水曲柳FmPHV基因克隆及在形成层愈伤组织中的表达分析
2020-08-26张佳薇张旭肖英韩朝君刘华领张振峰梁楠松詹亚光
张佳薇 张旭 肖英 韩朝君 刘华领 张振峰 梁楠松 詹亚光


摘 要:该研究通过基因克隆获得了水曲柳PHV基因,并命名为FmPHV。生物信息学分析表明,水曲柳FmPHV基因编码区全长为2 112 bp,包含有一个完整的开放阅读框,其编码了一个由703个氨基酸组成的蛋白。亚细胞定位预测其主要存在于叶绿体中,为稳定亲水蛋白。保守域及同源分析表明,FmPHV与油橄榄、芝麻和烟草等物种的同源蛋白保守结构域同源性高达99%。在低温4 ℃条件下,使用50 mg·L-1吲哚丁酸(IBA)溶液对水曲柳树皮进行处理,获得了水曲柳形成层细胞,并进一步诱导获得形成层愈伤组织。对FmPHV基因的时空表达模式进行分析表明,FmPHV基因6月表达量最高,同时FmPHV能在芽中高表达,通过树皮获得的不同来源的愈伤组织相互比较可知,FmPHV在来源为形成层分生组织形成层愈伤组织中的表达量显著高于其在其他来源的愈伤组织中的表达。此外,对水曲柳幼苗瞬時过表达FmPHV基因,对其所在通路关键基因的表达特征进行分析,FmPHV瞬时过表达后,生长素相关基因表达下降,细胞分裂素相关基因表达上升,有利于芽的分化。以上结果表明,揭示了水曲柳FmPHV在水曲柳植株生长过程中的表达模式以及FmPHV过表达对芽再生通路各关键基因的调控情况,为研究水曲柳FmPHV调控生长发育的分子机制以及其在生长素和细胞分裂素响应通路中发挥的作用奠定基础。
关键词:水曲柳,PHV基因,生物信息学分析,瞬时侵染,形成层细胞
中图分类号:Q943
文献标识码:A
文章编号:1000-3142(2020)07-0963-14
Abstract: PHAVOLUTA(PHV)transcription factor is one of the most important members of homeodomain-leucine zipper Ⅲ (HD-ZIP) family,and plays an important role in the regulation of formation of plant stems apical and germ meristem. In this research,the PHV gene of Fraxinus mandshurica was acquired by gene cloning and was named by FmPHV. Detailed bioinformatics analysis showed that the length of FmPHV gene was 2 112 bp,and contained a complete Open Reading Frame (ORF) which encoded a protein with 703 amino acids. The FmPHV gene encoded a stable hydrophilic protein,the subcellular localization prediction of FmPHV protein showed the FmPHV was mainly present in the chloroplast. Conserved domain and homology analysis of FmPHV protein indicated that,the homology of FmPHV between the homology PHV proteins in other species,such as Olea europaean,Sesamum indicum and Nicotiana tabacum,was 99%. Under 4 ℃ condition,with the treatment of 50 mg·L-1 3-indolebutyric acid(IBA)solution,we obtained the cambium callus from cambium stem cells of Fraxinus mandshurica. Analysis of the gene expression pattern of FmPHV showed that the FmPHV was the highest expression in June. Furthermore,the expression of FmPHV in buds was stronger than which in any other tissues. Compared with the callus from different sources,which obtained from the bark of F. mandshurica,the expression level of FmPHV gene in callus,derived from meristem formation,was significantly higher than which from other sources. Moreover,overexpression the FmPHV gene in F. mandshurica seedlings will reduce the expression of auxin-related genes and increase the expression of cytokinin-related genes. This phenomenon indicated that the function of FmPHV gene was more beneficial to the process of plant shoot differentiation and regeneration. In summary,this research revealed the expression pattern of FmPHV gene during the growth and development in F. mandshurica. And revealed the expression pattern of key genes in plant shoot regeneration pathway,which were regulated by FmPHV gene. Our research reveals the potential value of the molecular characteristic of FmPHV gene in the response of auxin and cytokinin pathways during the process of the formation of plant callus generation.
