α-淀粉酶对猪粪厌氧发酵产甲烷的影响及动力学分析
2020-05-14肖艳春
陈 彪, 黄 婧, 肖艳春,2
(1.福建省农业科学院农业工程技术研究所, 福建 福州 350003; 2.福建技术师范学院 近海流域环境测控治理福建省高校重点实验室, 福建 福清 350300)
利用厌氧发酵技术处理畜禽粪便是目前世界范围内公认的最为有效的处理方式之一。厌氧发酵技术可将生物质等转化为沼气,在获取能源物质的同时,实现减少环境污染的目的[1-2]。2015年我国畜禽粪便资源总量为20.08亿t,畜禽粪便资源总量约合2.87亿t干物质,具有963.77亿m3沼气潜力[3]。提升厌氧发酵产沼潜力可有效促进厌氧发酵技术在污水处理领域的工程实践应用与发展,尤其对规模化养殖污水的治理及其资源化利用具有十分重要的意义。
厌氧发酵过程可分为有机物的水解、产酸和产甲烷3个阶段,而发酵原料的不同性质决定了限制阶段的不同。猪粪中主要大分子复杂有机物是淀粉、蛋白质和纤维素。因此,猪粪的水解阶段是厌氧发酵过程的限速步骤。已有相关研究报道,利用水解酶提高发酵原料水解产物的浓度。水解酶是一类能够催化促进有机物水解反应的酶,包括淀粉酶、纤维素酶、蛋白酶、脂肪酶等。Yang[4]等在厌氧发酵条件下添加淀粉酶和蛋白酶提高剩余污泥的生物水解效率,淀粉酶处理比蛋白酶处理具有更高的水解效率,淀粉酶处理比对照组显著提高了54.24%;许美兰[5]等利用α-淀粉酶和蛋白酶构成的复合酶对猪粪进行处理,可溶性化学需氧量(SCOD)提高106.9%;王婷婷[6]等通过添加酶对酿酒废糟进行干式厌氧发酵,提高甲烷产率达36.8%。目前关于水解酶对猪粪厌氧发酵产甲烷的影响的动力学方面的研究鲜见报道。本研究采用升流式厌氧污泥床(UASB)工艺,以猪粪为发酵原料,应用Gompertz和一级动力学模型解析外源α-淀粉酶对厌氧发酵产甲烷的影响效应,以期提升发酵原料的产甲烷潜力,提高厌氧发酵技术在养殖污水处理领域的工程实践应用效应。
1 材料与方法
1.1 材料和装置
鲜猪粪取自福建某养殖公司,接种污泥取自常年正常运行的沼气发酵装置,猪粪及接种污泥成分组成见表1。α-淀粉酶购自上海源叶生物科技有限公司,酶活≥4000 U·g-1,pH值范围为5.5~7.5,最适温度为50℃~70℃。试验采用UASB工艺,装置示意见图1。

表1 猪粪及接种污泥的成分

1.三相分离器; 2.水封瓶; 3.气体流量计; 4.出水口; 5.取样口; 6.水泵; 7.蠕动泵; 8.恒温水浴箱; 9.废水进水池图1 UASB发酵装置示意
1.2 试验方法
试验设置1个对照组(CK组)、1个空白组和4个试验组,每组设置3个重复,厌氧发酵周期为35 d。每组取猪粪1225 g,以未添加α-淀粉酶的处理为CK组,以只添加接种污泥与蒸馏水的处理为空白组,4个试验组按照猪粪的总固体(TS)重量每克分别加入α-淀粉酶(酶添加量参考相关文献[5, 7])10,20,30,40 mg,加入α-淀粉酶搅拌均匀后在50℃水浴锅中恒温震荡处理4 h,充分发挥α-淀粉酶活性,待猪粪冷却至常温后每组接入1 L的接种污泥,经调配后发酵料液总体积为5 L(不足部分用蒸馏水补足、TS约为8%),料液通过蠕动泵导入UASB发酵装置,进料结束后向发酵装置中通N210 min保持厌氧环境,调节装置温度为35℃。厌氧发酵过程中每日定时测定沼气产气量、甲烷体积分数。
