miR-124 靶向OSX 对牙周膜干细胞成骨分化能力的调控作用*
2020-05-27王献刚冯保静
王献刚 冯保静
牙周炎是一种由牙菌斑等多种因素引起的慢性炎症性疾病,可导致牙周组织进行性丧失,从而导致牙齿松动和脱落。近年,组织工程再生和牙周膜干细胞成为牙周疾病治疗的前景和研究热点。PDLSCs 是来源于牙周支持组织中的一群未分化细胞,具有高增殖率和多向分化潜能,在相应诱导条件下可向成骨细胞、成软骨细胞、脂肪细胞以及内皮细胞等分化,被认为是牙周组织工程中牙周改建、修复的理想材料和种子细胞[1]。牙周病的理想治疗目标是实现牙周支持组织再生,而PDLSCs在牙周骨质缺损修复及组织工程再生中发挥重要作用[2]。成骨分化是一个高度协调的过程,涉及Runx2 和OSX 等多种转录因子,Runx2 是间充质干细胞向成骨前体细胞分化的关键,OSX 是成骨前体细胞向功能性成骨细胞分化的关键[3-5]。OSX缺失的小鼠出生后立即死亡,并显示完全没有骨形成[6,7]。OSX 作为成骨细胞的特异性转录因子可通过上调成骨细胞特异性分化标志物的表达,包括碱性磷酸酶(ALP)、骨钙素(OCN)、骨连接蛋白(ON)和骨桥蛋白(OPN),从而影响骨形成[8]。在PDLSCs中OSX 基因可调控Wnt/β-catenin 信号通路从而影响牙周膜干细胞的成骨向分化[9]。因此,研究OSX 对牙周膜干细胞成骨分化能力的调控机制对牙周组织工程应用具有重要意义。
MicroRNAs (miRNAs)是非编码家族的单链RNA,长度约18~25 个核苷酸,可与靶基因mRNA的3’UTRs 相结合,促进靶mRNA 降解或抑制靶mRNA 翻译,进而介导基因抑制[10]。MicroRNA在调控间充质干细胞增殖、成骨分化能力方面具有重要作用,研究显示miR-124 在强直性脊柱炎成骨细胞中增加GSK-3β 进而下调Wnt/β-catenin入核而抑制成骨分化过程[11]。研究表明,miR-124可通过靶向调控骨髓间充质干(BMSCs)细胞中的OSX 进而抑制成骨分化[12]。然而miR-124 是否影响牙周膜干细胞成骨分化能力及其作用机制尚不清楚。本实验将通过研究miR-124 对牙周膜干细胞成骨分化的影响及其机制,以期为牙周膜干细胞在牙周组织修复再生应用中提供理论依据。
1.材料和方法
1.1 实验材料 细胞孵育箱(Heraeus,德国);酶联仪(BIO-TEK,美国);离心机(Eppendorf,德国);胎牛血清(Gibco,美国);胰蛋白酶、PBS、α-MEM 培养基(Hyclone,美国);倒置相差显微镜(OLYMPUS,日本);miR124mimics、miR124 inhibitor、si-RNA(OSX)(锐博生物公司,中国);RIPA 蛋白裂解液、双萤光素酶报告基因检测试剂盒(碧云天,中国);OSX 抗体(货号ab94744,Abcam,美国);Lipofectamine2000(Invitrogen,美国)。
1.2 细胞培养及成骨诱导 经安阳市第六人民医院伦理委员会同意,收集18~25 岁口腔科患者者即刻拔除的第三磨牙,要求牙齿健康完整无龋坏及牙周病,超净台内充分冲洗牙齿,使用刀片刮取牙齿根中1/3 处牙周膜,加入I 型胶原酶(3mg/L)放入细胞培养箱消化40min,每10 分钟摇匀一次,使用含10%FBS 的培养液终止消化,去除上清后用含20%胎牛血清的α-MEM 培养液后重悬后接种于培养板中,于细胞培养箱内培养,每2 天换一次液,待细胞长至80%融合率时,挑选单克隆纯化PDLSCs 后用于后续实验。