1 材料与方法
1.1 实验材料来源与处理
水曲柳材料取自东北林业大学实验林场,以成熟水曲柳植株树皮以及种子萌发所获得的30 d幼苗为实验材料,根、茎、叶取自水曲柳3年生盆栽苗,5月—9月水曲柳全株(包括根,茎,叶)及雄花雌花取自野生水曲柳用于基因定量测定。对水曲柳树皮进行消毒后,置于4 ℃中,使用50 mg·L-1IBA高渗溶液进行20 h处理后,放入诱导培养基培养两周后,获得水曲柳形成层愈伤组织。将材料至于液氮中冷冻保存。
1.2 实验方法
1.2.1 FmPHV基因编码区全长的克隆 使用CTAB方法提取水曲柳组培苗总RNA。经反转录获得cDNA。实验前期对水曲柳转录组测序获得水曲柳转录组数据库。应用BioEdit 软件中Local blast 功能中的blastn 和 blastx 功能,比对分析确认PHV基因序列并设计引物,对PHV编码区全长序列进行克隆(引物如表1所示)。使用Omega胶回收试剂盒对特异性扩增产物进行回收纯化,使用pEASY-T5载体连接特异产物,转化克隆进入Trans-T1感受态细胞中。送至生工生物工程(上海)公司测序。
1.2.2 FmPHV基因编码区序列的生物信息学分析 用NCBI在线工具中的Open Reading Frame (ORF) Finder预测FmPHV基因的编码区(https://www.ncbi.nlm.nih.gov/orffinder/)。使用SignalP5.0在线分析软件对FmPHV蛋白进行信号肽的预测和分析(http://www.cbs.dtu.dk/services/SignalP/)。利用ExPASy中ProtParam工具对该蛋白理论等电点pI(isoelectric point)值、亲水性/疏水性等理化性质进行分析(https://web.expasy.org/protparam/、https://web.expasy.org/protscale/)。使用在线分析软件对FmPHV蛋白的跨膜结构域进行分析(https://embnet.vital-it.ch/software/TMPRED_form.html)。利用WOLF-PSORT在线软件对FmPHV蛋白的亚细胞定位情况进行预测分析(https://wolfpsort.hgc.jp/)。利用 NCBI数据库Conserved Domain Search Service(CD Search)在线分析软件分析FmPHV的保守结构域。使用MEGA7.0.2软件的Neighbor-Joining(NJ)算法对其他物种中同源性较高的PHV蛋白构建系统发育进化树。利用在线多序列比对软件PRALINE对不同物种的PHV结构域进行多重序列比对。
1.2.3 水曲柳外植体无菌材料的获得 选择并收集一年生或多年生水曲柳茎条。截剪长度为4~6 cm的茎段在流水下冲洗2~4 h,将清洗过的组织放无菌烧瓶中,75%乙醇以及5%的次氯酸钠(日本Junsei)对外植体消毒。材料接种于MD201培养基(WPM+4.0 g·L-1琼脂+20 g·L-1蔗糖+1 mg·L-1毒莠定+1 mg·L-16-BA),并于黑暗条件培养7 d。
1.2.4 实时荧光定量对水曲柳不同部位及瞬时侵染植株中FmPHV和相关基因表达量分析 以水曲柳微管蛋白基因Tu作为内参基因(Livak et al.,2001),引物见表2。PCR反应体系如下:10 μL Master Mix,2 μL模板cDNA,10 μmol·L-1的正反向引物各1 μL,用ddH2O补足至20 μL。扩增反应利用Applied Biosystems7500荧光定量PCR仪进行,反应程序:95 ℃30 s、95 ℃5 s、60 ℃ 34 s(共40個循环)、95 ℃15 s、60 ℃1 min、95 ℃15 s。每份样品进行3次重复。目标基因表达水平运用相对定量的2-ΔΔCt(Livak et al.,2001)进行计算,式中:Ct是热循环仪检测到反应体系中荧光信号的强度值。
2 结果与分析
2.1 FmPHV基因全长克隆与序列分析