猪粪及接种物的TS、挥发性固体(VS)采用差重法测定;C,H,O,N,S元素的质量分数采用元素分析仪(Vario EL III,Elementar,德国)测定;沼气日产气量使用湿式流量计(LML-1,长春汽车滤清器有限责任公司)测定;沼气中甲烷体积分数和总挥发性脂肪酸浓度采用气相色谱法测定。
1.3 数据分析
1.3.1 发酵原料理论产甲烷潜力值
目前,发酵原料理论产甲烷潜力值的计算是通过分析物料中的C,H,O,N,S等元素含量进行估算,碳水化合物、蛋白质等有机物可用CxHyOzNnSs表示,通过方程式(1)估算出发酵原料的理论产甲烷潜力值Xe[8]。该研究中的甲烷产率均是基于猪粪中挥发性固体(VS)含量计算。
(1)
式中:Xe为发酵原料的理论产甲烷潜力值,mL·g-1;x,y,z,n,s分别代表了发酵原料C,H,O,N,S元素含量与C,H,O, N,S元素相对原子质量的比值(取整数)。
1.3.2 发酵底物产甲烷动力学
应用一级动力学和Gompertz模型拟合厌氧发酵过程并计算动力学反应常数,根据决定系数R2及均方根误差RMSE对模型的拟合度进行评估。
一级动力学方程:
xt=x∞[1-exp(-kt)]
(2)
式中:X∞为整个发酵周期内猪粪甲烷产率,mL·g-1;Xt为t时刻猪粪甲烷产率,mL·g-1;k为反应速率常数,d-1。
Gompertz模型:
(3)
式中:Rm为产甲烷速率,mL·g-1·d-1;e为常数,2.718282;λ为迟滞时间,d。
1.3.3 发酵底物生物转化产甲烷效率
以不同时刻甲烷累积产量与整个发酵周期甲烷总产量的比值表示发酵过程甲烷产率比(MPR)。以厌氧发酵不同时刻实际甲烷产率与理论甲烷产率的比值表示底物的生物产甲烷效率(BDA),计算公式:
(4)
式中:Xt为t时刻实际甲烷产率,mL·g-1;Xe为理论甲烷产率,mL·g-1。
1.3.4 数据处理
采用SPSS 19.0进行数据处理及方差分析,采用Origin 9.1软件进行绘图及方程拟合。各试验组的总挥发性脂肪酸浓度、沼气产气量、甲烷体积分数取3次重复试验的平均值。
2 结果与讨论
2.1 α-淀粉酶对总挥发性脂肪酸的影响
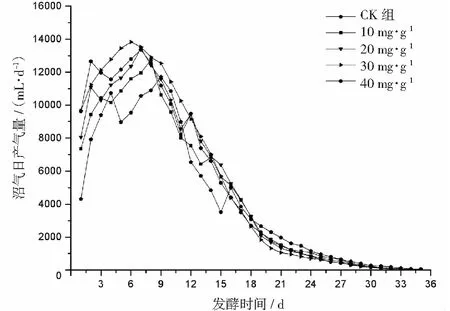
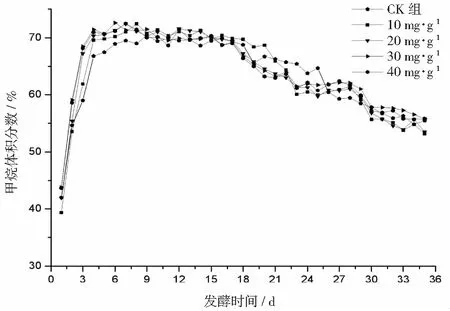
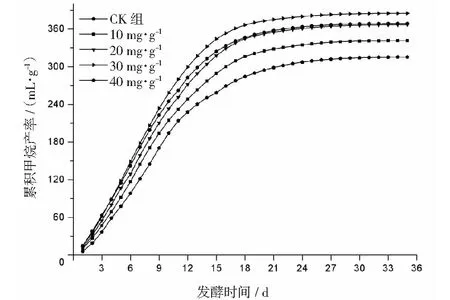
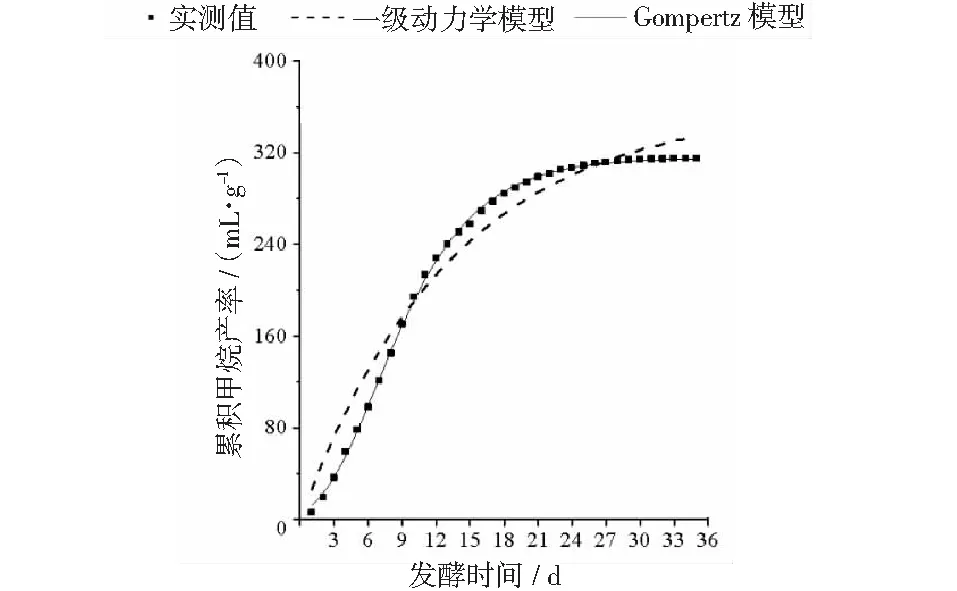
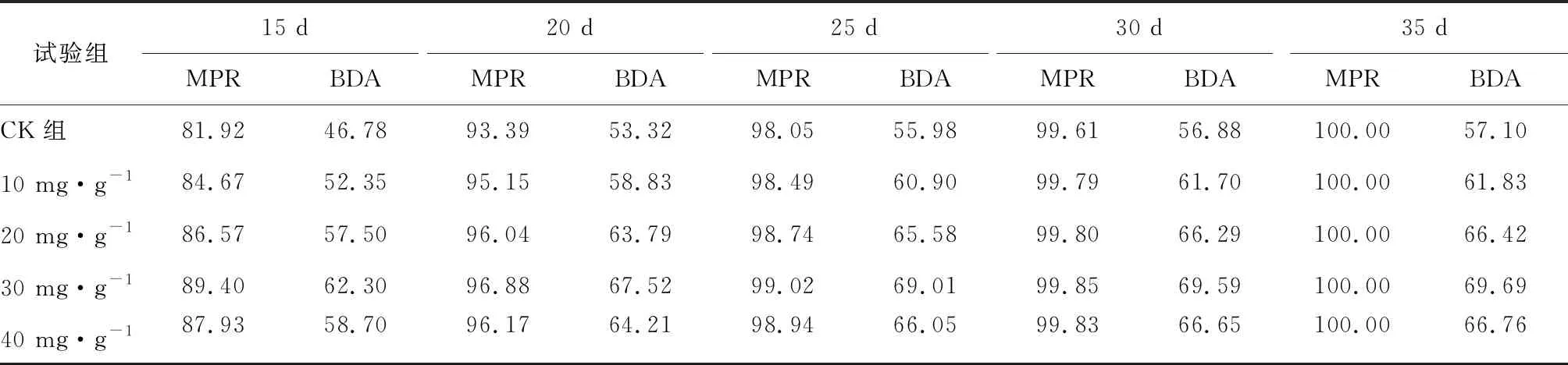
