混菌发酵酸面团对全麦面包风味与烘焙特性的影响
2020-06-06邹奇波杨文丹张宾乐周黎源黄金鑫陈军民黄卫宁
邹奇波 - 程 新 陈 诚, , 杨文丹 - 张宾乐 -周黎源 - 黄金鑫 - 陈军民 - 黄卫宁, -,
(1. 张家港福吉佳食品股份有限公司暨江南大学福临门烘焙研究所,江苏 无锡 214122;2. 江南大学食品科学与技术国家重点实验室,江苏 无锡 214122;3. 山东稻香村食品工业有限公司,山东 菏泽 274000;4. 无锡麦吉贝可生物食品有限公司,江苏 无锡 214131)
全麦粉含有较多的维生素、矿物质、纤维(如非淀粉多糖,包括阿拉伯木聚糖)、抗氧化剂和有益于人体健康的酚类化合物[1],近年来在烘焙行业的应用研究备受关注。但全麦面粉中的麸皮单独使用会对面团和面包特性产生负面影响[2],其中主要原因在于麸皮成分破坏面筋网络,导致面团持气能力较差,降低了其在发酵和烘烤过程中保持面包结构的功能[3]。同时纤维与其他聚合物对水分的竞争导致面筋蛋白和淀粉的水化不足,破坏了面团的黏弹性[4]。在现代烘焙技术中,利用酸面团发酵技术[5]能改善面包的感官品质、风味、营养价值[6-7],延长了产品保质期[8]。
现代酸面团发酵技术在全麦面团中应用具有明显优势,其部分乳酸菌发酵产生的胞外多糖可提高面团的吸水性,从而改善面团的网络结构和面包的烘焙品质及老化特性[9]。杨文丹[10] 40-41采用马克斯克鲁维酵母分泌的多种纤维素水解酶水解面团中的麦麸,可有效改善全麦面包的烘焙特性。Tang等[11-12]研究表明,高产EPS的融合魏斯氏菌可以改善面团持水性,从而改善面包面团品质。蒋慧等[13]研究发现融合魏斯氏菌和生香酵母混菌发酵可改善荞麦馒头的品质,但是产胞外多糖乳酸菌和马克斯克鲁维酵母混菌发酵在全麦面包体系中的应用尚未见报道。
试验拟采用课题组分离自酒曲的产胞外多糖食窦魏斯氏菌T5(WeissellacibariaT5)和马克斯克鲁维酵母ATCC36534(KluyveromycesmarxianusATCC36534)为研究对象,通过对比单菌发酵与混菌发酵的区别,研究酸面团发酵过程中的生化变化及其对面包面团的影响,评估混菌发酵全麦酸面团对面包品质的影响,以期为工业化开发天然、营养的高膳食纤维面包提供一定的理论支持与技术指导。
1 材料与方法
1.1 材料与试剂
全麦粉:山东省食安心安生态农场;
黄油:中粮东海粮油工业(张家港)有限公司;
即发性活性干酵母:法国乐斯福(上海)有限公司;
高筋粉:鹏泰(秦皇岛)面粉有限公司;
食窦魏斯氏菌T5:分离自酒鬼酒曲;
马克斯克鲁维酵母ATCC36534:上海一研生物科技有限公司;