以水曲柳实生苗全株的cDNA为模板进行PCR反应扩增,获得一条大小为2 100 bp左右的片段,扩增结果见图1。经测序,通过NCBI数据库blastn比对,确定获得水曲柳PHV基因序列,并命名为FmPHV。FmPHV基因的开放阅读框全长为2 112 bp,编码了703个氨基酸(图2)。经过比对,与油橄榄(Olea europaea)OeATHB14基因核苷酸序列(XM_023011038.1)的一致性为99%;与芝麻(Sesamum indicum)SiATHB14基因核苷酸序列(XM_011098376.2) 的一致性为99%;与烟草(Nicotiana tabacum) NtPHV基因核苷酸序列 (JQ686932.1)的一致性为94%。所以我们确定水曲柳中该基因为HD-ZIP Ⅲ转录因子家族成员PHV,并命名为FmPHV。
2.2 FmPHV蛋白质结构分析
2.2.1 FmPHV蛋白质一级结构分析 利用ProtParam工具在线分析,FmPHV编码703个氨基酸,相对分子质量为76.89 kD,等电点(pI)为6.27,不稳定系数为55.08,确定为不稳定蛋白(不稳定系数大于40时,预测蛋白质不稳定,反之则稳定)。FmPHV蛋白的亲水/疏水性分析结果表明,FmPHV蛋白平均亲水系数(GRAVY)为-0.182,为亲水蛋白。此外,FmPHV蛋白亲疏水性图谱显示,FmPHV蛋白共有47个疏水区和59个亲水区(图3:A)(>0.5区域为疏水区,<-0.5区域为亲水区,介于+0.5~-0.5之间主要为两性区域)。跨膜结构域预测结果表明,FmPHV存在两个跨膜结构区域,分别在453~471 bp氨基酸序列片段上存在从里到外的跨膜结构区域,以及445~467 bp氨基酸序列片段上存在的从外到里的跨膜结构区域,分数分别为889和515(分数>500为显著跨膜结构),由内到外与由外到内的跨膜结构域基本重合,并且都有强烈的跨膜螺旋结构,由此可推断,FmPHV蛋白既可以具有由外到内的跨膜能力,也具有由内到外的跨膜能力(图3:B)。信号肽预测结果表明,FmPHV蛋白可能不存在信号肽,我们认为该蛋白为非分泌蛋白。利用WOLFPSORT预测FmPHV亚细胞定位,结果表明,FmPHV主要分布在叶绿体中,占57.14%,其次分布在细胞核中,占28.57%,少部分分布于囊泡中,占4.28%。
2.2.2 FmPHV蛋白的二级结构预测 利用SOPMA在线工具分析FmPHV蛋白的二级结构组成,结果显示蛋白二级结构主要由43.67%(307 aa)的无规卷曲(random coil)、36.84%(259 aa)的α螺旋结构(alpha helix)、14.51%(102 aa)的扩展长链(extended chain)、4.98%(35 aa)的β转角结构(beta turn)构成(图4)。
2.2.3 FmPHV蛋白氨基酸序列比对及保守结构域的预测 分别以FmPHV蛋白的氨基酸序列为基础,在NCBI数据库中检索与水曲柳FmPHV蛋白同源性较高的其他物种的氨基酸序列进行比对。包括油橄榄(Olea europaea)、可可(Theobroma cacao)、木薯(Manihot esculenta)、胡桃(Juglans regia)、烟草(Nicotiana tabacum)、芝麻(Sesamum indicum)、紫花风铃(Handroanthus impetiginosus)、紫瓶子草(Sarracenia purpurea)、细叶石竹(Nicotiana sylvestris)、牵牛(Ipomoea nil)、绿豆(Vigna radiata var. radiata)、大豆(Glycine max)、水蜜桃(Prunus persica)、甜樱桃(Prunus avium)、马铃薯(Solanum tuberosum)、长蒴黄麻(Corchorus olitorius)、亮叶桦(Betula luminifera)、土瓶草(Cephalotus follicularis)、柑橘(Citrus sinensis)、中粒咖啡(Coffea canephora)。采用MEGA7.0.2中的NJ方法构建蛋白的系统发育进化树,结果显示进化树聚为两组,其中水曲柳FmPHV与同属木犀科的油橄榄OeATHB14亲缘关系最为紧密(图5)。