厌氧发酵过程各试验组总挥发性脂肪酸(TVFAs)的浓度变化如图2所示。由图2可见:各试验组和CK组的变化规律一致,均呈先升后降,最后趋于平稳的特征。CK组在第3天达到产酸高峰(TVFAs质量浓度为7030 mg·L-1);α-淀粉酶添加量为20,30,40 mg·g-1的试验组均比CK组提前1天达到产酸高峰,其中添加量为40 mg·g-1的产酸峰值最大(8986 mg·L-1),比CK组提高了27.8%;其次是添加量为30 mg·g-1的试验组(8308 mg·L-1),比CK组提高了18.2%。在厌氧发酵后期,各试验组进入稳定期,至发酵结束时各试验组的TVFAs累积量均低于CK组,其累积显著度为30 mg·g-1<40 mg·g-1<20 mg·g-1<10 mg·g-1 图2 总挥发性脂肪酸浓度变化曲线 2.2.1 α-淀粉酶对日产沼气量的影响 各试验组日产沼气量变化情况见图3,从图3可以看出,在厌氧发酵的进程中,沼气日产气量总体呈现先上升后下降的趋势。各试验组的产气启动期均较短,在较短时间内就达到产气高峰,这应与本试验的接种污泥取自运行良好的沼气工程厌氧发酵装置,接种污泥中的功能微生物丰富且活跃有关。CK组,10 mg·g-1,20 mg·g-1和40 mg·g-1试验组均出现了3个产气高峰,CK组产气高峰分别出现在第4,9,16天,30 mg·g-1试验组只在第6天出现1个产气高峰,峰值期比CK组提前3 d。日产气峰值最大值(13829 mL)出现在30 mg·g-1试验组,与CK组(11710 mL)相比增加了18.1%。 有研究表明[9],乙酸是沼气发酵过程中重要的中间代谢产物。由于微生物优先利用小分子糖类及淀粉类等易降解的有机物,水解作用生成的葡萄糖在厌氧条件下经糖酵解(EMP)途径生成丙酮酸,丙酮酸是合成乙酸等挥发性脂肪酸的主要中间产物,生成的甲烷三分之二来源于乙酸,试验组添加的α-淀粉酶促进了溶解性多糖浓度的增加[10],它可将淀粉类物质酶解成糊精、麦芽糖、葡萄糖等这些容易被产氢产乙酸或耗氢产乙酸细菌利用的营养底物,进而为产甲烷细菌提供了大量的乙酸盐,CO2,氢等物质,促进沼气产气量提高,因此添加了α-淀粉酶的试验组比CK组更快达到产气峰值。而后产气速率下降,出现下1个产气高峰,这是因为容易降解的底物被消耗后,沼气发酵微生物不得不利用纤维素等较难降解有机物,产气峰值不如前期。由此,α-淀粉酶的适量添加有利于产气峰值的提前到达。 图3 沼气日产气量变化曲线 2.2.2 α-淀粉酶对甲烷体积分数的影响 图4为沼气中甲烷体积分数变化曲线图,该体积分数的变化是厌氧发酵过程中不同微生物菌群之间相互作用的结果。由图4可以看出,甲烷体积分数先呈现快速增长而后趋于稳定,末期则逐步降低。在厌氧发酵的第2~3天,各试验组甲烷体积分数就已经大于50%,均进入正常产气状态。