取P3 代PDLSCs,消化重悬后接种于培养板中,分为对照组(非成骨诱导组)和成骨诱导组,当PDLSCs 达80%左右的融合率,更换成骨诱导液(10%FBS,10 mm β-甘油磷酸钠、50ug/uL 维生素C 和10 nmol/L 地塞米松),成骨诱导2 周后检测miR124 及成骨基因的表达。
1.3 q-RCR 检测目的基因mRNA 的表达取P3 代PDLSCs 接种于培养板中,分为miR124 mimics、miR124 inhibitor、miR124 对照组(miR124-NC)、control 组(图2)及si-NC、si-OSX、miR-124 inhibitor+OSX 沉默组(miR-124+si-OSX)和miR-124inhibitor+OSX 沉默对照组(miR-124+si-NC)(图4),参照Lipo-2000 说明书对人PDLSCs 进行转染,转染48h 后提取miR124 mimics、inhibitor 和对照组细胞的总RNA,按照PrimeScriptTMRT reagent Kit 说明书逆转录合成cDNA。
1.4 Western blotting 检测PDLSCs 中OSX蛋白的表达 取P3 代PDLSCs 接种于6 孔培养板中,分为分为miR124 mimics、miR124 inhibitor、miR124-NC、control 组(图2)及si-NC、si-OSX、miR-124+si-OSX 和miR-124+si-NC 组(图4),提取各组细胞总蛋白后95℃变性5min,配制10%SDS-PAGE 胶,110mV 电压下电泳分离细胞蛋白、70mV 电压转膜、5%脱脂奶粉TBST 封闭后,加入OSX(1∶1000 稀释)抗体、β-actin(1∶10 000稀释)抗体,于冷房过夜孵育。TBST 轻柔漂洗3次,每次10 分钟,加入二抗后孵育1h,TBST 轻柔漂洗3 次,每次10 分钟,ECL 化学发光后采用Image J 软件分析结果,以OSX/β-actin 条带灰度值表示OSX 的相对表达水平。
1.5 茜素红实验检测成骨分化能力 取P3 代PDLSCs 接种于培养板中,分为miR124 mimics、miR124 inhibitor、miR124-NC、control 组(图2)及si-NC、si-OSX、miR-124+si-OSX 和miR-124+si-NC 组(图4),参照Lipo-2000 说明书分别对PDLSCs 进行转染,当细胞达到80%以上的融合率;更换成骨诱导液(10%FBS,10 mm β-甘油磷酸钠、50ug/uL 维生素C 和10 nm 地塞米松),成骨诱导2 周后,4%多聚甲醛加入培养板,固定细胞20min,PBS 漂洗2~3 次,加入茜素红染液孵育15min,PBS 充分洗去茜素红染液,显微镜观察、拍照茜素红染色结果。
1.6 双荧光素酶报告基因实验生物信息学分析预测miR-124 的靶基因,结果发现OSX 为miR-124 的靶基因,为确定miR-124 靶向结合于OSX的3’UTR 区,将OSXmRNA 突变型(OSX-MUT)和野生型(OSX-WT)的3’UTR 区合成,并连接到psiCHENK-2 载体质粒。在PDLSCs 中共转染miR124-NC、miR124 mimics 和OSX 质粒,分组为OSX-WT+miR124 NC、OSX-MUT+miR124 NC、OSX-WT+miR124 mimics、OSX-MUT+miR124 mimics。转染后冰上裂解细胞,检测荧光强度并进行分析。
