多房棘球绦虫重组抗原蛋白14-3-3对小鼠继发性泡球蚴病的预防与治疗效果#
2020-05-25李润乐
魏 威,王 蕾,周 培,张 颖,李润乐&,汤 锋
(1.青海大学高原医学研究中心,青海省高原医学应用基础重点实验室,青海 西宁 810001;2.青海大学基础医学部,青海 西宁 810001;3.青海大学研究生院,青海 西宁 810016)
14-3-3蛋白是众多真核生物中高度保守并广泛表达的酸性蛋白家族成员[1,2]。在不同的生长阶段都可检测到稳定存在的14-3-3蛋白并发现其在多房棘球绦虫(E.multilocularis)的增殖、分化等过程中发挥重要的调控作用[3-5]。本课题评价了重组抗原蛋白14-3-3的免疫原性和其在多房棘球绦虫原头节继发性感染小鼠模型中的预防及治疗效果。
1.材料与方法
1.1 实验材料
1.1.1 主要试剂
pCzn1载体(南京,钟鼎生物);TOP 10感受态细胞、BL-21感受态细胞(北京,天根生化);质粒提取试剂盒(上海,生工);镍离子亲和层析介质(南京,金斯瑞);Elisa抗体(英国,Abcam);细胞因子IL-4、IFN-γ检测试剂盒(瑞典,Mabtech)。
1.1.2 实验动物
36只5~7周龄雄性BALB/c小鼠(北京,斯贝福),动物合格证号:SCXK9(京)2019-0010;对实验动物的日常护理及实验操作均遵循NIH实验室动物护理和使用指南。
动物使用方案通过了青海大学动物伦理与实验委员会的审核(QHDX 2018 09)。
1.2 实验方法
1.2.1 pCzn1-14-3-3重组质粒的构建
从GenBank中获得14-3-3的氨基酸序列(GenBank ID:CDS36536.1),经OptimumTMCodon软件优化得到适合原核表达的碱基序列,引入NdeI和XbaI限制性内切酶位点并克隆入载体pCzn1中,通过NdeI和XbaI双酶切验证质粒是否构建成功。
1.2.2 重组蛋白14-3-3的表达及纯化
重组蛋白14-3-3的诱导表达参照Li等[6]的方法。经IPTG诱导并确定重组蛋白表达部位后,通过镍离子亲和层析法纯化14-3-3蛋白,以40、35、30、25、20 mM咪唑分别洗脱杂蛋白,之后使用200 mM咪唑洗脱目的蛋白并收集。
1.2.3 小鼠免疫与感染
将12只小鼠分为两组,分别用50 μg/只的14-3-3抗原蛋白和PBS免疫(4次,间隔1周),第一次免疫加入等量的完全弗氏佐剂,而第二、三次免疫加等量的不完全弗氏佐剂,末次免疫不加佐剂。最后一次免疫完成后(7天)取血清(4000rpm/min)并保存在-80 ℃冰箱。
原头节的分离:参照文献方法[6]。脱颈处死多房棘球蚴感染的保种沙鼠,从沙鼠腹腔分离囊泡后剪碎并依次经45目和150目筛网分离得到多房棘球蚴原头节。
预防评价模型的建立:参照文献方法[7]。每组6只的两组小鼠免疫接种14-3-3抗原蛋白(实验组)或PBS(对照组),共进行4次(间隔一周)。最后一次免疫接种后两周时向腹腔内注射原头节(1000个),感染4个月后采集小鼠血清并处死小鼠,对其囊泡进行计数与称重。
治疗评价模型的建立:参照文献方法[6]。向两组小鼠(每组6只)腹腔注射原头节(1000个),感染4个月后每隔1个月腹腔注射含等量完全弗氏佐剂的重组14-3-3蛋白(50μg/只)或PBS(50μg/只),共进行4次。末次免疫后两周,采集血清后处死小鼠并对其囊泡进行计数与称重。
1.2.4 特异性抗体检测

1.2.5 细胞因子检测
血清IFN-γ、IL-4的分泌水平通过ELISA法测定,具体的操作按试剂盒说明进行。
1.3 统计学方法
2.结果
2.1 重组质粒14-3-3的构建及酶切鉴定结果
pCzn1-14-3-3重组质粒的构建图谱如图1所示;重组质粒酶切鉴定显示目的条带与14-3-3标准序列长度一致,表明pCzn1-14-3-3目的序列已成功插入载体中,如图2所示。