利用在线多序列比对软件PRALINE对FmPHV蛋白序列与NCBI数据库中一些其他物种的PHV蛋白序列进行同源性比较,结果表明,FmPHV与其他PHV(ATHB14)蛋白序列的保守区主要集中在3个结构域,分别为HD、b-ZIP和START结构域(图6)。包括油橄榄(Olea europaea)(XP_022866806.1)、野生烟草(Nicotiana attenuata)(XP_009612342.1)、可可(Theobroma cacao)(EOY25496.1)、芝麻(Sesamum indicum)(XP_020554026.1)、 中粒咖啡 (Coffea canephora)灰色阴影区域为HD结构域;下横线区域为b-ZIP结构域;直角框内为START结构域;“*”代表终止密码子。
Gray shadow region is the HD domain; the lower horizontal line is the b-ZIP domain;right angle box is the START domain;“*” represents the stop codon.(CDP17314.1)、葡萄(Vitis vinifera)(RVW23993.1)的PHV(ATHB14)氨基酸序列蛋白高度一致。
2.3 水曲柳形成層愈伤组织的获得及培养
2.3.1水曲柳形成层愈伤组织的获得 获得水曲柳无菌茎段外植体后,为达到分离韧皮部和形成层的目的,使用50 mg·L-1的高浓度IBA(indole-3-butytric acid)溶液,在4 ℃条件下对已与木质部分离的其他部分处理20 h后,将材料放入0.5 mg·L-1IBA低渗溶液中处理15 min。之后将茎段置于MD201培养基(WPM+4.0 g·L-1琼脂+20 g·L-1蔗糖+1 mg·L-1毒莠定+1 mg·L-16-BA)中暗培养7 d后,取出,将木质部完全去除后将靠近韧皮部一侧置于MD201培养基上,暗培养7 d后即获得水曲柳形成层愈伤组织,如图7所示。从左至右分别为一年生树皮形成层愈伤组织(部分整齐排列的致密白色松散细胞团)、一年生树皮韧皮部愈伤组织(随机分布的偏绿色松散细胞团)、多年生树皮形成层愈伤组织(整齐排列的致密白色松散细胞团)、二年生树皮韧皮部愈伤组织(不启动)。
2.3.2水曲柳形成层愈伤组织的培养 实验为形成层愈伤组织继代培养选择了WPM、B5、MSB5三种培养基进行继代培养基优化,同时对形成层愈伤组织和普通皮层愈伤组织的生长量进行了比较(图8)。
实验选取WPM、1/2 WPM、B5、MSB5四种培养基对水曲柳形成层愈伤组织进行继代培养,其中B5培养基中材料出现不同程度的褐化,两次继代后材料死亡,在1/2 WPM培养基中材料长势缓慢,在褐化情况出现之前死亡。MSB5培养基中材料长势迅速升高,但随即材料排出大量多酚,2~3 d后死亡。我们初步判断,水曲柳形成层愈伤组织不适应富含丰富有机物质的培养基。WPM培养基中材料生长势虽然不如MSB5培养基,但是生长状态良好,显微镜下观察到松散细胞团,继代周期为21 d(图8:A,C)。
在对形成层愈伤组织和树皮愈伤组织在WPM培养基中进行继代20 d左右后,观察到极少数树皮边缘处出现与形成层愈伤组织完全不同的乳黄色团状紧密愈伤组织(图8:B)。本实验进一步对两种愈伤组织的生长量进行统计(图8:C)。结果表明,形成层愈伤组织的启动周期显著优于树皮愈伤组织,并且前者的生长量是后者的4倍之多。
由于水曲柳形成层愈伤组织具有生长迅速,细胞团处于松散易分离状态等类似于胚性细胞的优良特性,我们决定通过探究芽再生关键基因FmPHV在其中的表达情况,为水曲柳芽再生体系的建立奠定基础。