1.7 统计学分析 采用SPSS 19.0 统计软件对数据进行统计学分析,计量资料以均数±标准差表示,组间比较采用t 检验或单因素方差分析,P<0.05 表示差异有统计学意义。
2.结果
2.1 PDLSCs 成骨分化前后miR-124 和OSX的表达 qPCR 检测PDLSCs 分化后成骨基因和miR-124 的表达变化,结果显示,PDLSCs 成骨分化后miR-124 的表达明显下调(图1),而OSX、OCN、Runx2、ALP 的表达水平显著升高(图1),差异具有统计学意义(P<0.05),结果与文献报道一致[12]。
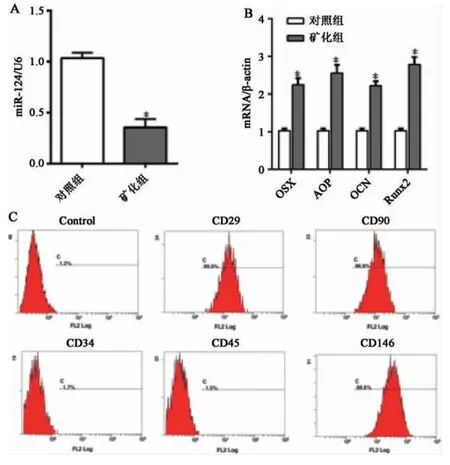
图1 牙周膜干细胞成骨诱导后miR-124 和成骨相关基因的表达
2.2 miR-124 对PDLSCs 成骨分化能力的调控PDLSCs 转染miR124 mimics 和miR124 inhibitor,q-PCR 验证转染效果,结果显示miR124 inhibitor组中miR-124 的表达下调,而miR124 mimics 组中miR-124 的表达明显上调,差异具有统计学意义(P<0.05)(图2)。PDLSCs 转染miR124 后成骨诱导2 周,茜素红结果显示miR-124 inhibitor 组中钙化结节明显增多,而miR-124 mimics 组则显著减少;q-PCR 结果显示miR-124 inhibitor 组中OSX、ALP、Runx2、OCNmRNA 及OSX 蛋白的表达明显上调,而miR-124 mimics 组则结果相反;差异具有统计学意义(P<0.05)(图2)。
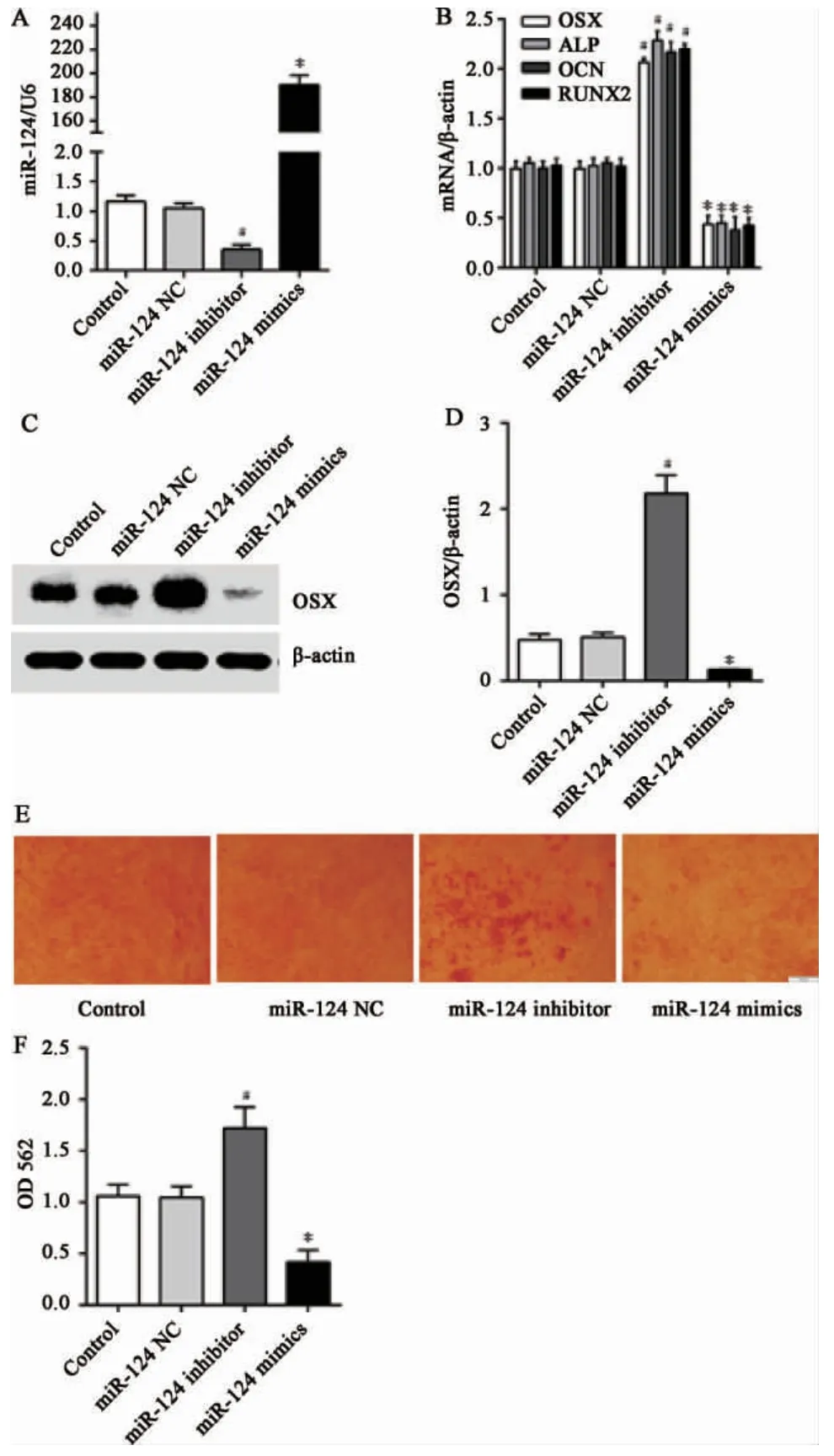
图2 miR-124 对PDLSCs 的成骨分化能力的影响
2.3 miR-124 的靶基因为OSX 生物信息学分析预测发现OSX 为miR-124 的潜在靶基因,图2 结果显示miR-124 mimics 组中OSX 的表达明显低于对照组,说明OSX 可被miR-124 负向调控。采用双荧光素酶报告基因实验证实OSX 是miR-124 的靶基因,检测结果显示,OSX-WT+miR-124 组的荧光比值低于OSX-WT+miR-NC组(图3),差异具有统计学意义(P<0.05)。OSXMUT+miR-124 组和OSX-MUT+miR-NC 组的荧光比值分别为0.952±0.047 和1.0±0.073,差异无统计学意义(P>0.05)。表明miR-124 可与OSX 基因3’-UTR 位点结合,miR-124 与OSX存在直接负向调控关系。

图3 双荧光素酶报告验证OSX 为miR-124 靶基因
2.4 沉默OSX 能逆转miR-124 inhibitor 对PDLSCs 成骨向分化的促进作用 为进一步研究miR-124 和OSX 的关系,在牙周膜干细胞中同时转染siRNA-OSX 和miR-124inhibitor,分析对PDLSCs 的成骨分化能力的影响。结果显示,共转染siRNA-OSX 和miR-124inhibitor 可逆转miR-124 inhibitor 对OSX 的表达促进作用(图4)。茜素红染色结果可见siRNA-OSX 和miR-124 inhibitor组(miR-124+si-OSX)与miR-124 inhibitor 组相比,miR-124+si-OSX 组中钙化结节数明显减少(图4)。说明沉默OSX 能够逆转miR-124 inhibitor对PDLSCs 的成骨分化的促进作用。
