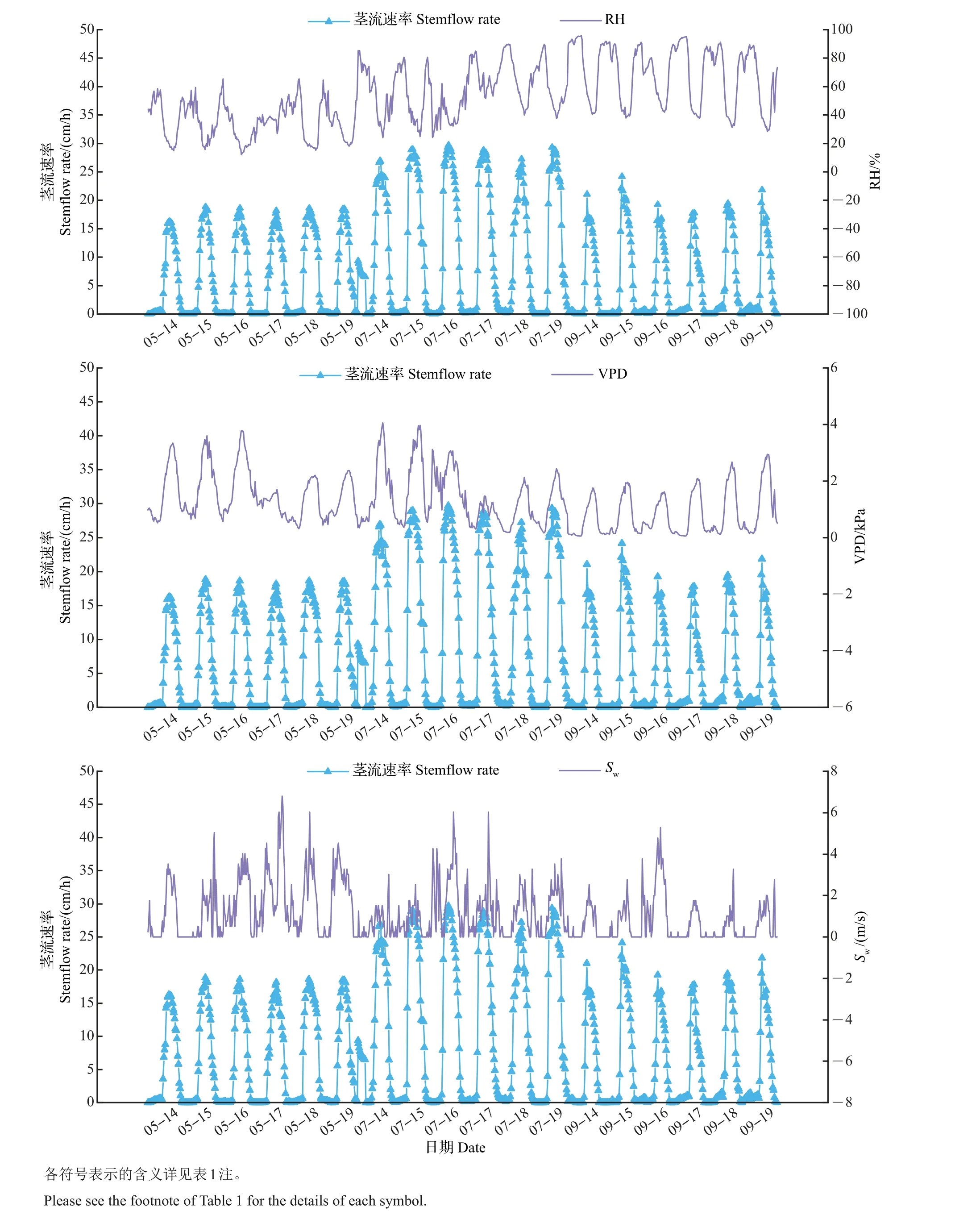
图3 富士苹果树干茎流速率与主要气象因子之间的关系Fig.3 Relationship between stemflow rate of red Fuji apple and main meteorological factors
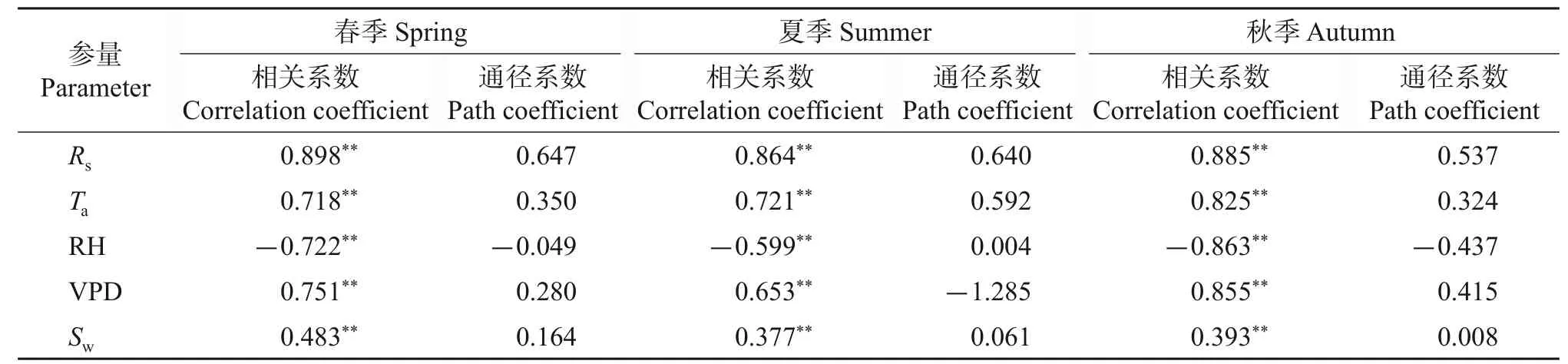
表4 不同季节茎流速率与气象因子的相关系数与通径系数Table 4 Correlation coefficient and path coefficient of stemflow rate with meteorological factors in different seasons
由于试验地果园苹果树根系主要分布在0~80 cm之间[21],因此,本试验对苹果生长期内10、30、50、70 cm 土层日均土壤体积含水量与日均茎流速率进行分析。由图4 可见:各土层含水量受天气变化和果园灌溉的影响,变化趋势和幅度差异较大。10 cm 处土壤含水量主要受果园灌溉影响,在生长期内变化幅度较大,其变化趋势和日均茎流速率的变化表现出较好的相关性;50 cm处土壤含水量从5月初到7月底呈现递减趋势,进入夏季,果园灌溉量增大,7月25日左右土壤含水量达到峰值后下降(生长期内30 cm 处土壤含水量的变化与50 cm 处基本一致,数据未显示);随着土层深度增加,70 cm处土壤含水量变化较稳定,总体上呈现先减后增的趋势,与环境因子空气相对湿度变化一致。对不同土层含水量在不同季节与日尺度下的茎流速率进行相关分析,结果如表5所示:在春季和夏季,10和30 cm处土壤含水量与日均茎流速率具有显著的相关性,在秋季相关性不显著;50 cm 处土壤含水量与秋季茎流速率显著相关。
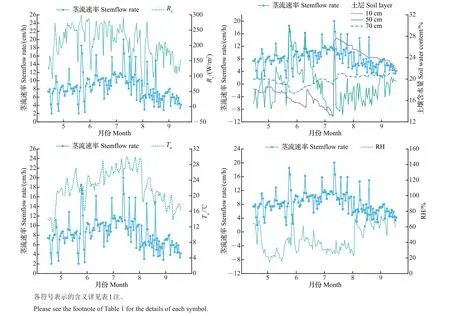
