长江中下游湖泊蚌类β 多样性分解 及其保护对策探讨
2020-07-02王魏根李永民
王魏根,李永民
(阜阳师范大学生物与食品工程学院,安徽,阜阳 236037)
由于水体污染、栖息地破坏、过度利用和生态入侵等多种原因,淡水生物多样性面临严重的威胁[1]。蚌类是淡水生态系统食物网中连接底栖群落和浮游群落的一个重要环节,通过生物扰动促进物质循环和能量流动[2]。蚌类资源对于人类也具有很高的经济价值。蚌类营底栖生活,运动迟缓,受栖息地环境变化影响大,是淡水生物类群中最容易受到威胁的类群之一[3]。全球蚌类资源的衰退也逐渐引起社会的关注和重视[4]。
长江中下游平原拥有独特的浅水湖群,形成复杂的河流-泛滥平原生态系统,有丰富的淡水物种资源[5]。长江中下游地区是东亚蚌类的分布中心,我国淡水蚌类特有种类58 种,主要就分布在长江流域一带[6]。近些年来长江中下游湖泊水体受到强烈干扰,水体污染、栖息地丧失以及水生生境破坏等因素导致蚌类资源急剧降低。舒凤月根据物种保护价值指数初步评估显示,长江流域蚌类处于受威胁等级的有21 种,处于近危等级的有8 种,许多蚌类的分布区明显缩小[7]。从20 世纪50 年代起,对长江流域一些湖泊贝类进行的物种调查和区系研究为蚌类资源保护奠定了基础[8-10]。近年来,一些研究也对长江流域蚌类资源分布现状及保护措施提出了一些建议[7,11-12],但这些研究主要集中在α多样性水平进行分析,不能提供更多该区域蚌类群落分布变化信息。
β 多样性反映了物种组成在空间上的变化[13],了解物种β 多样性背后的形成机制可以为区域生物多样性保护计划提供重要参考[14]。Legendre 提出一种通过计算由位点-物种构成的群落组成资料矩阵的方差(VAR)来估计总β 多样性(BDTotal)的方法,总β 多样性可以分解成单一物种对总β 多样性的贡献度 SCBD(Species Contribution to Beta Diversity)和单个位点对总β 多样性的贡献度LCBD(Local Contribution to Beta Diversity)。某位点LCBD 值大则表示该样点群落组成和区域内其它位点组成差异大,也说明该位点具有较高的保护价值,相反则说明该位点物种贫乏或者环境被破坏需要修复。