YM肉汤培养基、MRS肉汤培养基:杭州百思生物技术有限公司;
对硝基苯基-β-D-吡喃葡萄糖苷、对硝基苯纤维二糖苷、对硝基苯:分析纯,阿拉丁试剂(上海)有限公司;
木糖、羧甲基纤维素钠(CMC-Na)、氯化钠、间苯三酚、琼脂粉、冰乙酸、苯酚、浓硫酸、三氯乙酸、无水乙醇:分析纯,国药集团化学试剂有限公司。
1.2 仪器与设备
恒温恒湿培养箱:SPX-150C型,上海逊博实业有限公司医疗设备厂;
醒发箱:SPC-40SP型,新麦机械(无锡)有限公司;
冷冻离心机:1850R型,湖南湘仪实验室仪器开发有限公司;
切片机:SM-302型,新麦机械(无锡)有限公司;
搅拌机:TDL-5SM-25型,新麦机械(无锡)有限公司;
烤箱:SM-503型,新麦机械(无锡)有限公司;
全温振荡培养箱:HZL-F160型,太仓市强乐实验设备有限公司;
pH计:FE-20型:梅特勒—托利多国际贸易(上海)有限公司;
紫外分光光度计:TU-1810型,北京普析通用仪器有限责任公司;
全质构分析仪:CT3型,美国Brookfield公司;
超快速气相色谱电子鼻:HeraclesⅡ型,上海瑞玢贸易公司。
1.3 方法
1.3.1 全麦酸面团的制备
(1) 采用食窦魏斯氏菌单菌发酵的全麦酸面团(WCF):将食窦魏斯氏菌接入MRS培养基中,活化2代后(37 ℃培养24 h为活化一代),离心(6 000 r/min,5 min),洗涤2次后得到T5菌泥。按照DY值(面团与面粉质量之比乘以100)为200制作100 g全麦酸面团WCF,接种量7.7 lg (CFU/g),添加5 g蔗糖后混合均匀,置于恒温培养箱(30 ℃)中发酵24 h。
(2) 采用马克斯克鲁维酵母单菌发酵的全麦酸面团(KMF):将马克斯克鲁维酵母接入至YM肉汤培养基中进行活化,后续步骤同1.3.1(1)。
(3) 采用混菌发酵的全麦酸面团(MBF):将经活化的培养基体积比为1∶50的食窦魏斯氏菌和马克斯克鲁维酵母接种制作混菌发酵酸面团MBF,其接种量7.7 lg (CFU/g)、蔗糖添加量10%(以全麦粉为基础)、DY值为200的条件接种制作酸面团,发酵方式同1.3.1(1)。
1.3.2 发酵过程中全麦酸面团的菌株生长曲线和酸度变化
(1) 全麦酸面团菌落总数:通过每隔2 h监测酸面团中食窦魏斯氏菌和马克斯克鲁维酵母的菌落数反映菌株生长特性及其相互作用。将10 g样品放入90 mL无菌生理盐水中摇匀,稀释至合适的浓度,分别取样品液100 μL涂布接种于MRS和YM固体培养基,于37 ℃与30 ℃的环境下培养48 h后,进行菌落计数,样品重复计数3次。
