基于表型和ISSR标记滇产黄杨叶栒子遗传多样性分析
2020-04-21朱燕蕾郭凤根
朱燕蕾,郭凤根
(1. 云南农业大学 园林园艺学院,云南 昆明 650201;2. 云南农业大学 农学与生物技术学院,云南 昆明 650201)
黄杨叶栒子(Cotoneaster buxifoliusLindl.)是蔷薇科栒子属(Cotoneaster)的常绿至半常绿灌木,产四川、贵州、云南等省,生于海拔1 000~3 300 m的多石砾坡地、灌林丛中[1]。黄杨叶栒子在4—6月开花,花3~5朵,在8月至次年2月红色的果实挂满植株,是优良的观花、观果植物,根系发达,是优良的护坡植物,具有很高的开发价值[2]。野外调查发现,不同产地的黄杨叶栒子间在表型性状上存在较大差异,但根据前期查阅的文献,有关滇产黄杨叶栒子的遗传多样性研究尚未见报道[3-5]。
用表型性状来检测遗传变异是直接、简便、易行的方法,但表型性状受环境因素的影响,在某些情况下并不能反映遗传变异的真实状况。分子生物学技术的快速发展为遗传多样性检测提供了更直接和精确的方法,其中,ISSR(inter-simple sequence repeat)分子标记技术由于具有引物通用性强、利用率高、操作简便和重复性好等优点,在植物的遗传多样性、系统进化、DNA指纹图谱及核心种质构建等方面已得到广泛的应用[6-10]。本文以云南不同产地的黄杨叶栒子为材料,从表型性状和ISSR分子标记2个角度分析其遗传多样性,为其种质资源的合理利用和保护奠定基础。
1 材料与方法
1.1 材料
对云南省内7个州市15个县区市的51个野生黄杨叶栒子居群进行了调查和表型性状的观测,采集各居群叶片并用硅胶干燥于自封口袋中,编号后保存于-80 ℃冰箱,用于ISSR分子标记试验。各居群的产地、代码、纬度、经度和海拔等信息见表1。
