模拟干旱胁迫下六个种源梓树种子萌发特性及地理变异研究
2020-08-26何润华李媛肖遥王军辉李志辉麻文俊
何润华 李媛 肖遥 王军辉 李志辉 麻文俊


摘 要:該研究采用聚乙二醇(PEG-6000)模拟干旱胁迫,测定了发芽率、发芽势和胚根长度等指标,运用相关性分析、聚类分析和隶属函数法综合评价梓树六个种源种子萌发期的抗旱性大小,初步筛选出抗旱性较强的种源。结果表明:随着溶液水势的降低,六个种源种子的发芽率、发芽势、相对发芽率、发芽指数、活力指数表现为持续下降趋势;除河南洛阳种源的胚根、胚轴长度以及甘肃正宁种源的胚根长度外,其余种源胚根、胚轴长度均表现为逐渐下降趋势;溶液水势为-1.0 MPa时,河南洛阳种源失活,其余种源各指标趋于一致。将各种源种子发芽特性与原产地地理环境因子进行相关分析表明,种子活力指数与经度、纬度呈显著正相关(0.903和0.871),经度、纬度较大的种源活力更强、抗旱性更佳;年均气温越高,种子发芽率和活力均降低;年降雨量与种子的发芽率和活力指数呈中偏弱的负相关,但与胚根和胚轴的生长呈中等正相关;温度和降雨量对梓树种子的抗旱性有一定影响,主要体现在年均温和年降雨量越低,种子的发芽率也越低;初步发现处于干旱和寒冷等相对恶劣环境下的梓树种源表现为以提高种子发芽率和降低种子胚生长活性的调节机制来适应不良环境。运用聚类分析及隶属函数法对六个种源梓树种子萌发期进行抗旱性综合评价,辽宁恒仁种源种子抗旱性最强,河南洛阳种源种子抗旱性最弱。综上结果认为,梓树六个种源抗旱性差异明显,温度和降雨量在一定程度上影响了种子萌发的抗旱性。
关键词:梓树,种源,PEG胁迫,种子萌发,抗旱性
中图分类号:Q948
文献标识码:A
文章编号:1000-3142(2020)07-0944-10
Abstract: In this study,polyethylene glycol (PEG-6000) was used to simulate drought stress,the germination percentage,germination potential and radicle length were measured. Correlation analysis,cluster analysis and membership function method were used to comprehensively evaluate six provenances of Catalpa ovata. The most resistant sources of drought resistance were screened good rootstocks out for the grafting of Chinese C. bungei,to promote the breeding,and to lay a material foundation for its large-scale application. The results were as follows:The germination rate,germination potential,relative germination rate,germination index and vigor index of six provenances of C. ovata seeds decreased with a trend continuous decline. Except for the radical length,hypocotyl length of the Luoyang in Henan provenance,and the radical length of the Zhengning in Gansu provenance. The length of radicle and hypocotyl of other provenances showed a trend of gradual decrease. When the osmotic stress was -1.0 MPa,the Luoyang in Henan provenance was deactivated,and the indexes of another provenances tendency to be consistent. The correlation between the germination characteristics of each group and the geographical environment factors of origin indicated that there was a significant positive correlation between vigor index,longitude and latitude (0.903 and 0.871). The higher average annual temperature,the lower seed germination rate and vigor. Annual precipitation was negatively correlated with seed germination rate and vigor index,but moderately positive correlated with the growth of radicle and hypocotyl. Temperature and precipitation affected the drought resistance of seeds of C. ovata to some extent,mainly reflected in the lower annual temperature and annual precipitation,the lower germination rate of seeds. It was preliminarily found that provenances of C. ovata in the relatively harsh environment of drought and cold showed the regulation mechanism to improve seed germination rate and reduce the growth activity of seed embryo to adapt to the unfavorable environment. The results of cluster analysis and membership function method were used to comprehensively evaluate the drought resistance of C. ovata seeds in six groups. The seeds of Liaoning in Hengren provenance had the strongest drought resistance,while the Luoyang in Henan provenance had the weakest drought resistance. In summary,the drought resistance of six provenances of C. ovata is obvious,temperature and rainfall affect the drought resistance of seed germination to some extent. The provenance with strong drought resistance are selected to be the rootstocks of C. bungei breeding and lays a material foundation for the large-scale application of C. bungei.