某物种SCBD 值大小则反映该物种对该区域β 多样性影响的相对重要性[15]。根据群落间β 多样性来源的相关理论,Baselga 基于Jaccard 相异性指数提出总β 多样性(βjac)可分解成物种在空间上周转(βjtu)和嵌套(βjne)两个部分,即βjac= βjtu+βjne,分别代表在空间上的物种替代和物种得失两个不同的过程[16]。如果β 多样性格局以嵌套为主,则需要优先保护物种多样性丰富度高的小区域;相反,以周转为主的则需要同时保护多个小区域[14]。因此,我们采用两种β 多样性分解方法,对长江中下游地区部分湖泊淡水蚌类多样性组成资料进行β 多样性分析,探讨该区域蚌类β 多样性变化的模式,从一个新的角度为该区域淡水蚌类物种多样性保护和资源合理利用提供参考依据。
1 材料与方法
1.1 研究区概况
长江中下游地区属于亚热带季风气候,年平均气温13~20℃,年平均降水量800~1600mm[5]。根据已发表的文献,本次共编辑整理长江中下游18 个湖泊的蚌类名录资料进行β 多样性分析,分属湖南、湖北、江西、安徽、江苏省和上海市。湖泊分布地理位置及概况见表1。
在这18 个湖泊中,共记录了螺类名录70 种,隶属于6 个科,蚌科(Unionidae)种类最多59 种,其次是蚬科(Corbiculidae)6 种,球蚬科(Sphaeriidae)2 种,截蛏科(Solecurtidae)、贻贝科(Mytilidae)和珍珠贝科(Margaritanidae)各1 种。蚌科的背角无齿蚌(Anodonta woodiana)和圆顶珠蚌(Unio douglasiae)在18 个湖泊中都有分布,17 个物种仅在单个湖泊中有记录。在各个湖泊中,鄱阳湖记录蚌类种数最多为62 种,保安湖最少为5 种。各湖泊蚌类物种分布主要参考文献有:湖南省的洞庭湖[8,10]、大通湖[17];湖北省的洪湖[10,18]、梁子湖、牛山湖、保安湖、扁担塘[10]、东湖[10,19]、涨渡湖[20]、花马湖[21];江西省的鄱阳湖[10,22-23]、青岚湖[24]、军山湖[25];安徽省的巢湖[26]、菜子湖[27]、南漪湖[28];江苏的太湖[9-10,29]和上海的淀山湖[30]。