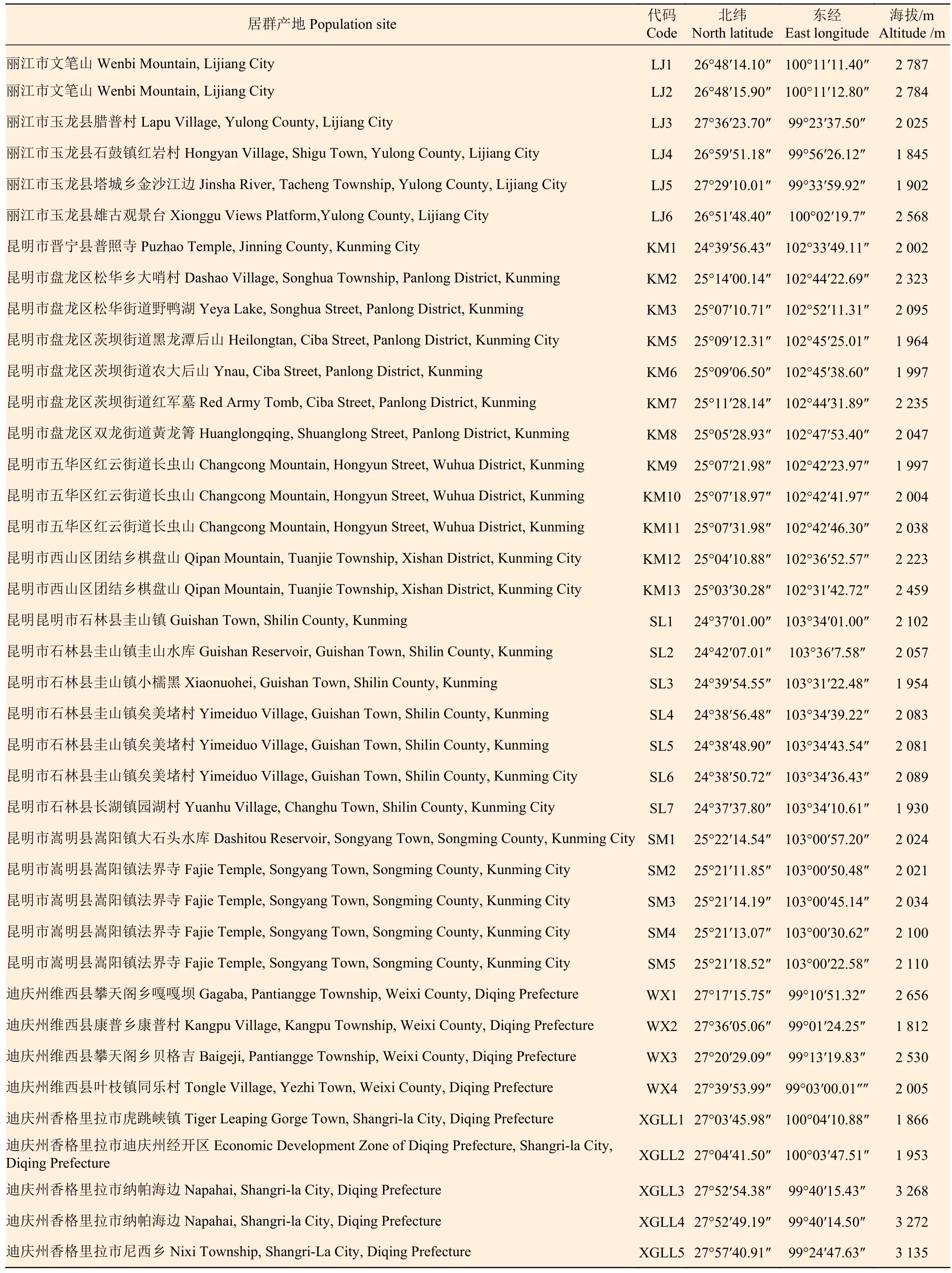
表 1 51份黄杨叶栒子材料的来源及重要地理特征Table 1 Sources and important geographic characters of 51 C. buxifolius samples
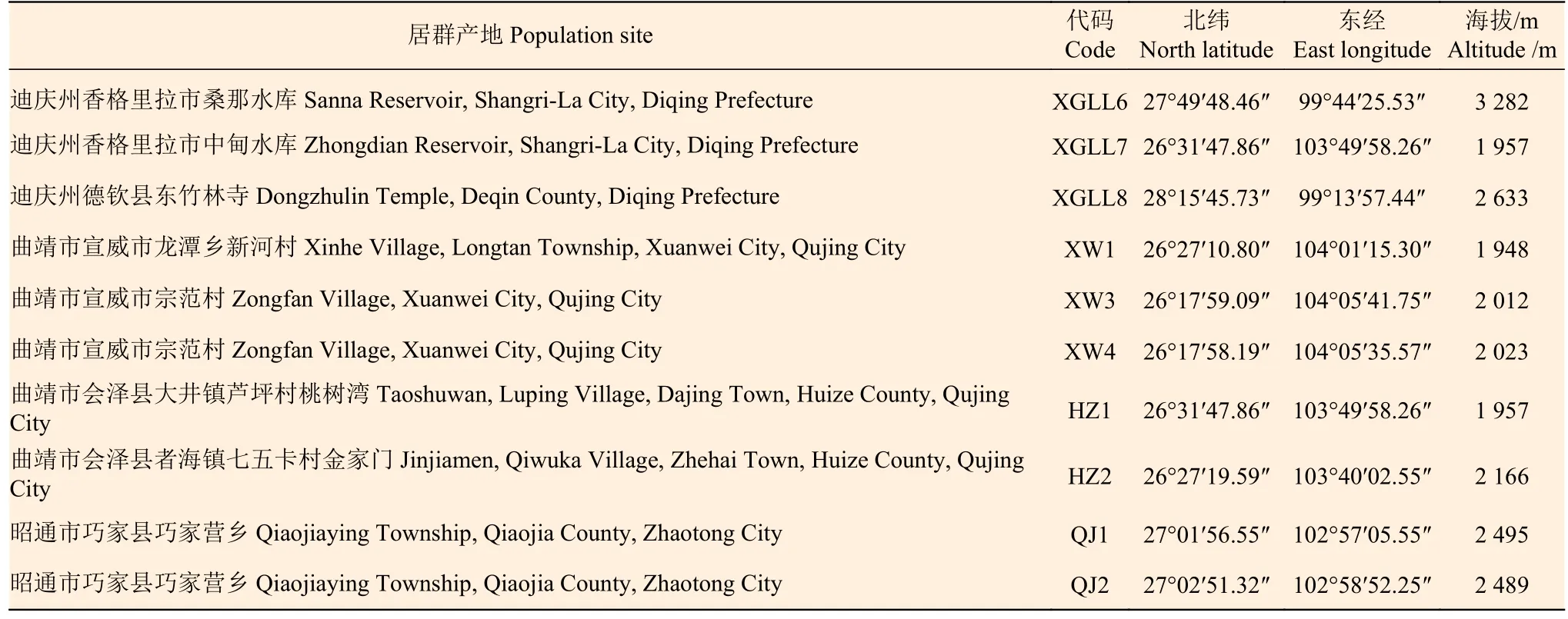
续表 1
1.2 表型性状的观测
观测了51个野生黄杨叶栒子居群的枝条颜色和夹角及毛被情况、叶片的形态及毛被情况、花朵的数量、花瓣的形状和颜色,将采集的果实带回实验室后观测果实的颜色、形状及大小、萼筒毛被情况、萼裂片形状及毛被情况、每果小核数,共计观测了33个表型性状。
1.3 ISSR分析
采用改良CTAB法提取黄杨叶栒子各样品的叶片总DNA,用1%琼脂糖凝胶电泳检测DNA质量,用紫外分光光度法测定DNA浓度,达到ISSR分子标记要求的DNA保存于-20 ℃冰箱备用[11-13]。对DNA质量检测,有27个居群达到后续分析的要求。从100条ISSR引物中筛选出24条适于黄杨叶栒子扩增的引物,用优化后的反应体系(25 μL反应体系中含 2.5 mmol·L-1Mg2+、2U Taq DNA聚合酶、0.15 mmol·L-1dNTPs、0.48 mol·L-1引物和100 ng模板DNA)对黄杨叶栒子的DNA模板进行扩增,程序如下:95 ℃预变性5 min,1个循环;94 ℃变性1 min,退火1 min,72 ℃延伸1.5 min,进行38个循环;最后72 ℃延伸10 min。用1%的琼脂糖凝胶电泳检测PCR产物并拍摄照片,根据有带为1、无带为0的原则对重复性好且清晰易辨的条带进行统计,获得27个居群的24条引物的“0,1”ISSR 数据矩阵。
1.4 数据的统计与分析