Key words:Catalpa ovata,provenance,PEG stress,seed germination,drought resistance
嫁接是无性系繁殖的有效措施之一,已被广泛应用于植物繁育。决定嫁接成败的主要因素除嫁接方式外,砧木与接穗的匹配性是影响接穗生长的关键。近年来,已有研究表明砧木类型影响嫁接苗的生长和抗性。王新建等(2008)研究发现金丝楸砧木嫁接苗提高了楸树的抗性,且显著大于自砧苗和灰楸砧木嫁接苗。孙胜等(2010)研究表明金瓜、中国南瓜、葫芦三种砧木中,以葫芦为砧木能显著提高西瓜叶片中的营养含量。此外,还有报道证实合适的砧木类型有效改善了接穗的抗旱性(李兴亮等,2018)、耐盐性(Podda et al.,2017)、耐寒性(李惠等,2017)和耐涝性(唐玲玲等,2016)。由此可见,选择适宜的砧木能大大提高接穗的各项抗性指标。楸树(Catalpa bungei)是原产于我国的古老优质珍贵用材和观赏树种(岑显超,2008),由于其硬枝插条的生根率低(杨如同等,2014),所以主要采用芽接的方式进行嫁接扩繁。梓树(C. ovata)与楸树同属,具有根系发达、抗性强等特性,与楸树接穗具有良好的匹配性,一直以来楸树嫁接都采用梓砧(陈丛梅等,2007;王苏珂等,2008)。目前,我国年产楸树嫁接苗在1 000万株以上,并广泛应用于国家储备林、园林绿化、通道绿化等工程项目。但是,在规模化嫁接繁育中出现了新的问题,如梓树种源与楸树品种的亲和性以及不同梓树种源的抗性差异等问题,这在一定程度上影响了楸树嫁接苗的生产应用。因此,开展梓树种源抗性评价,是拓展楸樹应用范围和提升良种使用水平的有效途径。
种子萌发期既是植物生活史的关键时期,又是衡量其抗旱性的重要时期,能直接决定植株后期的生长状态(李培英等,2010)。有研究表明,种子抗旱性和苗期抗旱性具有高度相关性(于卓等,1997;王飞等,2014;吉庆勋等,2017),种源间种子萌发差异可能是由海拔高度、经度、纬度、气候等因素造成的(Matthies,1990;Herrera,1991;Brad et al.,2004)。本研究选取梓树六个种源种子进行PEG模拟干旱胁迫处理,分析PEG胁迫对不同种源种子萌发和芽苗生长的影响,并评价种源的抗旱性差异,以期为楸树优质嫁接苗的繁育及良种推广提供技术支撑。
1 材料与方法
1.1 材料
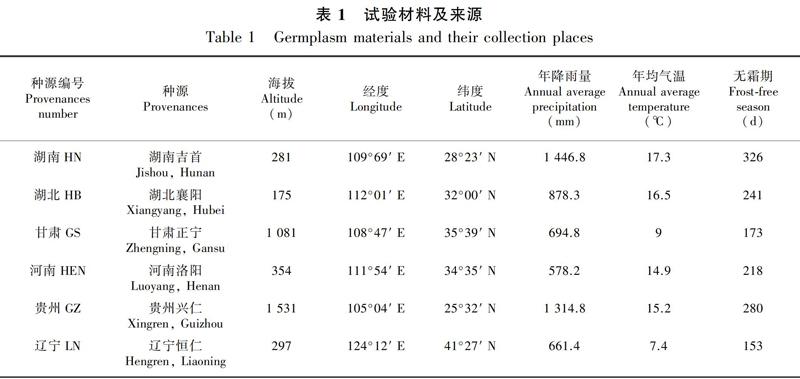
2014年,从梓树主要分布区的6省6县(市、区)采集种子,每个采种点的采种母树为10株。母树树干通直、生长良好、无病虫害侵袭,达到正常的开花结实年龄,各母树间直线间隔大于100 m。同一采种点采集的种子混合均匀,作为供试种源材料。各种源编号、地理位置、气候环境状况见表1。
1.2 试验设计
以不同浓度的PEG-6000溶液模拟干旱胁迫,溶液水势为干旱程度的度量指标,设置4个梯度,分别为0.0(CK)、-0.5、-1.0、-1.7 MPa,每种处理设置4次重复,每重复100粒。
1.2.1 种子预处理 播种前一天,每个种源种子用浓度为5‰的高锰酸钾溶液消毒1 h,用自来水将高锰酸钾冲洗干净,40 ℃初温的水浸泡(让其自然冷却)24 h。
1.2.2 播种和干旱胁迫处理 根据试验设计,先用不同水势的PEG溶液浸湿滤纸,然后分别将不同处理的滤纸铺两张在培养皿底部,并将种子整齐摆放在滤纸上面,培养皿内的溶液高度以达到种子厚度的1/3为准,对照组(CK)处理用蒸馏水培养。利用电子天平给每个培养皿称重(精度为0.01 g),并进行编号。