2.4 FmPHV基因的表达分析
2.4.1 FmPHV基因在水曲柳中表达分析 分别取实验室培养的水曲柳愈伤组织和水曲柳种子组培培养30 d的幼苗。根、茎、叶取自水曲柳3年生盆栽苗。5月—9月混合样(包括根,茎,叶)及雄花雌花取自野生多年生水曲柳,以及以形成层为来源的形成层愈伤组织(图9:A,B,C)。
对水曲柳FmPHV基因在不同部位中定量表达分析结果表明(图9:B),该基因在芽中高度表达,而在其他组织中根、茎、叶、花以及种子中的表达量普遍不高。形成层愈伤组织中FmPHV基因的表达量是普通培养愈伤组织表达量的22.35倍,说明FmPHV对调控形成层的愈伤组织形成起到了重要作用。
水曲柳快速生长期为每年的5月—9月,5月复苏,6月萌芽,同时FmPHV表达量在6月达到最高,在9月表达量最低,6月表达量是9月的29.738倍(图9:A)。可以推测该基因在水曲柳芽发生阶段发挥着正调控作用。同时形成层愈伤组织中该基因特异性高表达,推测该种愈伤组织具有芽再生潜力,可以作为水曲柳再生体系建立材料的来源。
为进一步证明FmPHV在芽再生过程中的正向作用,分别选取水曲柳种子萌发过程中0、2、4、6、8、10、12以及14 d的实生苗作为实验材料,对水曲柳种子萌发过程中的FmPHV表达情况进行实时荧光定量PCR分析(图9:C)。结果表明,在种子萌发过程中,FmPHV在第2天和第8天分别达到两个峰值,分别是第0天的11.379倍和10.236倍。在芽再生过程中,关键基因的表达模式被发现为两次峰值出现的现象,而这两次峰值分别与水曲柳种子芽的发生阶段和芽伸长阶段相互对应。FmPHV基因的表达与水曲柳,进一步说明该基因在芽再生过程中的重要作用。
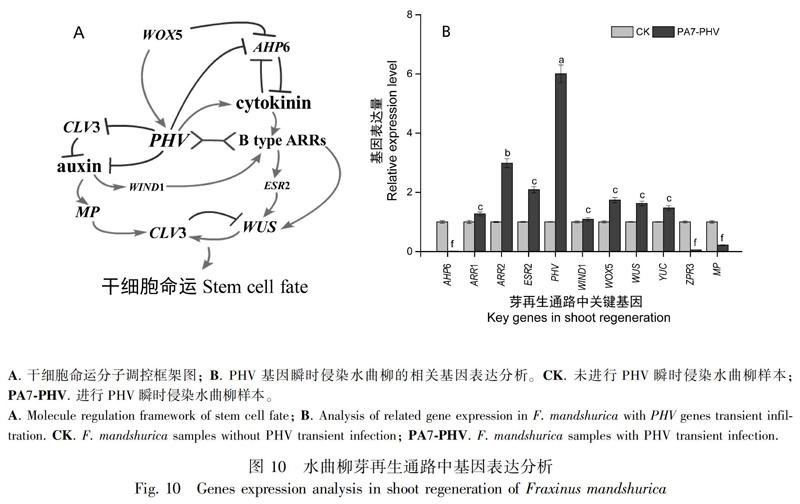
2.4.2 FmPHV瞬时侵染水曲柳对芽再生通路基因表达的影响 前人的研究表明,PHV(ATHB14)受到包括WOX5,细胞分裂素通路相关的AHP6和生长素通路相关的YUC5等生长素以及细胞分裂素多个通路的关键基因的调控,此外,PHV能够进一步调控细胞维持与芽再生过程的重要基因,包括ARR1,ARR2,WIND1,ESR2以及WUS等(图10:A)。为了揭示水曲柳中FmPHV对下游基因的调控关系,我们应用35S:PHV-GFP过表达载体,对水曲柳实生苗进行瞬时过表达实验。
结果表明,瞬时过表达后,FmPHV表达量显著提高,而FmPHV下游基因AHP6,ZPR3和MP呈现出显著的下调表达,与之对应的是,包括ARR2,WOX5,ESR2,YUC和WUS 等基因表达量呈现出不同程度的升高(图10:B)。值得注意的是通路中受PHV直接抑制的YUC表达量轻微上调现象,我们认为PHV可能分别通过不同通路直接或间接地抑制生长素的积累。同样引起我们注意的还有,与HD-ZIP Ⅲ转录因子家族存在共同作用的B型ARRs家族成员ARR2的高表达情况,HD-ZIP Ⅲ-B型ARRs复合物与WUS启动子结合,并起到进一步激活WUS启动的重要作用。同时受PHV直接负调控和通过细胞分裂素间接调控等多种通路同时抑制的AHP6的低表达,以及受生长素直接调控的MP的低表达,我们认为这表明在PHV高表达后,存在生长素表达水平下降,细胞分裂素表达水平上升的现象,这种变化在细胞微环境中更利于芽分化。
3 讨论与结论
本研究通过生物软件,对FmPHV蛋白进行了预测和生物信息学分析。经预测分析结果表明FmPHV蛋白为稳定亲水蛋白,亚细胞分析表明,其可能主要存在于叶绿体中。通过对 FmPHV蛋白进行的结构域分析,表明了FmPHV蛋白有一个保守START结构域以及C末端序列高度保守氨基酸序列。与Ursache et al.(2014)的研究結果类似,
PHV基因编码植物特有的b-ZIP类转录因子,通过接受相关靶基因的调控或者蛋白之间的互作而发挥着重要的调控功能。
将FmPHV的氨基酸序列应用NCBI网站的Blastp进行比对,比对结果表明,FmPHV蛋白与油橄榄、芝麻、烟草、马铃薯、番茄、葡萄、毛果杨、蓖麻、可可、拟南芥等物种的对应蛋白的同源性较高。同时应用比对结果构建了系统进化树。进化树表明,水曲柳FmPHV 蛋白与同科同属植物油橄榄中的同源蛋白在进化上亲缘关系较近。