对表型性状中的质量性状进行赋值后参与聚类运算;对数量性状则直接用SPSS 16.0软件进行平均值、标准差、标准误的计算和方差分析,进而计算出各性状的变异系数和相对极差。依据33个表型性状的欧氏平方距离系数,用UPGMA法构建基于表型性状的树系图。
依据获得的27个居群的24条引物的ISSR数据计算各引物的总条带数、多态性条带数和多态性百分率;应用SPSS 16.0软件,根据欧氏平方距离系数,用UPGMA法构建基于ISSR标记的27个黄杨叶栒子居群的树系图。采用POPGEN 32软件计算滇西北群体(DXB)、宣威群体(XW)、嵩明群体(SM)、石林群体(SL)、昆明群体(KM)和会泽群体(HZ)等6个产区的黄杨叶栒子居群的多态性位点百分比(PPB)、观测等位基因数(Na)、有效等位基因数(Ne)、Nei’s基因多样性指数(H)、Shannon信息指数(I)和遗传多样性指数(Ht)。
2 结果与分析
2.1 基于表型性状的遗传多样性分析
2.1.1 表型性状分析 各黄杨叶栒子居群的枝条颜色全部为棕褐色;所有小枝都被糙伏毛;二级枝条与主枝之间的夹角为80.07°~37.67°,平均为42.4°,平均变异系数为16.53%;叶片长度为0.6~1.83 cm,叶片宽度为0.37~1.26 cm,二者变异系数分别为23.19%和28.78%;叶片质地纸质占100%;叶片形状为卵形;叶基形状98.04%为近圆形,还有1.96%为宽楔形;叶尖形状有98.04%为锐尖,有1.96%为钝尖;叶片正面98.04%有毛,有1.96%为幼时有毛;叶片正面都疏被白色绒毛;叶片背面都疏被灰白色绒毛;100%花序具有1朵以上的花;花朵颜色全部为白色;果实均为红色钟状果;萼筒全部密疏绒毛;果实纵径为5.48~8.29 mm,果实的横径为5.37~7.76 