1.2.3 培养条件 将PEG不同处理下的种子放入光照培养箱(HPG-280BX)进行培养。光照:荧光灯光强150 μmol·m-2·s-1、28 ℃、8:00至18:00;黑暗:22 ℃,18:00至次日8:00。
1.3 种子发芽率调查和芽苗测定
1.3.1种子发芽调查 培养处理后,每天观察记录发芽的种子数,以胚根长度和种子长度相等,胚芽长度为种子长度一半时作为种子的发芽标准(李晓洁和徐化成,1989)。每天定时记录种子的发芽个数,连续2周不再有种子发芽时记录结束。
1.3.2 芽苗测定 在培养梓树种子第12天测定发芽种子胚根和胚轴的长度,测定精度为0.1 cm。在每种处理下,每个重复选择长势均等的10个芽苗进行测定。
1.4 指标测定与数据分析
利用Excel 2010 和SPSS 22.0软件进行数据处理、统计分析和制作图表。使用方差分析检验不同种源梓树种子性状和芽苗生长是否存在显著差异,采用Duncan法进行多重比较。
相关生长指标的具体计算公式如下:
发芽率(G):G=nN×100%。
式中:n为发芽种子数; N为种子样本数(孙时轩,1992)。
相对发芽率(%)=(G/对照发芽率)×100%。
式中:G为种子发芽率。
发芽势=发芽高峰期(一般以最初1/3天数内发芽种子数计)发芽种子数/供试种子数×100%。
发芽指数(Gi):Gi=∑(GtDt)。
式中:Gt为时间t日的发芽数; Dt为相应的发芽天数(郑光华,2004)。
活力指数(VI):VI=s×Gi。
式中:s为幼苗鲜重(郑光华,2004)。
1.5 抗旱性综合评价方法
采用模糊隶属函数法进行抗旱性综合评价(李源等,2010;任文佼等,2013)。同时,为了合理评价梓树各种源对PEG胁迫的敏感性,采用种子发芽临界水势的处理数值作为评价指标。通过隶属函数公式,计算各指标的隶属函数值。当指标与抗旱性呈正相关时,用公式(1);当指标与抗旱性呈负相关时,用公式(2)。
式中:μ(Xij)为i种源j指标的隶属函数值;Xij为i种源j指标的值;Xjmax为各种源j指标值中的最大值;Xjmin为该指标中的最小值。
2 结果与分析
2.1 干旱胁迫对不同种源梓树种子发芽特性的影响
图1结果显示,HEN种源在不同程度胁迫处理下发芽情况均显著低于其余种源。当溶液水势为0和-0.5 MPa时,LN种源发芽率均显著高于其余种源(图2:A),但各水势下HB、HN、GS种源发芽率差异不显著,GZ、HEN种源发芽率显著低于其他种源。当水势为-1.0 MPa 时,六个种源发芽均受到明显的抑制,其中HEN种源发芽率趋近于0,其余5个种源发芽率均处于较低水平。当水势达到-1.7 MPa时,除GS种源有较低发芽率外,其余5个种源发芽率基本为0。从种子的相对发芽率(图2:B)来看,HEN种源相对发芽率下降幅度最大,且在水势为-1.0 MPa时明显低于其余5个种源,在-1.7 MPa胁迫强度下,GS种源相对发芽率明显高于其余种源,其余5个种源相对发芽率趋近于0。从发芽率和相对发芽率来看,LN和GS种源具有较强的抗旱性。
表2结果显示,随着PEG胁迫的增强,供试种子的发芽势逐渐降低,说明在PEG干旱胁迫处理下,种子的发芽高峰期滞后,但不同种源种子萌发对干旱胁迫响应不同。GS、HB、LN种源具有普遍高的发芽势,但HN、HEN、GZ种源发芽势较低,其中LN种源下降幅度最大,与对照相比下降了74.75%。当水势为-0.5 MPa时,HEN种源的发芽势明显低于其余种源,仅为0.50%;当水势达到-1.0 MPa时,只有HB、GZ种源有1.00%和0.25%的发芽势,其余种源都为0。这表明HB、GZ抗旱潜力相对较大。