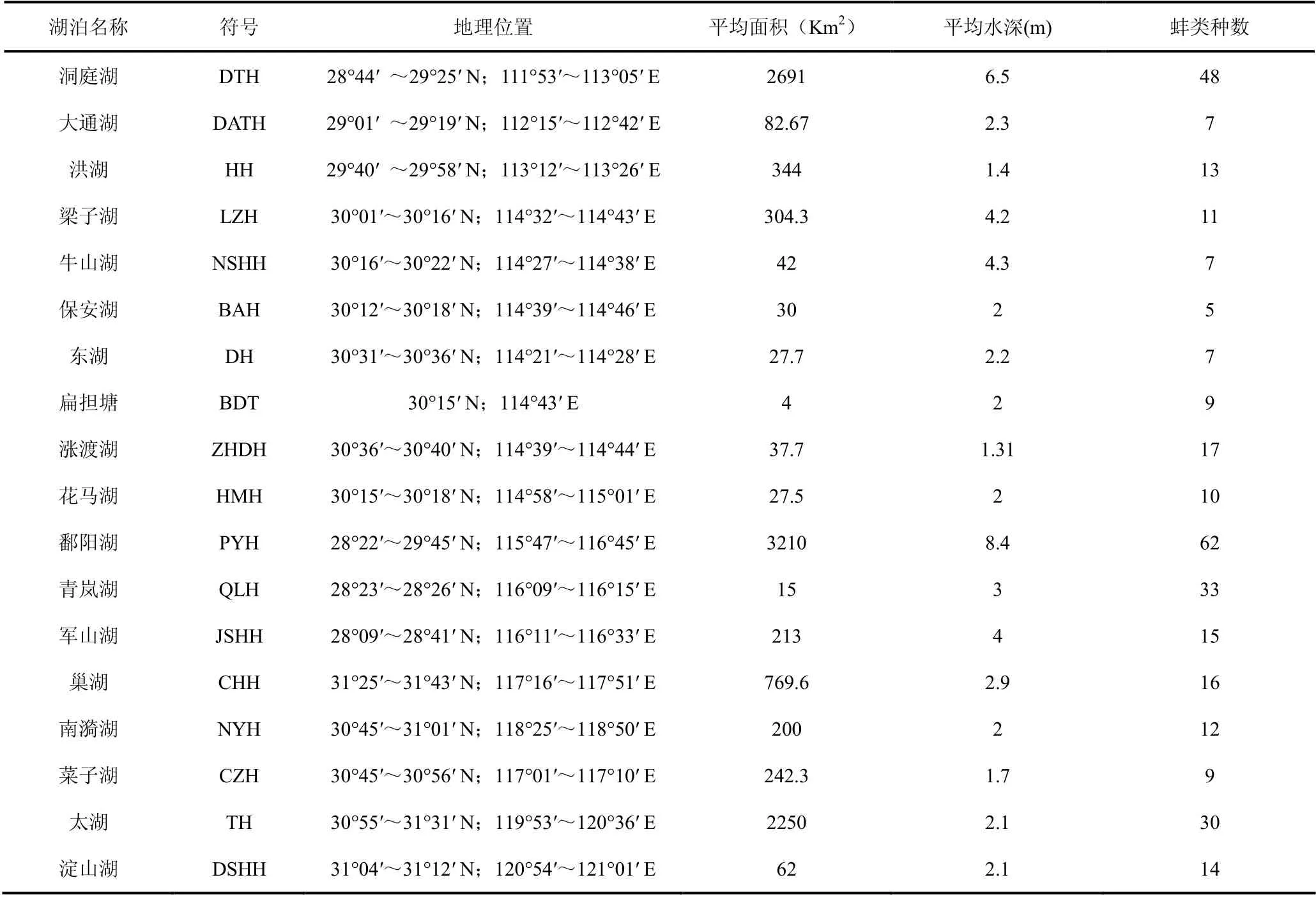
表1 长江中下游湖泊特征及蚌类物种数 Table 1 The number of bivalves species in the lakes of mid-lower reaches of the Yangtze River
1.2 数据整理
我们主要依据20 世纪60 年代以来公开发表文献的记录报告进行编辑整理并构建长江中下游湖泊淡水蚌类物种名录及分布状况数据库(γ 多样性)。数据整理成二元数据表,每行代表一个湖泊,每列代表一个物种,某蚌类物种在该湖泊有记录就标记为“1”,没出现则标记为“0”。有的物种有多个亚种,则将所有亚种归为一个物种,例如蚶形无齿蚌(Anodonta arcaeformis arcaeformis)、黄色蚶形无齿蚌(A. arcaeformis flavotincta)都统一记录为蚶形无齿蚌。不同文献中记录的同一物种中文名不同,根据其拉丁名统一为同一物种。有的文献中记录的物种没有鉴定到种名,这些记录则不包括在本次数据分析中。
1.3 β 多样性分解方法
采用Legendre 方法度量单一物种对总β 多样性的贡献度(SCBD)和单个位点对总β 多样性的贡献度(LCBD),即计算湖泊螺类组成矩阵总方差(VAR)作为总β 多样性值(BDTotal),然后再分解为单个物种和单个湖泊对总β 多样性贡献度[15],计算公式见表2。
采用Baselga 分解方法量化β 多样性及其周转和嵌套组分,即利用Jaccard 相异性指数,分别采取多位点和配对计算方法,分析整个区域湖泊以及各湖泊对之间蚌类总β 多样性(βjac)及其周转(βjtu)和嵌套组分(βjne)。配对方法的β 多样性计算公式见表2,相应的多位点计算方法(βJAC、βJTU和βJNE)请参阅文献[16]。
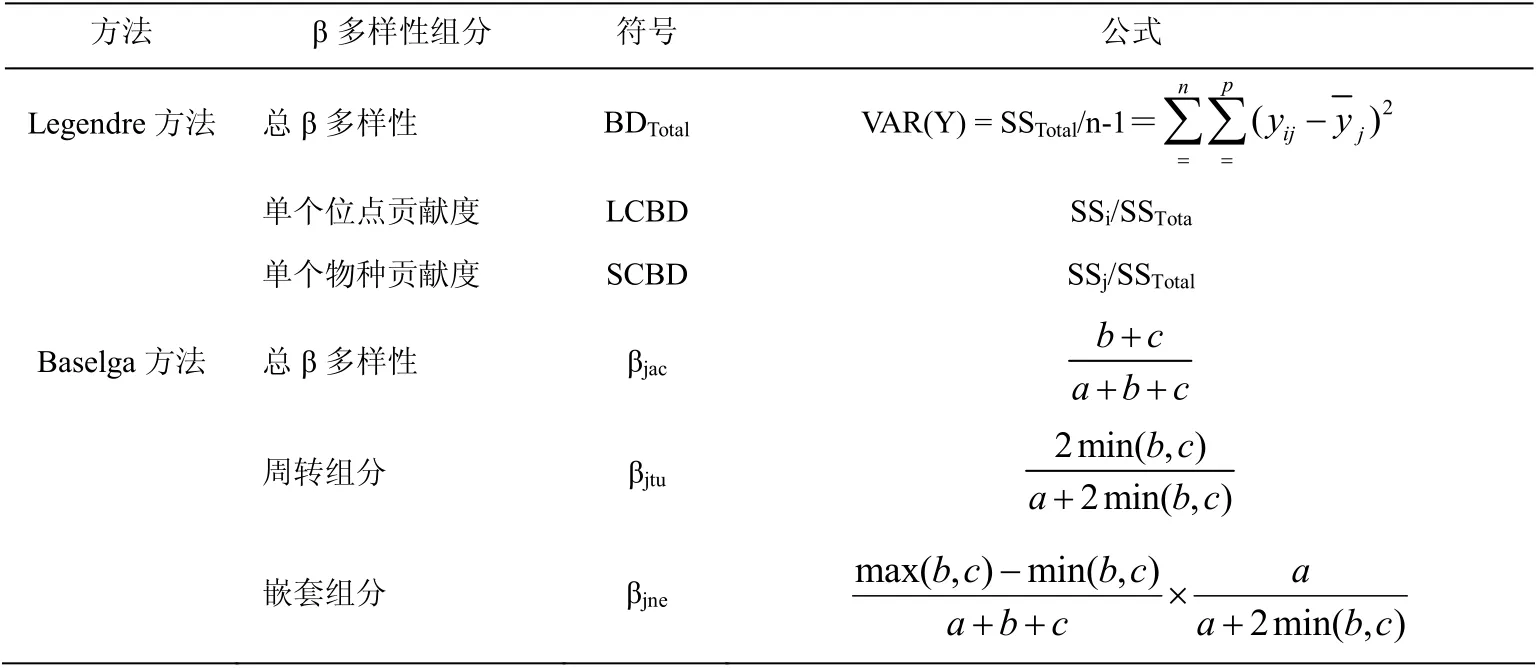
表2 β 多样性分解方法 Table 2 The decomposition method of beta diversity
1.4 数据分析