CK组的甲烷体积分数从第4天起就进入稳定期,至第26天结束,共保持了23 d,甲烷体积分数60.4%~70.4%;30 mg·g-1试验组的稳定期最长,从第3天持续到第29天,甲烷体积分数61.6%~70.6%,比CK组略有增加,试验组添加的α-淀粉酶使厌氧发酵系统中可利用的糖类物质增加,提高了系统中甲烷途径中碳物质浓度,促进最终产物甲烷体积分数的提升。 图4 沼气中甲烷体积分数变化曲线 2.2.3 α-淀粉酶对甲烷产率的影响 图5为各试验组累积甲烷产率曲线图。由图5可知,CK组的累积甲烷产率最低(315.4 mL·g-1VS)。杜连柱[11]等研究发现在35℃条件下,TS为7.5%的猪粪进行厌氧发酵时,其甲烷产率为326 mL·g-1VS,与本试验的结果较接近。随着α-淀粉酶添加量从10 mg·g-1增加到30 mg·g-1时,甲烷产率亦随之增加,在添加量为30 mg·g-1时达到最高的384.9 mL·g-1VS,比CK组提高了22.1%;而当添加量继续增至40 mg·g-1时,其对应的甲烷产率相较30 mg·g-1试验组反而降低。由此分析,与CK组相比,在适当的范围内添加α-淀粉酶可以促进发酵料液中的淀粉水解为小分子糖类,即丙酮酸的前体物质,产氢产乙酸细菌将丙酮酸转化为如乙酸、丙酸、丁酸、甲醇等挥发性有机酸,反应底物的增加会提升产甲烷菌转化速率,提高甲烷产率;但过量添加的α-淀粉酶可导致过快的有机物水解及后续过快的挥发性有机酸的产生,一旦超过产甲烷菌甲烷转化的速率,挥发性有机酸的过量积累将抑制产甲烷菌的活性,此外,底物中部分细菌的新陈代谢活动获取营养物质的速度高于酶促反应速度,导致可溶性物质浓度下降,使甲烷产率下降。试验结果表明,在猪粪厌氧发酵进程中,溶解性碳源物质的浓度决定了厌氧发酵的甲烷产率,α-淀粉酶的添加阈值为30 mg·g-1。 图5 累积甲烷产率曲线 本试验采用经典的一级动力学以及Gompertz模型对不同试验组的累积甲烷产率曲线进行拟合,具体见图6~图10。表2列出了2种模型的拟合结果,其中,产甲烷动力学参数包括动力学常数(k)、产气速率(Rm)、发酵迟滞期时长(λ)等。根据决定系数R2及均方根误差RMSE对模型拟合的表现进行评估,从拟合结果可以看出,采用Gompertz模型拟合各试验组产甲烷过程,拟合度较好,R2均大于0.998,模型拟合的累积甲烷产率最大值与实测值接近,说明该模型可以较为准确的反应各试验组的产气情况;一级动力学模型的拟合精确度较低,拟合结果与实测值存在偏离,因此,该模型不适用于本研究中产甲烷过程拟合。 从Gompertz模型的拟合结果可以看出,CK组的Rm最慢(26.18 mL·g-1d-1),30 mg·g-1试验组的Rm最快(32.63 mL·g-1d-1),比CK组提高24.6%。随着α-淀粉酶的增加λ随之缩短,CK组λ最长(2.29 d),40 mg·g-1试验组λ较短(1.08 d);30 mg·g-1试验组λ迟滞期次之(1.52 d)。这是由于添加的酶增加了发酵底物中小分子糖类的浓度和可吸附位点数量,促进产甲烷微生物菌群繁殖生长,进而提升甲烷产率。试验结果表明,α-淀粉酶的添加不仅可以缩短迟滞期,而且还能提高反应速率。 