从图3可以看出,当水势为0和轻度胁迫(-0.5 MPa)时,LN种源发芽指数均为最高,但当水势为-1.0 MPa时,发芽指数降幅较大。HB、GS和HN种源发芽指数随着水势降低降幅小于LN种源,且该指标处于较高水平,说明这三个种源的种子存在潜在的抗旱性。六个种源的种子发芽指数随着干旱胁迫程度的增大变化趋势基本一致。
活力指数同每粒种子的相应发芽时间和整齐度均有联系,活力强的种子在受胁迫的环境下出苗能力强,贮存后能保持萌发能力,活力弱的种子则相反。种子活力不是单一的指标,而是从各方面描述种子出苗能力的综合指标(陈蕾太等,2016)。随着水势的降低,六个种源种子活力指数下降的趋势有所不同(图4)。其中:GS及HEN种源的种子活力指数在受到轻度胁迫(-0.5 MPa)时迅速下降,变化幅度最为明显,其余4个种源下降趋势基本一致;GS和GZ种源在下降过程中经历一个平台期,其余4个种源则受到明显抑制;六个种源中,LN种源活力指数降幅最大,HEN种源在水势为-1.0 MPa时已失去活性。这表明GS和GZ种源在一定程度的干旱胁迫下仍能保持较强的种子活力。
2.2 干旱胁迫对不同种源梓树芽苗生长的影响
表3结果显示,HEN种源胚根和胚轴长在水势为0时显著低于其余种源,且处于极小值。这表明HEN种源种子生长势较低,可能会影响后期苗木的生长。在对照条件下,GZ和HB种源初生根长显著高于其余4个种源,GS、HN和LN种源的初生根长没有显著性差异且显著高于HEN种源;当水势降到-1.0 MPa时,GZ和HB种源初生根长仍显著高于LN和HN,说明这两个种源种子发芽后生长势较强。
不同抗旱类型梓树胚轴生长速度是鉴定抗旱能力的一项重要指标。在对照条件下,仅HEN种源胚轴长显著低于其余5个种源,当水势达到-0.5 MPa时,各种源胚轴长已受到明显抑制但彼此无显著差异;当水势降低至-1.0 MPa时,GS、HB、GZ种源胚轴长显著高于LN和HN种源,其中HB种源受抑制作用最为明显。从种子胚根和胚轴生长来看,HEN种源种子抗旱性最弱。
2.3 不同种源梓树种子各指标与地理海拔及气候因子的相关分析
一般认为植物在其分布范围内的不同区域,由于地理气候条件的差异和植物对其响应不同,所以在自然选择过程中会逐渐形成不同的遗传结构,从而产生地理变异(徐化成,1992)。将不同种源各指标与相应种源地的地理海拔与气候因子进行相关分析,表4结果表明,除胚根、胚轴长度以外,其余5项指标与海拔高度和年均气温呈负相关,与地理经度、纬度呈正相关,其中种子活力指数与经度、纬度呈显著正相关,相关系数分别高达0.903和0.871。大致呈现出随着海拔高度与年均气温的增加,种子萌发对干旱胁迫的敏感性逐渐增加的变化趋势。从与气候因子的相关分析(表4)可以看出,年均氣温低的种源种子萌发对干旱胁迫较敏感,其与发芽率(-0.689)、发芽势(-0.623)呈较强的负相关,年降雨量则与胚根长和胚轴长呈中度正相关,表明种源地水分充足使得种子具有更强的胚生长性。此外,在一定范围内无霜期越长,种子萌发后生长状况越好,这可能是改变其遗传效应的环境机制。
2.4 不同种源梓树种子的抗旱性综合评价
植物的抗旱机制是一个复杂的体系,但种子的萌发过程是一个复杂的生理生化过程,单一的指标很难评定某类植物抗旱性的强弱。因此,评价植物抗旱性时需要进行多个指标的综合判断(李培英等,2010)。选择PEG水势为-1.0 MPa的各指标数据,用模糊隶属函数法对六个种源梓树种子发芽率、发芽势、相对发芽率、发芽指数、活力指数以及胚根、胚轴长进行综合评价,得到六个梓树种子材料的隶属函数总平均值,并根据均值的大小进行排序(表5)。其中:LN种源综合隶属函数值最高(达0.866),表明其抗旱性最强;相反,HEN种源的抗旱性最弱。