(2) 全麦酸面团pH和TTA:通过每隔2 h监测酸面团发酵过程pH值和总可滴定酸度(TTA)的变化来反映食窦魏斯氏菌和马克斯克鲁维酵母的酸化作用。其测试方法参照AACC方法(2000)02-52。
1.3.3 纤维素酶(内、外切葡聚糖酶,β-葡萄糖苷酶)、木聚糖酶活力测定
(1) 粗酶液的提取:参照杨文丹[10]11-14的方法。
(2) 内切葡聚糖酶活力测定:参照Dhillon等[14]的方法。以2.0~12.0 μmol/mL葡萄糖绘制标准曲线计算内切葡聚糖酶酶活。
(3) 外切葡聚糖酶活力测定:参照杨文丹[10]11-12的方法。以0.01~0.06 μmol/mL对硝基苯酚溶液绘制标准曲线计算外切葡聚糖酶酶活。
(4)β-葡萄糖苷酶活力测定:结合钱超的方法[15]。使用0.01~0.06 μmol/mL对硝基苯酚为标样绘制标准曲线计算酶活。
(5) 木聚糖酶活力测定:结合Chapla等[16]的方法测定木聚糖酶活力。采用0.2~0.8 μmol/mL木糖绘制标准曲线计算木聚糖酶酶活。
1.3.4 可溶/不溶阿拉伯木聚糖测定 采用Zhao等[17]的方法将1.0 g全麦酸面团与20 mL去离子水震荡提取可溶性阿拉伯木聚糖(WEAX)。使用Kiszonas等[18]的方法对总阿拉伯木聚糖含量(AX)进行测定。采用Douglas[19]的方法绘制AX标准曲线。水不溶性阿拉伯木聚糖(WUAX)为AX与WEAX的差值。
1.3.5 胞外多糖(EPS)产量测定 采用Tang等[12]的方法对EPS进行提取后,利用苯酚硫酸法[20]测定溶液中多糖的含量。
1.3.6 全麦面包的制备 表1展示了普通小麦面包(KBB)、对照全麦面包(QBB)、采用食窦魏斯氏菌T5发酵的全麦酸面团面包(WCB)、采用马克斯克鲁维酵母发酵的全麦酸面团面包(KMB)、采用混菌发酵的全麦酸面团面包(MBB)的制作配方。
不同面包的制作工艺为:将除黄油外的原料投入搅面缸,提前将盐和糖溶解到水中,慢速混合搅拌3 min再快速搅拌1 min 形成少量面筋。加入黄油,慢速混合3 min,高速搅拌2 min 至面筋完全扩展完成。松弛面团5 min后分割成90 g/个的面团,面团搓圆后继续松弛10 min,放进温度为38 ℃,RH为85%的恒温恒湿箱发酵120 min,取出后在上火210 ℃、底火170 ℃的烤箱中焙烤22 min,取出后脱模冷却2 h得到成品面包。
1.3.7 全麦酸面团面包烘焙和风味特性
(1) 比容测定:待面包冷却后,参照杨文丹[10]14-16的方法测定,重复3次取平均值。
(2) 全质构测定:面包冷却1 h后,使用切片机切成10 mm的均匀薄片,全质构测定参数设置:P/36,压缩程度40%,测试间隔时间1 s。每组面包重复3次试验取平均值[10]14-16。