Legendre 方法中单一物种对总β 多样性的贡献度(SCBD)和单个样方对总β 多样性的贡献度(LCBD)分析过程用R 语言工具包adespatial 中的beta.div 函数完成。计算总方差前,蚌类组成数据先进行“hellinger”转化,使相异性矩阵符合Euclidean 距离特征。LCBD 值的显著性检验采用置换检验,置换次数为999。对LCBD 值与各个湖泊面积、水深以及其中蚌类物种数进行相关分析[31]。Baselga 方法β 多样性及其周转和嵌套组分计算分析过程采用R 语言工具包“betapart”完成[32]。为了可视化β 多样性及其组分分布,用beanplot 图展示β 多样性及其组分度量值和分布密度[33]。利用配对计算的相异性矩阵采用“vegan”R 语言工具包进行非度量多维尺度分析(NMDS)和聚类分析(UPGMA 法)展示不同湖泊对之间的β 多样性及其组分间关系[34]。以上统计分析过程均在R 语言环境中进行[35]。
2 结果
2.1 单一螺类SCBD 和单一湖泊LCBD
采用Legendre 的β 多样性分析方法计算表明,长江中下游18 个湖泊蚌类总离均差平方和(SSTotal)为8.3983,总β 多样性值(BDTotal)为0.4940。
SCBD 最小值为0.001814,有河蛏蚌(Solenaia rivularis)和环带尖丽蚌(Aculamprotula zonata),这两种蚌类均只分布在鄱阳湖。最大值为0.05366,由湖球蚬(Sphaerium lacustre)贡献,该物种在8个湖泊有分布。SCBD 中位值为0.01120,平均值为0.01429,有25 个物种超过平均值。SCBD 值分布情况见图1、图2。
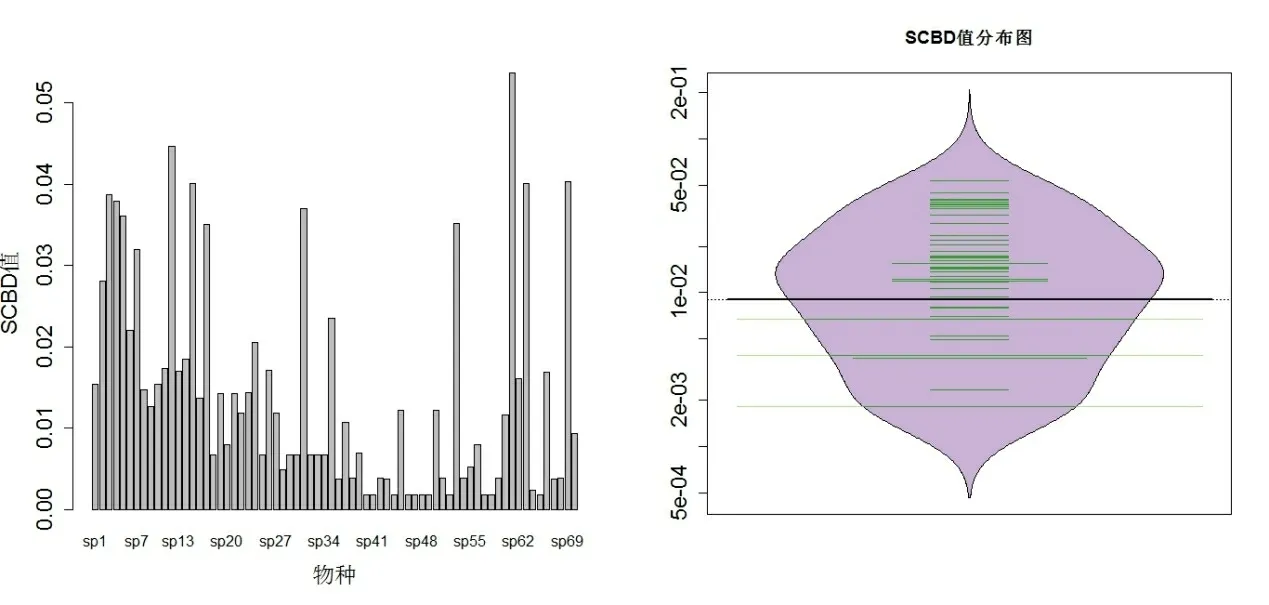
图1 SCBD 值及分布 Fig.1 The value and distribution of SCBD
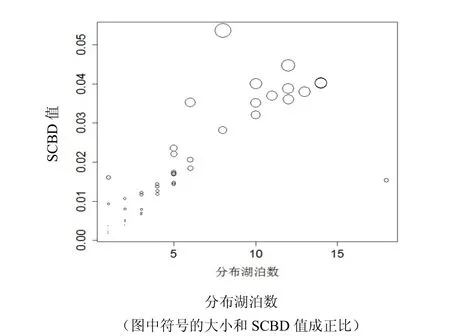
图2 SCBD 值与分布湖泊数关系 Fig.2 The correlation between species SCBD value and occupied lake numbers