综合分析不同种源梓树种子对干旱胁迫的敏感性,以欧氏距离为标准对六个种源进行聚类分析,结果见图5。由图5可知,六个种源可归为三个类群:LN、HN、GS种源聚为一类,通过抗旱性综合分析发现,这三个种源抗旱性相对较强,在胁迫处理下这三个种源抗旱性主要体现在种子具有较高的发芽率和发芽势等方面;HB和GZ种源归为一类,抗旱性次之,但其特点为种子胚根和胚轴生长能力极强,可能原因是这两个种源自身的遗传物质主要调控着种子的生长,激发种子发芽的基因表达能力可能较弱;HEN种源单独成为一类,各项发芽指标及生长指标均最差,抗旱性在六个种源间最弱。
3 讨论与结论
植物对干旱胁迫的响应取决于其生存环境的条件及水分胁迫的时长和强度(Dale,1988)。由于同一物种不同种源的母树为了适应原产地环境变化而产生遗传变异,并将稳定的遗传变异性状反映在种子的各方面特质中,所以同一物种不同种源种子的质量和芽苗生长等性状均存在差异(杨晓霞等,2016)。本研究结果表明,不同种源梓树种子发芽率与萌发后生长均存在显著差异。种子萌发期是衡量植物抗旱性强弱的重要时期,水分是影响种子质量和发芽的重要因素(Chamorro et al.,2016)。单一将发芽率作为种子萌发品质指标存在很大的局限性,它只能反映种子在适宜条件下的发芽状况,不能反映出芽苗的质量和生长势。同时结合发芽势、相对发芽率、发芽指数等一系列指标,既能反映植物种子的发芽水平,又能表现出种子的发芽整齐度及芽苗质量是否良好(关世超和王丕武,2011)。通过PEG模拟干旱胁迫促使梓树种子表现出受旱现象,随着水势的降低,发芽率持续降低,这与于军等(2010)对不同浓度PEG处理下沙冬青种子发芽率降低的研究结果一致。当水势达到-1.0 MPa时,对六个种源梓树种子萌发期各项指标影响明显,河南洛阳种源梓树种子已经失去活力;而当PEG水势达到-1.7 MPa时,各种源种子基本受到完全抑制,表明-1.0 MPa已达到梓树种子萌发的临界点。但是,河南种源种子即使在正常水分条件下,其发芽率、活力指数、发芽指数以及发芽后胚的发育和生长也均呈较弱的状态。造成这种现象的原因较为复杂,下一步将通过解剖途径观测种胚的发育情况,从本研究所缺乏的对种胚发育形态学方面来探究差异形成的原因。
本研究中,六个种源种子萌发和芽苗生长对干旱胁迫的影响差异明显,且通过地理环境因子与种源发芽特性的相关分析表明,种子活力指数与经度、纬度显著正相关,这说明梓树种子的萌发和生长具有依赖于经度、纬度的地理变异模式。魏胜利等(2008,2009)指出,温度适宜和年降雨量丰富地区梓树种源的采种母树在生长期未受到胁迫,其机体也未产生相应的应对干旱胁迫的防御机制;相反,出现干旱、气温过高地区梓树种源的采种母树在逆境下已经产生了特有的响应机制,且这种机制已经形成稳定的遗传效应,通过种子不断的传递下去。综合相关分析和胁迫条件下各种源的发芽及胚生长情况发现,种源地环境较差的梓树种子表现出较高的发芽率和较弱的胚生长活性,这类种源具有更高的存活率,这可能是梓树种子的一种干旱机制。本研究采用聚类分析和模糊隶属函数值法筛选出抗旱性最强种源为辽宁恒仁,抗旱性种源的成功筛选为楸树良种壮苗的规模化繁育奠定了物质基础。
参考文献:
BRAD RM,BROWN A,CHRISTOPHER RD,et al.,2004,Geographical gradients in seed mass in relation to climate[J]. J Biogeogr,31(3):379-388.