(3) 超快速电子鼻HeraclesⅡ分析:取全麦面包样品3 g,置于20 mL顶空瓶中,于50 ℃烘箱中30 min。采用Yang等[21]的方法分析全麦面包样品,得到电子鼻数据输出雷达指纹图谱和PCA分析图。
(4) 感官品评得分:选取24位经过感官品评训练的人员通过9分嗜好评分法对面包外观、色泽、口感、风味及整体可接受度五维度进行感官品评分析[22]。
1.4 数据处理
采用Excel 2016、Origin 9.1和SPSS来进行数据分析、绘图、显著性分析和主成分分析(PCA),数据、图表中均采用字母上标来表示显著性分析结果,字母不同表示具有显著性差异(P<0.05)。
2 结果与分析
2.1 全麦酸面团发酵过程中菌株的生长情况及酸度变化
2.1.1 菌落总数 图1展示了酸面团发酵过程中食窦魏斯氏菌和马克斯克鲁维酵母的生长曲线,结果表明在WCF中的食窦魏斯氏菌稳定期菌落数达到9.51 lg (CFU/g),而MBF中食窦魏斯氏菌T5的菌落数达到9.61 lg (CFU/g),原因在于马克斯克鲁维酵母在酸面团发酵代谢过程中会产生食窦魏斯氏菌生长代谢所需的维生素与氨基酸等营养物质,从而促进其更好的生长[23]。相比于食窦魏斯氏菌,马克斯克鲁维酵母的生长稳定性较强,KMF和MBF组中马克斯克鲁维酵母稳定期的菌落数分别为8.21,8.09 lg (CFU/g),原因在于食窦魏斯氏菌产生的低pH值环境或食窦魏斯氏菌与马克斯克鲁维酵母竞争碳源,从而一定程度上抑制马克斯克鲁维酵母的生长[24]。
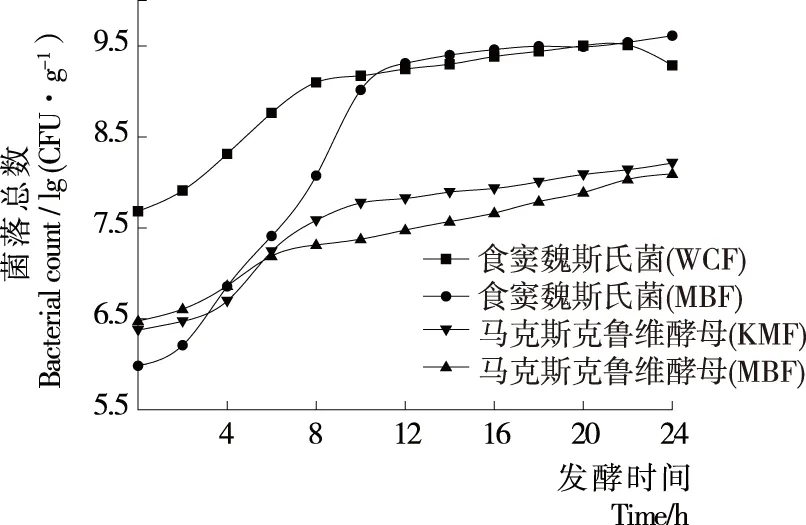
图1 全麦酸面团发酵过程中食窦魏斯氏菌和马克斯克鲁维酵母的生长曲线
Figure 1 The growth curve ofWeissellacibariaandKluyveromycesmarxianusduring the fermentation of whole wheat sourdough
2.1.2 pH和TTA 由图2可知,全麦酸面团的pH值随发酵时间延长呈下降趋势,TTA与pH值呈反相关。比较混菌发酵和单菌发酵组结果,混菌发酵的酸面团酸化速率介于两种单菌发酵之间。随着食窦魏斯氏菌的引入,相比于仅含马克斯克鲁维酵母的酸面团,混菌组的酸化速率显著上升。低pH环境有利于激活全麦粉中的内源性蛋白酶和淀粉酶活性以及β-葡萄糖苷酶活力,有利于游离氨基酸和可溶性糖的代谢,改善酵母产气从而改善全麦面团的面筋结构。
TTA是评价面团酸化动力学的有力指标,酸面团发