LCBD 值分布情况见图3。最小为0.03648,由淀山湖贡献;最大值为0.07994,由南漪湖贡献。各个湖泊平均贡献度为0.05556,中位值为0.05589,有8 个湖泊LCBD 贡献度低于平均水平(从低到高依次是DSHH、HH、CHH、HMH、BDT、TH、CZH、ZHDH)。LCBD 置换检验表明,南漪湖(P=0.003)和青岚湖(P=0.015)统计概率达到显著水平,鄱阳湖(P=0.051)接近显著。Pearson 相关分析结果表明,LCBD 值与湖泊面积(P=0.7884)、平均水深(P=0.2074)和湖泊中蚌类物种数(P=0.3597)三者相关性均不显著。
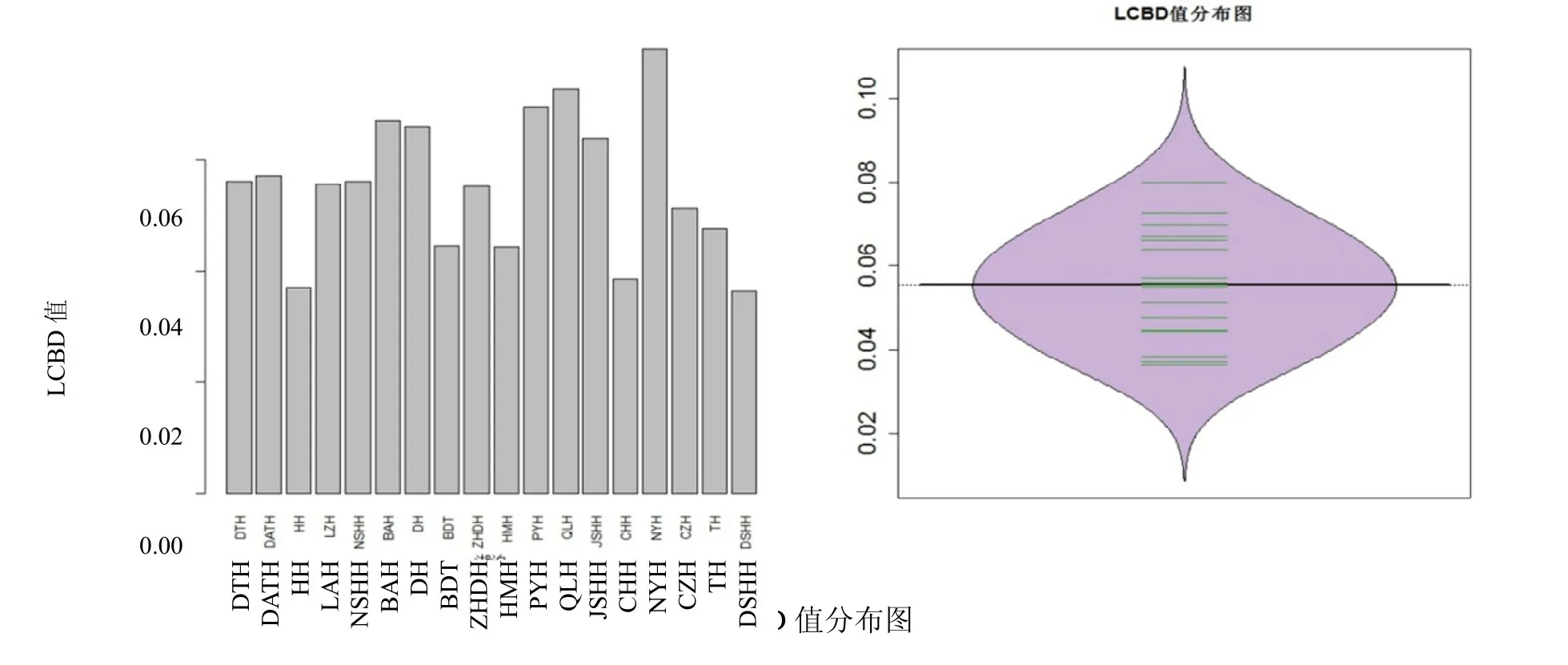
图3 LCBD 值分布图 Fig.3 The value and distribution of LCBD
2.2 周转(βsim)和嵌套(βnes)成分
根据Baselga 的分解方法,多位点计算结果表明,长江中下游湖泊蚌类总β 多样性(βJAC)为0.9238,周转成分(βJTU)为0.7488,嵌套成分(βJNE)为0.1750,总β 多样性主要由物种在空间上的周转形成,比例为81.06%。
配对方法的计算结果表明,不同湖泊对间蚌类总β 多样性βjac最大值为0.9365,最小值为0.3077,平均值为0.6829。周转成分βjtu最大值为0.8,最小值为0,平均值为0.3661。βjtu在βjac中所占比例平均为53.61%,变化范围从0 到100%。嵌套成分βjne最大值为0.8958,最小值为0,平均值为0.3168。βjne在βjac中所占比例平均为46.39%,变化范围从0 到100%(见图4)。βjtu和βjne最小值都为0,说明湖泊对之间β 多样性存在完全的嵌套和完全周转的形式。其中湖泊DTH 与DATH、HH、LZH、NSHH、BAH、BDT、HMH、JSHH、CHH、CZH 之间,湖泊DATH 与HH、PYH、CHH、TH、DSHH 之间,湖泊LZH 与HMH、TH 之间,湖泊NSHH 与PYH、CZH 之间,湖泊BAH 与BDT、HMH、TH 之间,湖泊PYH 与QLH、JSHH、CZH 之间,湖泊HH 与BAH、BDT 和TH、ZHDH 与PYH、HMH 与TH、TH 与DSHH 之间βjtu值为0,即相应各对间完全嵌套。