图4 生长期内富士苹果树干日均茎流速率与主要环境因子之间的关系Fig.4 Relationship between daily average stemflow rate of red Fuji apple trunk and main environmental factors during the growth period
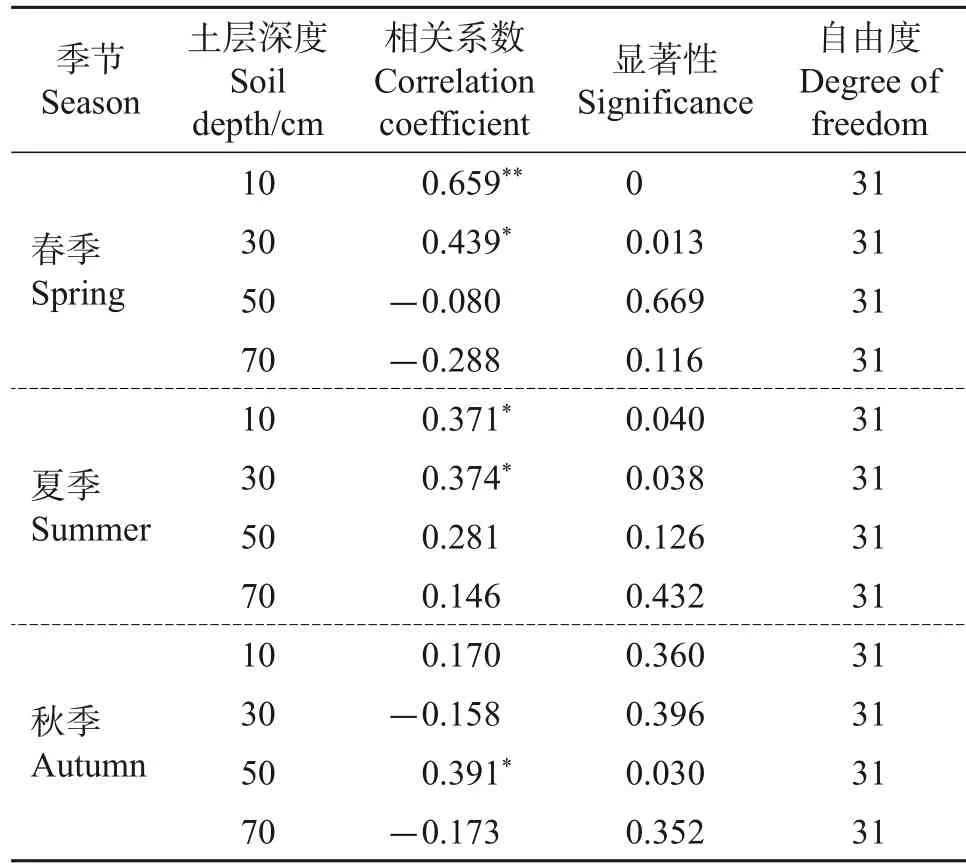
表5 日均茎流速率与不同深度土壤含水量的相关系数Table 5 Correlation coefficient between daily average stemflow rate and soil water content at different depths
基于生长期内5—9 月153 d 观测的茎流数据,对日尺度下的茎流速率与气象因子、土壤含水量、30 cm 处土壤温度进行分析。如表6 所示:整个生长期内,富士苹果树干日均茎流速率与太阳辐射、大气温度、水汽压差、土壤含水量、土壤温度呈极显著正相关;与空气相对湿度呈极显著负相关;与风速的相关性不显著。通径系数反映出水汽压差、太阳辐射是影响日均茎流速率季节变化的关键因子。
2.4 富士苹果树干茎流速率的环境因子响应模型
基于对富士苹果树干茎流速率连续动态监测,分别以3个季节(春季、夏季、秋季)瞬时尺度下的茎流速率为因变量,以气象因子为自变量进行逐步回归,得到3 个季节树干茎流速率的回归方程,如式(1)~(3)所示。以生长期(5—9 月)内日尺度下的茎流速率为因变量,以同时期日均气象因子、土壤因子为自变量进行逐步回归分析,得到整个生长期内树干茎流速率与环境因子的回归方程,如式(4)所示。
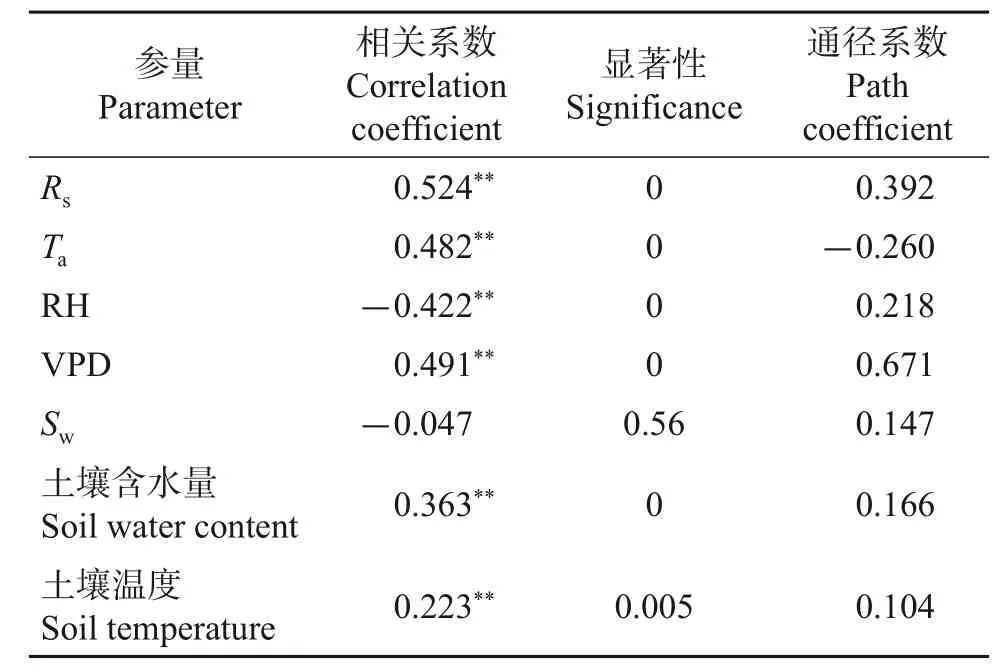
表6 生长期内富士苹果树干日均茎流速率与环境因子之间的相关系数与通径系数Table 6 Correlation coefficient and path coefficient between daily average stemflow rate of red Fuji apple trunk and environmental factors in growing season