CEN XC,2008. The physiology reaction of different species(var.) of Catalpa bungei under drought stress[D]. Nanjing:Nanjing Forestry University.[岑显超,2008. 不同品种(类型)楸树苗木对干旱胁迫的生理响应[D]. 南京:南京林业大学.]
CHAMORRO D,PARRA A,MORENO JM,2016. Reproductive output,seed anatomy and germination under water stress in the seeder Cistus ladanifer subjected to experimental drought[J]. Environ Exp Bot,123(2016):59-67.
CHEN CM,LI SM,SHEN YP,et al.,2007. Technical of grafting seedling for Catalpa bungei C.A.Mey and cultivation technology of Catalpa rootstock[J]. Fore Sci Technol,(11):22-23.[陳丛梅,李淑梅,申艳普,等,2007. 楸树嫁接育苗与梓砧培育技术[J]. 林业科技通讯,(11):22-23.]
CHEN LT,SUN AQ,YANG M,et al.,2016. Seed vigor evaluation based on adversity resistance index of wheat seed germination under stress conditions[J]. J Appl Ecol,27(9):2968-2974.[陈蕾太,孙爱清,杨敏,等,2016. 基于小麦种子发芽逆境抗逆指数的种子活力评价[J]. 应用生态学报,27(9):2968-2974.]
DALE JE,1988. The control of leaf expansion[J]. Ann Rev Plant Physiol Mol Biol,39(1):267-295.
GUAN SC,WANG PW,2011. A brief talk on the method for determining the vigor of genetically modified soybean seeds[J]. Seed World,(9):20-21.[关世超,王丕武,2011. 浅谈转基因大豆种子活力测定方法[J]. 种子世界,(9):20-21.]
HERRERA CM,1991. Dissecting factors responsible for individual variation in plant fecundity[J]. Ecology,72(4):1436-1448.
JI QX,CHEN T,HAN S,et al.,2017. Effects of NA-MY additives on seed germination drought resistance of maize seedling[J]. Acta Agric Nucl Sin,31(2):364-370.[吉庆勋,陈涛,韩松,等,2017. NA-MY助剂对玉米萌发及苗期抗旱性的影响[J]. 核农学报,31(2):364-370.]