相反,湖泊DATH 与NSHH、DH 之间,湖泊NSHH 与DH、BDT 与CZH、CHH 与DSHH 之间βjne值为0,即各湖泊对之间为完全周转(湖泊符号名称见表1)。
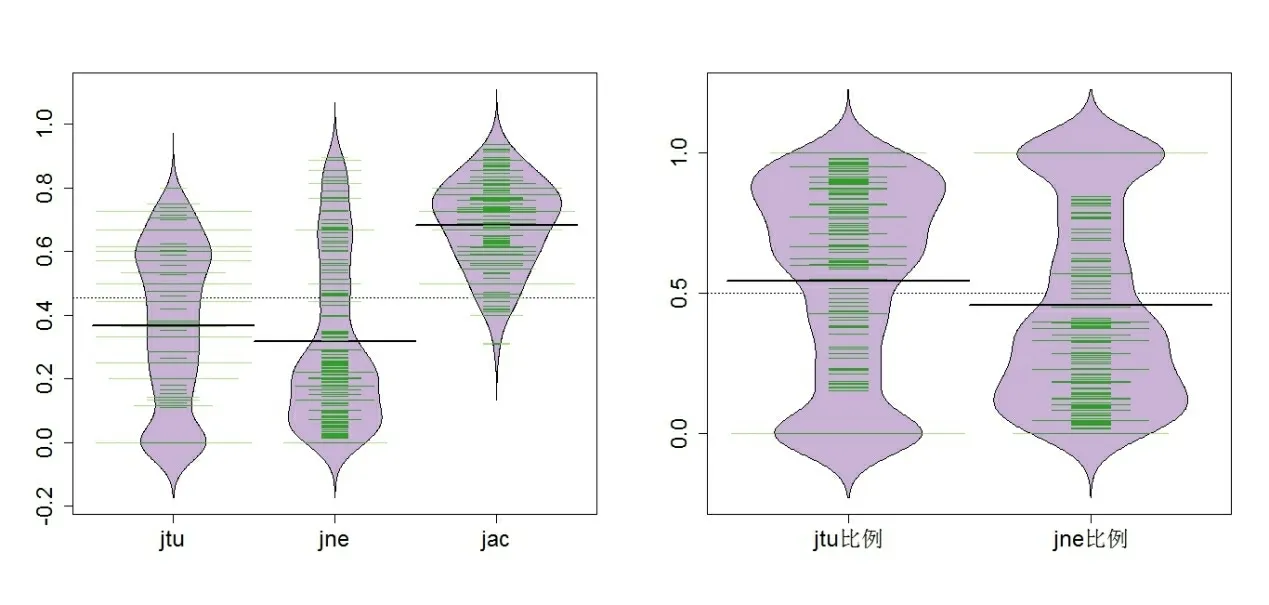
图4 βjtu 和βjne 及其在βjac 中所占比例分布 Fig.4 The distribution of βjtu , βjne and their proportion in βjac
根据配对计算的βjtu、βjne和βjac的值分别进行非度量多维尺度分析(NMDS)和聚类分析(UPGMA 法)。其中,利用βjac和βjne进行NMDS分析的应力函数值较小,分别为0.1208 和0.1229,说明相应二维点阵图排序较好,可以较好反映蚌类分布的关系。利用βjac聚类分析表明,各湖泊可在0.7 处分成3 个类群,湖泊ZHDH、JSHH、TH、QLH、DTH 和PYH 为一群,NYH 独立为一群,其余为一群(图5)。利用βjne聚类分析在0.4 处可分为2 个群。
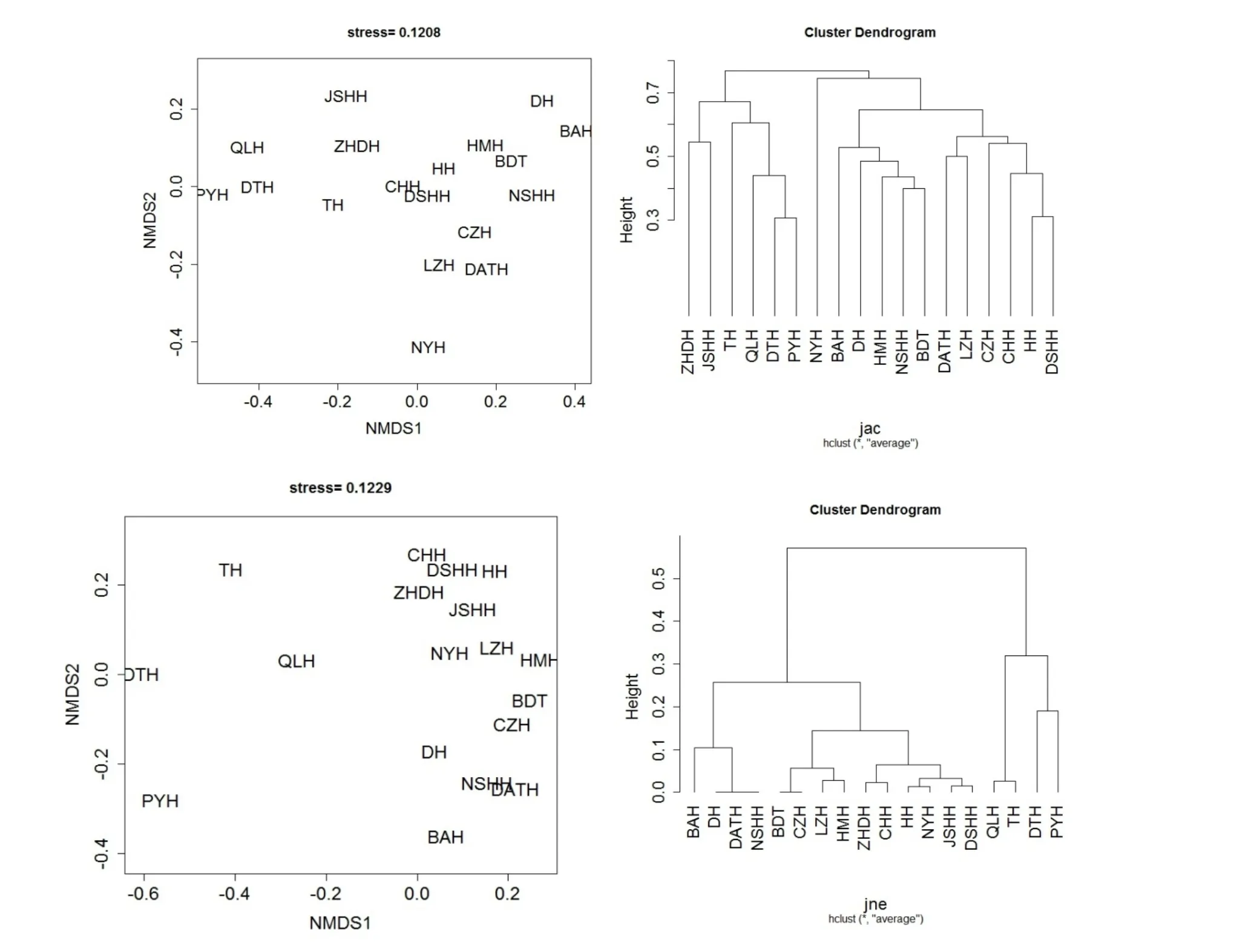
图5 βjac 和βjne NMDS 分析和聚类分析 Fig.5 The NMDS analysis and cluster analysis of βjac and βjne
3 讨论
为了在有限的资源中尽最大限度地保护生物多样性,一个地方具有很高的物种丰度(α 多样性)和(或)独特物种应该给予优先保护。在更大的区域水平上,反映各局域组成变化的β 多样性则可以提供互补的信息使保护计划能保护更多物种[36]。
长江流域分布着我国最大的淡水湖泊,其中洞庭湖和鄱阳湖物种多样性特别丰富[37],也形成和保存了众多我国特有的淡水双壳类,它是我国淡水蚌类保护和关注的热点地区[7]。在本研究β 多样性研究中,虽然洞庭湖(0.05609)和鄱阳湖(0.06967)LCBD 值都超过平均值(0.05556),但LCBD 最大值却由南漪湖(0.07974)贡献,说明长江中下游各湖泊对蚌类α 多样性与β 多样性贡献趋势并不一致。