3.讨论
牙周炎发病后导致牙周支持组织渐行性破坏,牙槽骨的修复弱于吸收,进而导致牙槽骨的高度降低[13]。牙周炎治疗不仅要控制牙周炎症,还要促进牙周支持组织的修复再生。因此,明确牙槽骨形成的细胞分子机制对于牙槽骨修复和牙周组织再生的至关重要。2004 年,Seo 等[1]人类牙齿牙周膜中培养出PDLSCs。PDLSCs 具有较强的自我更新能力和成骨、成脂等多向分化潜能,使PDLSCs 在牙周组织工程再生领域中拥有广阔的应用前景[14]。本实验探索了miR-124 对牙周膜干细胞成骨向分化能力中的影响,研究结果显示PDLSCs 在成骨分化后miR-124 的表达水平呈现下调趋势,而OSX的表达则显著升高。通过inhibitor 抑制miR-124的表达可上调牙周膜干细胞的成骨分化能力,并可促进OSX mRNA 和蛋白的表达,miR-124 与OSX的表达呈负相关,结果与文献报道相一致[12]。OSX作为一种调控细胞成骨分化的转录因子,可特异性调控多种成骨相关基因的转录[7]。OSX 的表达是成骨前体细胞向成骨细胞分化的必要条件,OSX 基因的转录表达可调控成骨细胞的晚期分化,当OSX 基因缺失时,则成骨细胞分化则基本完全被抑制[6]。此外,成牙本质细胞中条件性敲除OSX的小鼠可表现出短而薄的牙根表型,说明在牙根发育过程中OSX 发挥了重要作用[15]。机制研究表明,OSX 表达的调控网络可以通过调控上皮BMP 信号通路、间质Runx2 表达和细胞磷酸化水平来调控[16]。综上,OSX 对细胞成骨分化能力具有重要的调控作用,因此研究OSX 在PDLSCs 中的调控机制对牙周支持组织再生具有重要意义。为了进一步了解OSX 在PDLSCs 的作用机制。本研究通过生信分析发现OSX 为miR-124 的潜在作用靶点,双荧光素酶实验验证结果显示与OSX 突变组相比,OSX-WT+miR-124 组的相对荧光素酶活性显著抑制,而OSX-WT+miR-NC 组的无显著变化,表明miR-124 可与OSX 的3’-UTR 位点结合,两者存在直接负向调控关系。有研究表明miR-124可靶向结合OSX 进而抑制骨髓间充质干细胞体外成骨分化能力[12],与本实验结果相一致。MicroRNAs(miRNAs)是一种在结构上不同于双链小干扰RNA(siRNA)的非编码RNA,作用机制也与siRNA 介导的mRNA 基因沉默有所区别,miRNA-RISC结合复合体对靶基因降解效能主要取决于其与mRNA 转录本3’-UTR 序列互补的程度。本实验中miR-124 与OSX 的3’-UTR 序列高度互补,并证实miR-124 与OSX 互补结合后导致OSX mRNA 发生降解,抑制其翻译效率,进而影响OSX 蛋白的表达。本研究结果显示miR-124 可能是一个新的调控成骨分化的重要路径,参与了OSX 基因介导的PDLSCs 成骨分化进程,这对牙周组织损伤修复时的骨改建具有重要意义。
本研究采用双荧光素酶报告系统证实了miR-124 结合与OSXmRNA3’-UTR 区,即OSX 为miR-124 的靶基因,最后miR-124 inhibitor 和OSX-siRNA 共转染实验进一步证实了沉默OSX可逆转miR-124 inhibitor 对PDLSCs 成骨分化能力的促进作用。综上所述,miR-124 通过靶向负调控OSX 对牙周膜干细胞的成骨分化产生抑制作用,提示miR-124 在牙周膜干细胞和组织损伤修复过程中发挥重要作用。