图2 WCF、KMF和MBF全麦酸面团的pH与TTAFigure 2 The pH and TTA of WCF, KMF and MBF
酵过程中有机酸的积累是造成TTA增大的主要原因之一,在最后发酵阶段TTA存在增加趋势,但体系中pH无明显变化。其原因在于全麦粉中灰分含量高,对pH具有缓冲作用,从而维持体系的pH稳定。
2.2 纤维素酶(内、外切葡聚糖酶,β-葡萄糖苷酶)、木聚糖酶活力
图3展示了全麦酸面团在单菌与混菌发酵作用24 h后纤维素酶(内切葡聚糖酶、外切葡聚糖酶、β-葡萄糖苷酶)及木聚糖酶活力。WCF组发酵后的内、外切葡聚糖酶活力最低,而KMF与MBF中的内、外切葡聚糖酶活力无显著性差异。MBF组发酵24 h后内切葡聚糖酶活力达到7.33 U/g,外切葡聚糖酶活力达到1.35 U/g。图3(c)表明KMB组中β-葡萄糖苷酶活力最高,24 h后达到14.41 U/g。其原因主要为两方面:① 由于内、外葡聚糖酶活力较高,能为β-葡萄糖苷酶提供充分的底物;② 由于马克斯克鲁维酵母自身能分泌较多的胞外β-葡萄糖苷酶[10] 3-4。而MBF组中β-葡萄糖苷酶活力为略低于KMB组,为13.59 U/g,其原因为酸面团发酵后期的乳酸菌酸性较强,从而抑制酵母菌生长。
图3(d)展示了不同类型酸面团发酵24 h的木聚糖酶活力,MBF>KMB>WCF,且混菌发酵与单菌发酵具有显著性差异。Dhillon等[14]和Hansen等[25]研究结果表明麦麸本身含有一定量的木聚糖酶。木聚糖酶活力在发酵前期由于内源性酶被激活会有明显的上升,后期活力有所下降,可能与底物限制以及产物的反馈抑制有关[26]。而混菌发酵24 h后木聚糖酶活力达到2.81 U/g,表明可能混菌发酵过程中反馈抑制作用较弱,更有利于木聚糖酶保持活力。
2.3 全麦酸面团发酵过程中可溶性/不溶性阿拉伯木聚糖组成变化
阿拉伯木聚糖是全麦粉中重要的组分,未发酵前的AX的初始平均含量为8.95%,其中WEAX含量为0.77%。经过24 h酸面团发酵过程,体系中的WEAX含量都有所增加(见图4)。其中KMF和MBF中的WEAX含量增加明显,MBF组和KMF组WEAX含量分别为1.89,1.75 g/100 g,WEAX含量分别增加了145.5%,127.2%,结合木聚糖酶酶活的结果说明,体系中的WEAX含量与木聚糖酶活力直接呈正相关[如图3(d)],其原因在于木聚糖酶通过切割阿拉伯木聚糖的主链,降低其聚合度,生成小分子片段木寡糖、木聚糖以及游离木糖等可溶性阿拉伯木聚糖[27]。而WCF组发酵24 h后WEAX含量最低,为1.20 g/100 g,仅增加55.8%,其原因可能是乳酸菌发酵24 h后过度产酸,过低的pH抑制了大部分木聚糖酶酶活,降低AX降解。此外,无论在单菌与混菌酸面团的发酵,WUAX含量均出现下降。