FRANKENSTEIN C,ECKSTEIN D,SCHMITT U,2005. The onset of cambium activity - A matter of agreement[J]. Dendrochronologia,23(1):57-62.
HU B,ZHANG G,LIU W,et al.,2017. Divergent regeneration-competent cells adopt a common mechanism for callus initiation in angiosperms[J]. Regeneration,4(3):132.
HUANG T,YAEL HARRAR,LIN C,et al.,2014. Arabidopsis KANADI1 acts as a transcriptional repressor by interacting with a specific cis -element and regulates auxin biosynthesis,transport,and signaling in opposition to HD-ZIP Ⅲ factors[J]. Plant Cell,26(1):246.
JIA X,DING N,FAN W,et al.,2015. Functional plasticity of miR165/166 in plant development revealed by small tandem target mimic[J]. Plant Sci,233:11-21.
JING T,MA WL,KUJANSUU J,et al.,2002. Sprouting regeneration of Fraxinus mandshurica[J]. J Beijing For Univ,24(4):12-15.[荊涛,马万里,Joni Kujansuu,等,2002. 水曲柳萌芽更新的研究[J]. 北京林业大学学报,24(4):12-15.
KONG DM,PREECE JE,SHEN HL,2012. Somatic embryogenesis in immature cotyledons of Manchurian ash (Fraxinus mandshurica Rupr.)[J]. Plant Cell Tissue Organ Cul,108(3):485-492.
LIBAK KJ,SCHMITTGEN TD,2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method.[J]. Methods,25(4):402-408.
MULLER CJ,ANA EV,WANG G,et al.,2016. PHABULOSA mediates an auxin signaling loop to regulate vascular patterning in Arabidopsis[J]. Plant Physiol,170(2):956.
OTSUGA D,DEGUZMAN B,PRIGGE MJ,et al.,2001. REVOLUTA regulates meristem initiation at lateral positions[J]. Plant J,25(2):223-236.
PEREZ-PARRON MA,GONZALEZ-BENITO ME,PEREZ C,1994. Micropropagation of Fraxinus angustifolia,from mature and juvenile plant material[J]. Plant Cell Tissue Organ Cul,37(3):297-302.