图1 pCzn1-14-3-3重组质粒的构建图

Line1:酶切前质粒;Line2:酶切后质粒.酶切位点:NdeI-XhoI
2.2 重组蛋白14-3-3的表达与纯化结果
分别收集14-3-3蛋白原核表达中的菌液及沉淀进行凝胶电泳,重组蛋白在28 KD处形成明显条带,并且主要以包涵体的形式存在于沉淀中(图3A)。14-3-3蛋白包涵体经过变、复性处理后,通过Ni-IDA亲和层析法纯化14-3-3蛋白,以40 mM的咪唑洗去杂蛋白后得到的蛋白的纯度最高(图3B)。

A.原核表达的12%SDS-PAGE分析图;B.重组蛋白14-3-3的纯化图
2.3 重组14-3-3蛋白免疫后特异性抗体和细胞因子检测结果
采用间接ELISA法评价14-3-3对小鼠的特异性抗体诱导能力。与PBS免疫组相比,14-3-3免疫能显著增加特定的免疫球蛋白的分泌水平。小鼠血清抗14-3-3的抗体中除IgE与PBS免疫小鼠相比无差异(P>0.05)外,IgG1、IgG2a、IgG、IgM和IgA的抗体水平与PBS免疫小鼠相比都明显升高(P<0.05)(表1);通过ELISA法检测了小鼠血清中IFN-γ和IL-4的水平。与PBS免疫小鼠和正常对照组(NC)相比,14-3-3免疫小鼠IFN-γ细胞因子浓度显著升高(vs.PBS:P=0.004;vs.control:P=0.001;F=54.731);IL-4的分泌水平(vs.PBS:P=0.764;vs.control:P=0.370;F=1.715)在免疫后无明显升高(表2)。

表1 重组14-3-3蛋白免疫后小鼠特异性抗体检测结果
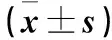
表2 重组14-3-3蛋白免疫后小鼠细胞因子检测结果
2.4 重组14-3-3蛋白的预防作用
14-3-3的预防效果通过检测囊泡的数量和重量来评估。14-3-3与PBS免疫小鼠的囊泡数量、重量差异显著,分别减少了59.84±15.09%(P<0.05)和83.28±18.01%(P<0.001)(图4、表3)。IgG1、IgG2a、IgG、IgM、IgA都较PBS组显著升高(P<0.05);IgE的水平与对照组相比无明显增高(P>0.05)(表4);小鼠血清的IFN-γ(vs.PBS:P=0.000;vs.control:P=0.000;F=621.968)、IL-4(vs.PBS:P=0.002;vs.control:P=0.000;F=19.430)水平显著增高(表5),提示重组14-3-3蛋白对E.multilocularis所致的泡球蚴病(Alveolar Echinococcosis,AE)有较好的预防作用。

图4 囊泡的大小和数量
表3 预防性免疫后重组14-3-3蛋白对囊泡数量和囊泡重量的影响结果

表4 预防性免疫后的特异性抗体水平

表5 预防性免疫后IFN-γ和IL-4水平
2.5 重组14-3-3蛋白的治疗作用
14-3-3的治疗效果的评估方法与预防性研究方法相同。14-3-3与PBS治疗小鼠的效果在囊泡数量上没有差异(P>0.05),在囊泡重量上减少了(P<0.05。72.28±11.47%。图5,表6)。在治疗组中,血清IgG1、IgG2a、IgG、IgM、IgA显著升高(P<0.05);IgE的水平与对照组相比无明显增高(P>0.05。表7);小鼠血清IFN-γ(vs.PBS:P=0.000;vs.control:P=0.000;F=129.171)、IL-4(vs.PBS:P=0.003;vs.control:P=0.000;F=57.053)水平显著增高(表8),提示重组14-3-3蛋白对E.multilocularis所致的AE有良好的治疗作用。