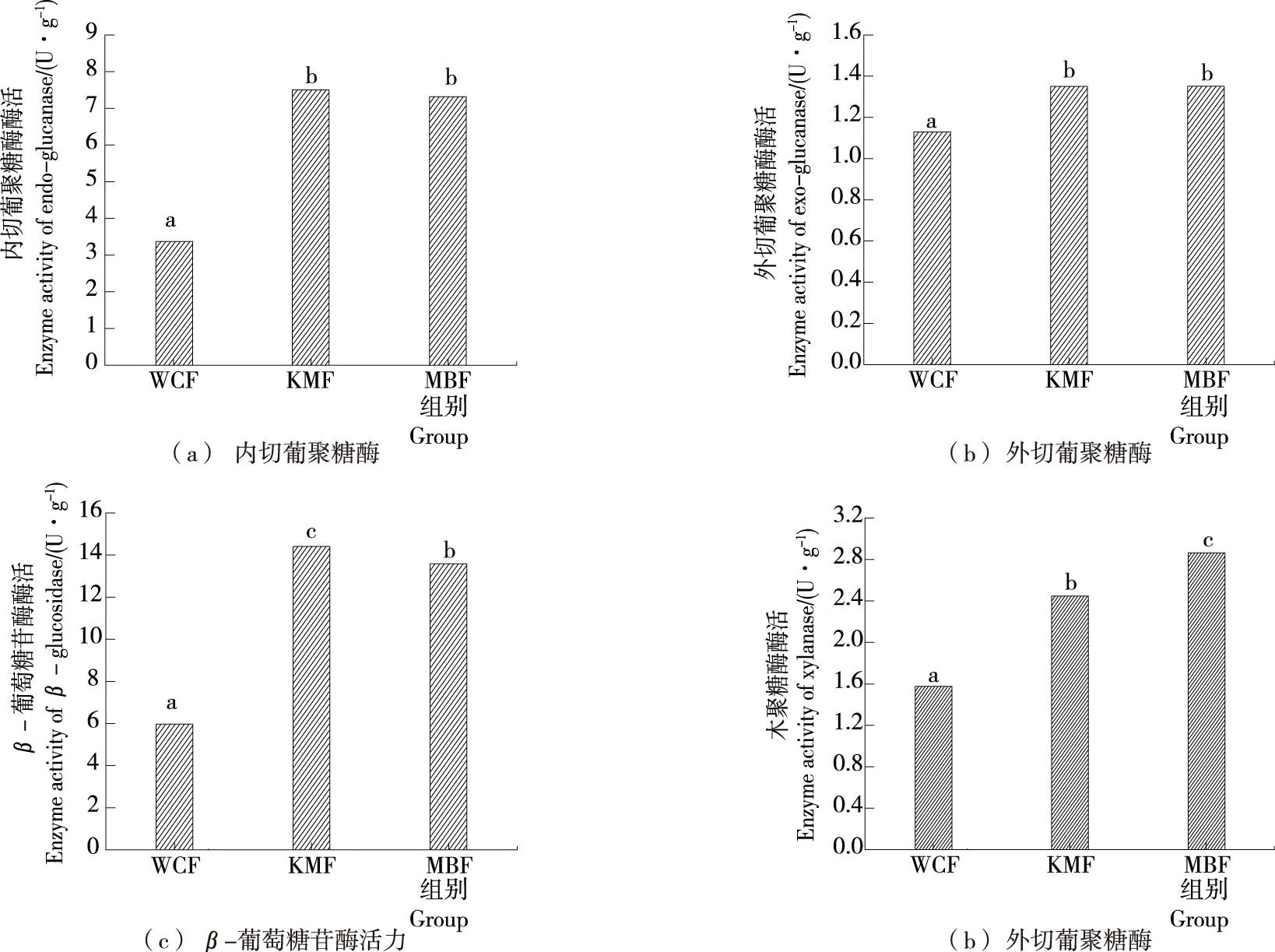
图3 含有不同菌种的全麦酸面团发酵24 h后各纤维素酶酶活活力Figure 3 The cellulase activity of whole wheat sourdough containing different strains after 24 h fermentation
2.4 发酵全麦酸面团EPS产量
图5展示了利用苯酚硫酸法测定的EPS含量标准曲线方程为y=0.043 6x-0.044 7 (R2=0.999 1)。图6结果表明,经过24 h发酵后,WCF组与MBF组的EPS产量分别为6.57,7.54 g/kg,远大于KMF的EPS产量(2.51 g/kg)。食窦魏斯氏菌以蔗糖作为糖基供体,通过葡糖基转移酶合成葡聚糖,从而增加EPS产量[28]。相比于WCF组,MBF组的EPS产量较高结合菌落数结果,其原因可能在于马克斯克鲁维酵母在酸面团发酵过程产生的氨基酸、维生素等营养物质可供食窦魏斯氏菌更好的生产[24],进而促进全麦酸面团体系中的EPS生成。

图4 全麦酸面团发酵过程中可溶性/不可溶性阿拉伯木聚糖含量变化
Figure 4 Content changes WEAX and WUAX during fermented process of whole wheat sourdoughs
2.5 全麦酸面团面包烘焙和风味特性
2.5.1 比容 表2中的面包比容结果显示,QBB面包比容最小,为5.20 mL/g。因为全麦粉的加入减少了高筋粉比例,面筋含量大幅度减少,并且全麦粉中的膳食纤维破坏了面团面筋网络结构连续性,影响面包中气泡的稳定性,导致其持气性差[3]。相比QBB,添加全麦酸面团后面筋网络结构得到强化,全麦面包比容得到明显提升,KMB组比容增加18.84%,原因在于酵母产气促进面团膨松,很大程度上可归因于木聚糖酶对AX组分的降解。Courtin等[29]认为木聚糖酶促进WUAX溶解,释放WEAX,从而降低WUAX对面团面筋体系的消极影响,进而增加面筋得率。MBB组的比容最大,达到6.47 mL/g,除WEAX和WUAX含量变化作用外,食窦魏斯氏菌的高产EPS特性在全麦面包体系中起到了关键作用,EPS作为生物亲水胶体能促进其与谷蛋白之间的交联作用,增强面团稳定性和持气能力,从而改善全麦面包的烘焙特性[30]。

图5 EPS含量(葡萄糖)标准曲线(苯酚硫酸法)
Figure 5 Standard curve of EPS (glucose) production (Phenol-sulfuric acid method)