式中:vsap为树干茎流速率,cm/h;Rs为太阳辐射强度,W/m2;VPD 为水汽压差,%;Ta为大气温度,℃;RH 为空气相对湿度,%;Sw为风速,m/s;Ts为30 cm处土壤温度,℃。以上4 个方程均达到了显著水平(P<0.01),复相关系数均在0.7 以上,拟合效果较好。由于春季、夏季和秋季建立模型的数据均来自典型晴天的数据,而季节耗水模型受阴雨天气数据的影响,其模型的拟合效果会出现一定的差异。
3 讨论
本研究结果表明,干旱绿洲区富士苹果树干茎流日变化在夏季和秋季主要呈双峰型曲线,春季主要呈单峰型曲线,具有明显的昼夜节律,不同季节树干茎流时空变化特征差异显著。这是由于极端干旱区太阳辐射极强,苹果树易遭受高温和干旱胁迫,植株通过调节叶片气孔闭合,增大气孔阻力,避免过多蒸腾耗水,而导致树干茎流速率下降,出现茎流峰值提前,或者进行午休而呈现双峰波型。夏季7 月茎流启动早,持续时间最长,峰值最大,树干日耗水量达到最大。李浩等[19]、刘毅等[22]的研究结果表明,树干茎流变化昼高夜低,昼夜差异显著,夏季茎流水平高于春秋季,本研究结果与此一致。这是由于夏季果树生长态势高,果实处于膨大期,加之太阳辐射强,树体需要大量水分维持生命活动,茎流变化活跃、持续时间长,茎流量大。本研究发现,生长期内5—9月树干茎流速率最大峰值出现的时间为13:00—15:00,在18:00—20:00 迅速降低。这与周玉燕等[4]对甘肃山旱塬区和崔俊芳等[23]对山东丘陵区苹果树生长期茎流的研究结果类似,而夏桂敏等[18]对东北冷凉地区寒富苹果树茎流的研究发现,峰值最早出现的时间为08:30,14:30左右迅速下降。这可能与不同地区存在时差及日照时长不同有关。