虽然南漪湖只有12 种蚌类,但却有较大的β多样性贡献度,并且显著性检验达显著水平(P=0.003)。另外,青岚湖也具有同样的特征。可能原因是LCBD值和该湖泊蚌类组成群落独特性相关,表明这两个湖泊蚌类群落组成可能与其它湖泊明显不同,分布有较少的广布种和(或)较多的狭分布种。
在不同的空间尺度,形成β 多样性的动力因素不同。在中观尺度水平,影响β 多样性的环境因素主要有气候、海拔、栖息地隔离和能量等,生物因素主要有扩散能力、洄游距离和营养地位等[38]。在流域尺度水平,相对隔离的湖泊栖息地和较弱的扩散能力可能是影响长江中下游湖泊蚌类β 多样性的主要因素[7]。由于资料中未能显示更多湖泊的环境因素信息,本研究仅仅对湖泊的平均面积和平均深度与LCBD 值相关性进行了分析,结果表明相关性均不显著。但长江中下游湖泊中蚌类物种数与该湖泊平均面积(R=0.8858,P=0.000)、平均深度(R=0.7774,P=0.000)具有显著的相关性。这也说明环境因素对β 多样性的影响与对α 多样性的影响不一致,因而在微观尺度上各种环境和生物因素对β 多样性的影响有待进一步研究。
各蚌类物种对总β 多样性的贡献从总体来看,分布湖泊数量中等左右的物种具有较大的SCBD 值(图2),因为其可以带来更多的方差(变异)。分布较少湖泊数量的物种SCBD 值较小,但普遍分布的物种SCBD 值也降低,所以长江中下游各湖泊蚌类分布范围与其SCBD 值之间呈现一种单峰的相关关系。在长江中下游湖泊蚌类中,有17 个物种分布湖泊数量为1,因此增加这些物种的分布范围可能会增加长江中下游湖泊蚌类β 多样性。相反,那些广布种的分布范围扩大,则会造成区域β 多样性降低,主要是因为造成了群落的同质化[39]。
周转和嵌套是形成β 多样性的两个对立的过程机制。在淡水生态系统中,如果一些物种具有严格限制的分布区,那么在一个流域的不同水体间更容易存在物种的周转[1]。依据Baselga 的多位点分解方法计算表明长江中下游湖泊蚌类周转成分占总β多样性的81.06%,说明该区域蚌类β 多样性主要是由于一些蚌类在不同湖泊中被其它蚌类物种所取代而导致的不同,即由不同空间的物种替换形成。