图6 全麦酸面团发酵24 h后EPS含量
Figure 6 The EPS content in whole wheat sourdoughs after 24 h fermentation
2.5.2 全质构分析 QBB面包芯硬度高达392.0 g,而酸面团发酵全麦面包的面包芯结构更加柔软,这归因于马克斯克鲁维酵母发酵全麦酸面团中的纤维素酶、木聚糖酶酶活的提升及食窦魏斯氏菌产生的EPS。木聚糖酶可以改善全麦面包的质构特性,原因在于木聚糖酶水解阿拉伯木聚糖后,其持水能力降低,释放的水分促进了面团体系中面筋蛋白的水合,酶解后产生的小分子WEAX以及少量寡糖、单糖可为酵母提供碳源用于面团发酵产气,增大了面包体积,导致硬度显著降低,弹性显著增加[31]。而乳酸菌发酵产生的EPS能明显改善面包的质构特性的原因在于其能提高面团乳化特性、增加面筋网络交联和增强面团持水性[30]。

表2 不同类型面包烘焙特性分析†Table 2 The baking characteristics analysis of different types of bread
2.5.3 HeraclesⅡ电子鼻香气成分特征分析 图7外围表示14种检测器下特征香气成分的信号强度。相比于QBB组,检测器1、4、5、9以及11的检测信号在KMB和MBB组中明显增强,而这些检测器都能检测出的特征性香味是芳香族化合物[21]。此外,检测器4、5以及检测器9还能检测出有机酸酯、萜烯类、醇类以及脂肪烃类。说明在混菌发酵全麦酸面团过程中,部分羰基类以及烷烃类的风味化合物消失或转化成风味更强的醇类、酸类和酯类等风味化合物。WCB中由于乳酸菌发酵产生的乙酸是引起差异的主要原因,KMB和MBB中由于酵母菌或酵母与乳酸菌的协同作用,合成了更多的醇类、酯类以及芳香族化合物。由于酯类物质阈值低,在相同含量下具有更浓郁的酒香、果香和花香[13]。KMB和MBB组的风味化合物相对含量明显高于QBB和WCB组,说明含有马克斯克鲁维酵母的混菌发酵全麦面包的风味强度优于单一乳酸菌发酵的,而KMB与MBB的风味强度差异不大。
在图8中,样品间的DI值为98,说明各样品间风味存在明显差异。PCA总体贡献率达到99.7%,说明该PCA分析方法能够准确描述样品之间的差异。从图8中结果表明,马克斯克鲁维酵母的引入是造成各样品间风味差异的主要原因,主要差距体现于PC1上;而引入食窦魏斯氏菌对全麦基质的面包产品的风味改善则是主要体现在PC2上。此外,可看出WCB组和QBB组在电子鼻检测器下风味化合物含量无明显差异。而KMB组与MBB组比较相近,与风味化合物的雷达图结果一致。因此,不同酸面团发酵形式对全麦酸面团风味有较大影响,混菌发酵组香气明显优于单一乳酸菌发酵组。