夜间茎流有利于调节树体水分平衡并驱动根系向上运输物质。本试验中生长期树干夜间存在微弱的茎流,呈时断时续的状态,与王力等[9]的研究结果一致。夜间茎流的存在可能是由于在白天树冠蒸腾失水过多导致树体内储存水较少,形成了根压差,促使水分主动被吸入树体,从而产生夜间补偿流,以此来补偿白天过度蒸腾散失的大量水分,从而维持植物体内的水分平衡。也有学者认为,树冠和大枝条是树木主要的储水部位,夜间茎流的存在是因为水流由根部上升至树冠储存,为下一时间段的蒸腾做准备[24-25]。至于本研究区苹果树夜间茎流的调控机制和夜间茎流是否有一部分用于夜间蒸腾,有待进一步研究。张婕等[26]的研究表明,夜间茎流对整天蒸腾贡献率可达40%;本研究发现,夜间茎流量占全天总茎流量的波动范围在1.06%~10.28%之间,并主要集中在日落后到次日00:30这个时间段内。可见,虽然夜间茎流量占比较小,但仍具有一定的调节植物与环境间水汽交换的能力。
气象因子对树干茎流变化具有重要影响,各气象因子在自然条件下并不是独立作用的,而是综合影响植物的蒸腾耗水过程[9]。众多研究表明,太阳辐射和水汽压差是驱动树木边材茎流动态的主要气象因子[9,19,27-29]。本研究发现,不同季节树干茎流速率对气象因子的响应具有明显差异,但无论在瞬时尺度下,还是日尺度下,太阳辐射和水汽压差始终是影响树干茎流速率的主导因子。有报道认为,太阳辐射对树干茎流的影响会表现出季节差异[30-31]。在本研究中这种差异并不明显,与党宏忠等[32]的研究结果一致,可能是由于生长期内灌水较充足,土壤水分并没有明显的亏缺。水汽压差是表征植物干旱胁迫下气孔调节行为开启的最主要的环境变量之一。夏桂敏等[33]的研究表明,在干旱荒漠区水汽压差对树干蒸腾的影响可能比太阳辐射强,本研究中不同尺度下不同季节茎流速率对环境因子的响应模型也印证了这一点,多元回归方程中水汽压差值的系数最大。尽管如此,树干茎流对环境因子的响应变化过程仍然要参照时间尺度,而土壤温度与土壤含水量对树干茎流的季节变化影响更显著[31]。本研究发现,整个生长期内土壤温度与日均茎流速率的相关性显著(r=0.223,P<0.01),不同季节茎流速率与不同深度土壤含水量在瞬时尺度下的相关性差异较大,而生长期内日均茎流速率与10 cm 处土壤含水量呈显著正相关(r=0.363,P<0.01)。说明不同环境条件下,在不同时间和空间尺度上果树蒸腾耗水的影响调控机制是复杂的、相对的。至于本文得出的蒸腾耗水模型的可靠性及预测值与实测值的误差率,还需进一步验证。