利用配对计算的方法进行分析,平均周转成分比例为53.61%,也高于嵌套成分平均46.39%,这一结果与云南高原湖泊软体动物β 多样性组分构成结果一致[40]。长江中下游湖泊蚌类β 多样性周转组分比重大可能与该区域蚌类物种形成历史或者蚌类扩散能力相关,例如,蚌类成体营底栖生物,运动能力弱。蚌类幼虫的扩散能力主要和其寄生的鱼类活动范围或洄游的距离长短有关,如果阻隔了鱼类江湖洄游的路线,将极大影响蚌类的扩散和分布[41]。长江中下游湖泊有较多的特有蚌类物种,加上相对隔离的湖泊环境,蚌类在湖泊之间缺乏有效的扩散,因而只保护其中部分水域对于保护整个流域蚌类多样性有可能是不够的。
在配对计算的β 多样性方法分析结果中,虽然平均周转组分占优势,但也有一定比重的嵌套组分(平均46.39%),也存在完全的嵌套格局(即βjtu=0,βjne=βjac),这主要和蚌类物种丰富的鄱阳湖、洞庭湖、太湖和青岚湖等湖泊有关。例如,大通湖等10 个湖泊中的蚌类在洞庭湖中都有分布,军山湖等7 个湖泊中的蚌类在鄱阳湖都有分布,前者蚌类组成是后者的一个子集。利用βjac和βjne进行NMDS 分析也表明这几个湖泊排序距离较近(图5)。以嵌套为主导的β 多样性则需要优先保护物种多样性丰富度高的湖泊,因此洞庭湖和鄱阳湖等湖泊仍是长江中下游蚌类保护的重点湖泊。 通过对长江中下游湖泊淡水蚌类β 多样性进行分析,结果表明各湖泊α 多样性与β 多样性贡献趋势并不一致,因此在制定蚌类保护计划时,有必要考虑利用β 多样性提供有效的补充信息。根据β 多样性中周转和嵌套组分的构成情况,应该考虑保护重点区域和多区域保护相结合的方法。本次研究利用公开发表的文献资料初步对长江中下游湖泊蚌类空间维度β 多样性进行了分析,未能反映出长江中下游湖泊蚌类在时间维度上β 多样性的变化。Angeler 在一个长期的监测项目中对瑞典26 个湖泊沿岸带无脊椎动物群落β 多样性进行了研究,发现在总体β 多样性未发生显著变化的情况下,其周转成分增加而嵌套成分下降[42]。因此,我们将来对长江中下游各湖泊(局域)蚌类的β 多样性贡献从时间、空间两个维度都进行分析,可为长江中下游湖泊蚌类保护提供更全面的信息。