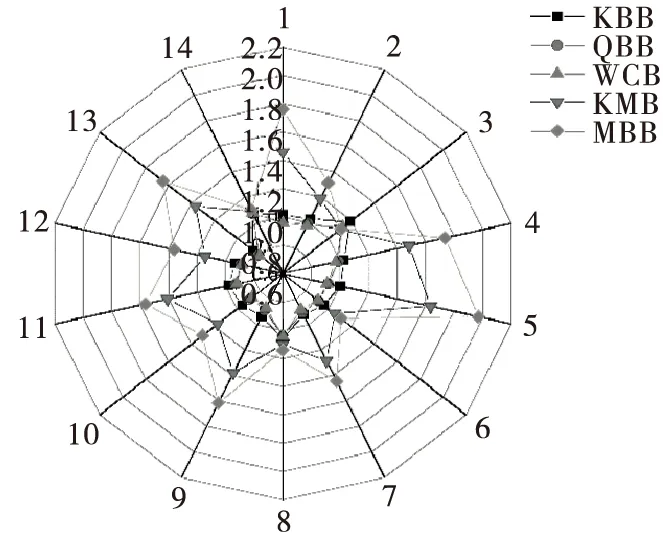
图7 全麦酸面团面包样品的Heracles Ⅱ雷达图
Figure 7 The heracles Ⅱ radar image of whole wheat sourdough breads
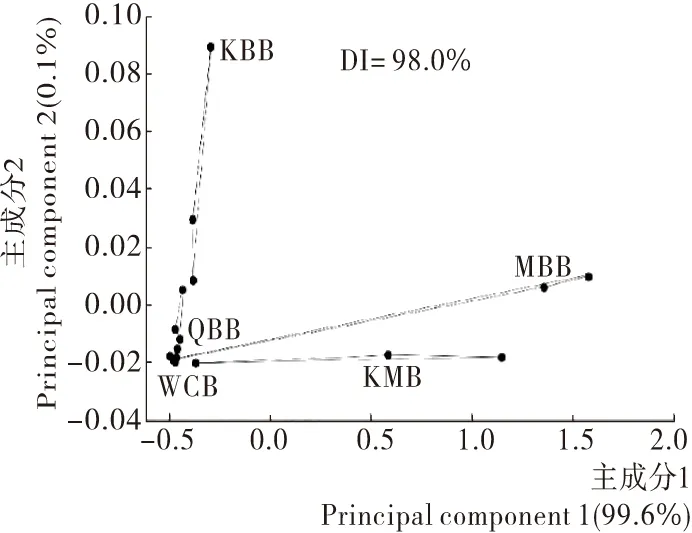
图8 全麦酸面团样品的Heracles Ⅱ 主成分分析图
Figure 8 The heracles Ⅱ principal component analysis (PCA) of whole wheat sourdoughs
2.5.4 感官品评 图9表明,全麦粉经过酸面团发酵处理后,其外观、口感、色泽以及风味得到显著性改善,乳酸菌与酵母菌在酸面团发酵过程中的独特的生长代谢特性起到了关键性的作用,主要是乳酸菌代谢产生的EPS以及酵母代谢激活的各种水解酶对面筋网络的改善作用。对比QBB和KMB组发现,酵母菌发酵可改善全麦面包的风味,但对其外观无显著影响;对比QBB和WCB发现,仅引入乳酸菌单菌发酵全麦酸面团能明显改善全麦面包的外观、色泽、口感,但风味改善效果不明显,同时终产品的酸味较强。在整体接受度维度评分中,得分最高的组别为MBB,与电子鼻和质构分析测定结果相符,经过食窦魏斯氏菌和马克斯克鲁维酵母的发酵协同作用后,所得的全麦面包香气更加浓郁,质构柔软,同时具有更加饱满的外观。

图9 不同组别的全麦面包的感官品评
Figure 9 Sensory evaluation result of different whole wheat breads
3 结论
马克斯克鲁维酵母的存在能够促进食窦魏斯氏菌的生长,二者在全麦酸面团体系中具有共生作用。从烘焙特性角度来看,混菌发酵中的马克斯克鲁维酵母通过提升体系中的纤维素酶酶活促进体系中阿拉伯木聚糖的降解,体系中的WXAX含量增加至1.89 g/100 g,WUAX含量降低,进而保护面筋结构的稳定性和全麦面包的烘焙特性;而混菌发酵中的食窦魏斯氏菌提高了全麦酸面团中的EPS含量,从而改善面包比容和质构。从风味角度来看,混菌发酵中的马克斯克鲁维酵母大大改善了全麦面包的风味强度,同时感官评定中取得最高的整体可接受度。结果表明,具有协同作用的混菌发酵更有利于改善全麦面包烘焙与风味特性。
采用混菌发酵制作的全麦面包品质和风味较好,后续将研究荞麦、藜麦等其他基质中不同前体物质经过混菌发酵后对面包风味的影响,为系统性开发天然营养的面包提供技术支撑。