图5 囊泡的大小和数量

表6 治疗性免疫后重组14-3-3蛋白对囊泡数量和囊泡重量的影响结果
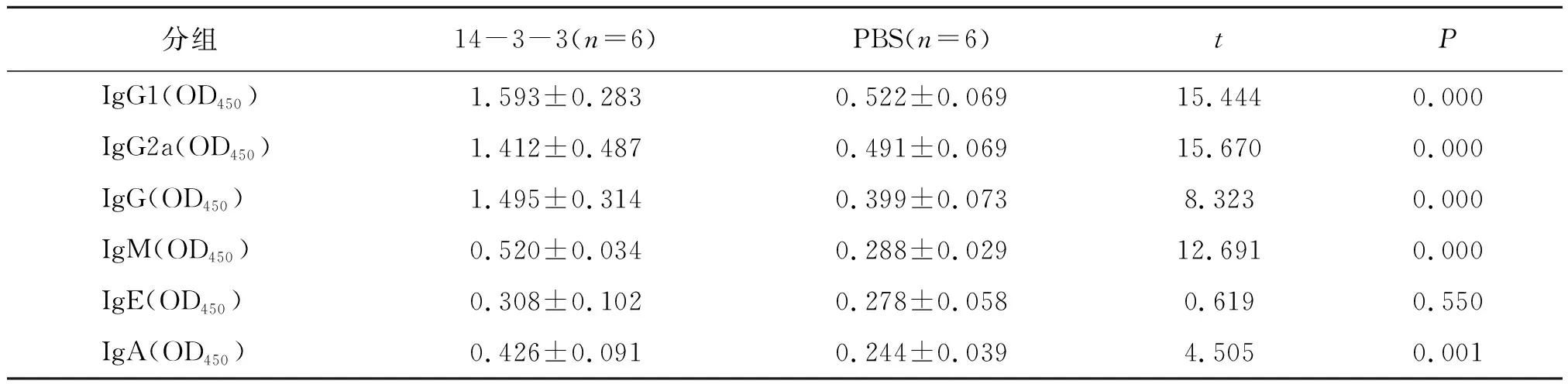
表7 治疗性免疫后的特异性抗体水平

表8 治疗性免疫后的IFN-γ和IL-4水平
3.讨论
AE被认为是对人类最为致命的寄生虫病之一,这种被忽视的人畜共患病呈现发展态势,在不受干预的情况下十年内超过九成的患者死亡[8]。近20年来,该病在全球的流行规模显著扩大。欧洲流行范围向东部和北部扩展,亚洲多地也是受影响区域,日本、中国等地都有相关报道[9]。对于这种有着严重危害的疾病,阿苯达唑或甲苯咪唑化疗是AE的药物治疗方法,但此药物长期使用所产生的副作用影响了疗效。
14-3-3蛋白与寄生虫的生长有着密切关系,所以针对多种病原生物14-3-3蛋白的研究已经展开了。Li等[10]发现在犬新孢子虫中该蛋白可以激活多种信号通路而引起机体免疫系统的反应,提示其可作为一种新的抗新孢子虫病候选疫苗。李文桂等[11]以该蛋白作为疫苗对其是否可以在小鼠中产生有效的保护作用进行研究,实验结果显示14-3-3可抑制棘球蚴生长,产生了较强的保护力。为探究14-3-3蛋白在人类和非人灵长类动物中的保护作用,Lampe等[12]以明矾作为佐剂的重组抗原14-3-3免疫恒河猴,证实了这种蛋白的免疫保护在非人灵长类动物中也是安全有效的。在多房棘球绦虫中,14-3-3表现出了更加多样的功能,它的多个亚型在生活史的各个阶段可根据不同需求差异性表达[13,14]。14-3-3蛋白参与了寄生虫关键的细胞生命活动,进一步探究该蛋白对宿主免疫系统和寄生虫生长发育的影响将有助于发现它的作用机制。
寄生虫感染通常导致白细胞中嗜酸性粒细胞的增加,这一群体的细胞具有限制寄生虫生长的功能,发挥作用的抗体包括IgG、IgM、IgE和IgA。14-3-3蛋白诱导了高水平的特异性免疫球蛋白,但血清特异性IgE抗体没有显著增加,这可能与寄生虫特异性T细胞产生的IL-4和IFN-γ的相对量增加有关,而其中IFN-γ可抑制寄生虫抗原所诱导的IgE产生[15,16]。在AE的感染过程中,免疫应答可分为三个明显的阶段:早期为Th1/Th2混合型免疫应答,中期则是以Th2型免疫应答为主导,在感染的最后阶段出现T细胞耗竭状态[17]。在本研究中,我们发现14-3-3蛋白作为预防和治疗的疫苗,可以使感染多房棘球绦虫小鼠的IFN-γ和IL-4显著增加,引发了Th1-Th2混合免疫反应。在感染多房棘球绦虫后,小鼠肝脏NK细胞数量减少,这可能通过降低IFN-γ的分泌而限制其细胞毒活性,并使得被感染小鼠肝脏的免疫耐受能力受到影响[18],而14-3-3蛋白使多房棘球蚴感染后小鼠体内的寄生虫囊泡重量明显降低的结果或许与免疫佐剂所诱导的IFN-γ水平显著增高有关。
本课题成功构建了多房棘球绦虫抗原14-3-3蛋白的原核表达系统并分离纯化了高纯度的14-3-3重组蛋白,对其所产生的免疫预防和治疗作用进行了研究,这对后续探究14-3-3蛋白在寄生虫感染等领域中的生物学作用提供了依据。