富士苹果树干在不同季节的多元线性回归模型的复相关系数均在0.8以上,说明这些模型可以模拟树干茎流速率对气象因子的响应特征;而生长期内日均茎流速率对环境因子的响应模型复相关系数为0.725,说明该模型可以较好地模拟某一季度或者整个生长季的蒸腾耗水量。日均茎流速率模型是基于整个生长期内日平均气象数据构建的模型,可以忽略瞬时尺度下的天气变化干扰,但构建模型的数据样本有所减少,模型的准确度较低。因此,构建一套系统科学、普遍适用的果树蒸腾耗水和调控机制模型,应基于不同时间尺度对环境因子和茎流速率的关系进行模拟。
4 结论
富士苹果树干茎流速率的日变化在夏秋季主要呈双峰型曲线,在春季主要呈单峰型曲线,夜间茎流速率变化稳定,并主要集中在日落后到次日00:30这个时间段内,昼夜差异显著。茎流速率时空特征表现出季节差异,夏季茎流启动早,停止晚,茎流速率大。太阳辐射和水汽压差是影响树干茎流的关键因子。不同季节树干茎流速率与气象因子的多元回归方程如下。春季:vsap=-1.758+0.014Rs+2.313VPD+3.21Sw;夏 季 :vsap=-5.927+0.24Rs+3.188VPD+0.273Ta;秋 季:vsap=23.455+0.016Rs-0.18RH-0.662Ta+3.716VPD。生长期内树干茎流速率与环境因子的多元回归方程为:vsap=0.758+0.014Rs-0.338Ta+0.974Ts+2.78VPD+0.498Sw。
新疆阿克苏地区富士苹果树在夏季7月蒸腾量较大,果树蒸腾耗水受太阳辐射、水汽压差、土壤含水量等环境因子的影响程度高,且此时段是本地区温度最高的时期,果园灌溉应注意在果树生长旺季,尤其是连续晴天的情况下及时补充灌水,保证果树在生长的关键时期不受水分胁迫而影响产量和品质,且选择在日出前或者日落后灌水,可以避免因强烈的蒸发而导致灌溉水散失。