LI XL,ZHANG JK,LI MJ,et al.,2018. Evaluation and gene expression analysis of different apple dwarfing stocks on drought resistance[J]. J Agric Biotechnol,26(3):401-409.[李兴亮,张军科,李民吉,等,2018. 不同类型苹果矮化砧木抗旱评价与基因表达分析[J]. 农业生物技术学报,26(3):401-409.]
LI H,LIANG MM,ZHAO S,et al.,2017. Effects of different rootstocks on cold resistance of “Lyuling” walnut[J]. N Hortic,(3):25-31.[李惠,梁曼曼,赵爽,等,2017. 不同砧木对‘绿岭核桃抗寒性的影响[J]. 北方园艺,(3):25-31.]
LI PY,SUN ZJ,ABLT,2010. Evaluation of drought resistance of 29 accessions of Elytrigria repens at seed germination stage under PEG-6000 stress[J]. Chin Acta Agr Sin,32(1):32-39.[李培英,孙宗玖,阿不来提,2010. PEG模拟干旱胁迫下29份偃麦草种质种子萌发期抗旱性评价[J]. 中国草地学报,32(1):32-39.]
LI XJ,XU HC,1989. A study on seed and germination characteristics of Pinus bungeana Zucc. et Endi. from different provenances[J]. Sci Silv Sin,25(2):97-105.[李曉洁,徐化成,1989. 白皮松种子发芽习性及其种源变异的研究[J]. 林业科学,25(2):97-105.]
LI Y,LIU GB,GAO HW,et al.,2010. Comprehensive evaluation of salt tolerance of alfalfa germplasm and physiological response under salt stress[J]. Acta Pratac Sin,19(40):79-86.[李源,刘贵波,高洪文,等,2010. 紫花苜蓿种质耐盐性综合评价及盐胁迫下的生理反应[J]. 草业学报,19(40): 79-86.]
MATTHIES D,1990. Plasticity of reproductive components at different stages of development in the annual plant Thlaspi arvense L.[J]. Oecologia (Berlin),83(1):105-116.
PODDA L,SANTO A,LEONE C,et al.,2017. Seed germination,salt stress tolerance and seedling growth of,Opuntia ficus-indica (Cactaceae),invasive species in the mediterranean basin[J]. Flora,229:50-57.
REN WJ,LI QH,WANG SX,et al.,2013. Difference in phy-siological characters of seedling and salt-tolerance of Artemisia sphaerocephala of different provenances under salt stress[J]. J NE For Univ,41(2):10-14.[任文佼,李清河,王賽宵,等,2013. 盐胁迫下不同种源籽蒿的生理生化特性与耐盐性评价[J]. 东北林业大学学报,41(2):10-14.]
SUN S,TIAN YS,LENG DD,et al.,2010. Effects of different kinds of rootstocks on economic yields and mineral nutrition contents of leaves of grafted watermelon seedlings[J]. Plant Nutr Fert Sci,16(1):179-184.[孙胜,田永生,冷丹丹,等,2010. 不同砧木对嫁接西瓜经济产量及叶片矿质营养含量的影响[J]. 植物营养与肥料学报,16(1):179-184.]
SUN SX,1992. Silviculture[M]. Beijing:China Forestry Publishing House.[孙时轩,1992. 造林学[M]. 北京:中国林业出版社.]
TANG LL,XIANG XQ,YANG JP,et al.,2016. Submergence tolerance of ‘LD-1 stock-grafted Actinidia Deliciosa Miliang 1[J]. Guihaia,36(6):646-650.[唐玲玲,向小奇,杨江平,等,2016.‘LD-1砧米良一号猕猴桃的耐涝性[J]. 广西植物,36(6):646-650.]
WANG SK,WANG JH,ZHANG SG,et al.,2008. Study on seed germination characteristic among different combination of Catalpa bungei[J]. For Res,21(2):275-278.[王苏珂,王军辉,张守攻,等,2008. 楸树不同交配组合种子发芽特性的研究[J]. 林业科学研究,21(2):275-278.]