图6 CK组实测累积产甲烷及一级动力学, Gompertz模型拟合曲线 图7 10 mg·g-1试验组实测累积产甲烷及 一级动力学,Gompertz模型拟合曲线 图8 20 mg·g-1试验组实测累积产甲烷及 一级动力学,Gompertz模型拟合曲线 图9 30 mg·g-1试验组实测累积产甲烷及 一级动力学,Gompertz模型拟合曲线 图10 40 mg·g-1试验组实测累积产甲烷及 一级动力学,Gompertz模型拟合曲线 韩娅新、李超等对不同有机废弃物的产甲烷过程进行动力学分析[8, 12-13],不同发酵物料的性质及接种比对动力学拟合模型的选择有显著性影响,一级动力学模型适用于拟合接种比大于3:1的厌氧发酵产甲烷过程,其他条件下适宜采用Gompertz模型进行拟合,与本研究的结果一致。 表2 拟合方程对试验组累积甲烷产率曲线的拟合结果 表3为各试验组厌氧发酵过程中MPR及BDA。本研究中以MPR等于90%表示厌氧发酵反应过程结束,根据该指标可以确定各试验组的厌氧发酵水力停留时间(HRT)。CK组的HRT为17.9 d,30 mg·g-1试验组的HRT最短(15.3 d),比CK组缩短了14.5%。在工程实践中,HRT的适当缩短有利于降低厌氧发酵系统的建设和运行成本,提高容积甲烷产率。 分析发酵原料的组成成分,得到猪粪的化学组成为C270H485O141N18S2,根据方程式(1)计算得出其理论产甲烷潜力值为552.3 mL·g-1。BDA是衡量发酵物料转化产甲烷是否彻底的重要指标,其值越高,说明生物质转化的程度越高。适当添加α-淀粉酶可以提高BDA,在第15~35天,各试验组BDA值由大到小排序为30 mg·g-1﹥40 mg·g-1﹥20 mg·g-1﹥10 mg·g-1﹥CK组。30 mg·g-1试验组在第13天的BDA值已超过CK组第35天的值;反应结束时,与CK组相比提高了22.1%,说明α-淀粉酶的添加对BDA促进作用显著。 表3 厌氧发酵过程部分时间点的MPR和BDA (%) (1)α-淀粉酶的适量添加有利于促进有机物厌氧发酵底物的水解,提高甲烷产率。当α-淀粉酶添加量≤30 mg·g-1时,添加量与甲烷产率增加呈正相关。 (2)α-淀粉酶添加量为30 mg·g-1的试验组对甲烷产率的促进作用最为显著,其日产气峰值、甲烷产率、BDA分别为13 829 mL,384.9 mL·g-1VS,69.69%,与对照组比分别提高了18.1%,22.1%,22.1%。 (3)添加α-淀粉酶的猪粪厌氧发酵过程比较符合Gompertz模型,添加量为30 mg·g-1的试验组最大反应速率为32.63 mL·g-1d-1,比CK组提高24.6%,决定系数R2为0.9995。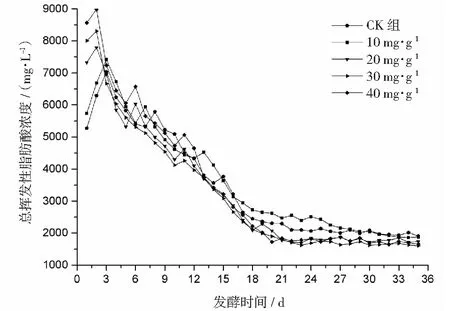
2.2 α-淀粉酶对沼气产气性能的影响
2.3 拟合模型的动力学分析





2.4 α-淀粉酶对BDA的影响
3 结论