PFEIFFER A,JANOCHA D,DONG Y,et al.,2016. Integration of light and metabolic signals for stem cell activation at the shoot apical meristem[J]. eLife,5:e17023.
PRIGGE MJ,2005. Class Ⅲ Homeodomain-Leucine Zipper gene family members have overlapping,antagonistic,and distinct roles in Arabidopsis development[J]. Plant Cell Online,17(1):61-76.
QIN YF,LI DD,LI XB,2009. Research progress on plant HD-Zip transcription factors[J]. Chin J Cell Biol,(4):514-520.[秦永芳,李登弟,李學宝,2009. 植物HD-Zip转录因子研究进展[J]. 细胞生物学杂志,(4):514-520.]
REINHART BJ,LIU T,NEWELL NR,et al.,2013. Establishing a framework for the Ad/Abaxial regulatory network of Arabidopsis:ascertaining targets of Class Ⅲ HOMEODOMAIN LEUCINE ZIPPER and KANADI regulation[J]. Plant Cell,25(9):3228-3249.
RUONALA R,KO D,HELARIUTTA Y,2017. Genetic networks in plant vascular development[J]. Ann Rev Genet,51(1):annurev-genet-120116-024525.
SANG YL,CHENG ZJ,ZHANG XS,2018. Plant stem cells and denovo organogenesis[J]. New Phytol,218:1334-1338.
SESSA G,CARABELLI M,et al.,2018. Multiple links between HD-Zip proteins and hormone networks[J]. Internat J Mol Sci,19(12).
SMITH ZR,LONG JA,2010. Control of Arabidopsis apical-basal embryo polarity by antagonistic transcription factors[J]. Nature,464(7287):423-426.
THAKUR R,SOOD A,2006. An efficient method for explant sterilization for reduced contamination[J]. Plant Cell Tissue Organ Cul,84(3):369-371.
URSACHE R,MIYASHIMA S,CHEN Q,et al.,2014. Tryptophan-dependent auxin biosynthesis is required for HD-ZIP Ⅲ-mediated xylem patterning[J]. Development,141(6):1250-1259.
YANG L,BIAN L,2013. Somatic embryogenesis and plantlet regeneration from mature zygotic embryos of Manchurian ash (Fraxinus mandshurica Rupr. )[J]. Plant Cell Tissue Organ Cul,115(2):115-125.
ZENG FS,ZHOU S,ZHAN YG,et al.,2014. Drought resistance and DNA methylation of interspecific hybrids between Fraxinus mandshurica,and Fraxinus americana[J]. Trees,28(6):1679-1692.
ZHANG TQ,LIAN H,TANG H,et al.,2015. An intrinsic microRNA timer regulates progressive decline in shoot regenerative capacity in plants[J]. Plant Cell Online,27(2):349-360.
ZHANG TQ,LIAN H,ZHOU CM,et al.,2017. A two-step model for de novo activation of WUSCHEL during plant shoot regeneration[J]. Plant Cell,29(5):1073.
ZHANG LJ,ZHANG LW,FENG DD,et al.,2007. In vitro culture of axillary bud sprouting of Fraxinus mandshurica[J]. Bull Bot Res,27(3):319-324.[张丽杰,张丽玮,冯丹丹,等,2007. 水曲柳腋芽离体快繁研究初报[J]. 植物研究,27(3):319-324.]
ZHANG Z,TUCKER E,HERMANN M,et al.,2017. A molecular framework for the embryonic initiation of shoot meristem stem cells[J]. Dev Cell,40(3):264.
ZHONG RQ,YE ZH,2001. Alteration of auxin polar transport in the Arabidopsis ifl1 mutants[J]. Plant Physiol,126(2):549-563.
ZHU Z,QI F,YAN C,et al.,2016. Sexually different morphological,physiological and molecular responses of Fraxinus mandshurica flowers to floral development and chilling stress[J]. Plant Physiol Biochem Ppb,99:97-107.
(責任编辑 周翠鸣)