mm,变异系数分别为18.00%和17.34%;果实萼裂片形状变异相对较大,变异系数为69.02%,其中,尖三角形占54.90%,宽三角形间隔缝大占33.33%,宽三角形占9.8%,几乎不开裂占1.96%;果实萼裂片中94.12%被毛,其中,66.67%是被密绒毛,而33.33%被疏绒毛,其变异系数达35.17%;26.56%的果实包含1个小核,39.06%的果实包含2个小核,28.91%的果实包含3小核,4.69%的果实包含4小核,只有0.78%的果实含5小核。
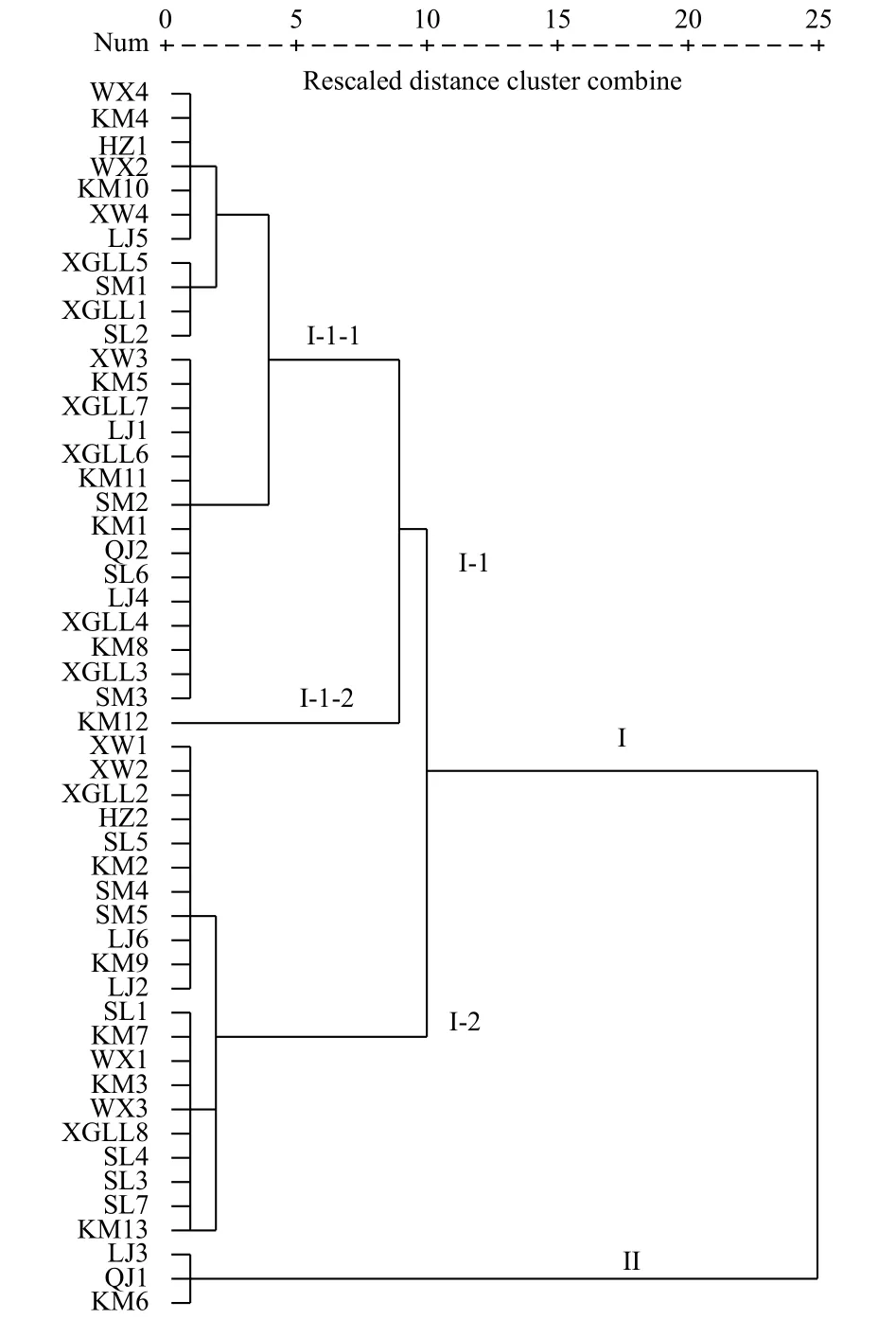
图 1 基于表型性状51份黄杨叶栒子资源UPGMA聚类图Fig. 1 UPGMA dendrogram of 51 C. buxifolius resources based on the phenotypic traits
2.1.2 各表型性状的聚类分析 依据33个表型性状的51个野生黄杨叶栒子居群的UPGMA树系图见图1。各居群之间的距离在25阈值内,51个样本在遗传距离系数25处可以划分为2个类群,其中,类群II包括了3个居群,二级枝条与主枝之间的夹角为 78°~80°,叶片长度为 0.9~1.0 cm,叶片宽度为0.5~ 0.55 cm,属于所有居群中夹角最大的一组;类群I中,在遗传距离系数为10处可以分为I-1和I-2两个分支,其中,I-2分支中包括了21个居群,二级枝条与主枝之间的夹角多数为62°~68°,居群分布海拔在 1 900~2 500 m;I-1 类群中,在遗传距离系数8处分为I-1-1和I-1-2两个分支,分别包括了26个和1个居群。I-1-1分支中,一个小分支夹角基本为53°~57°,另一小分支则保持在42°~49°;I-1-2分支中仅有1个来自昆明居群的栒子,其夹角最小(37°)。从图1可看出:51个黄杨叶栒子居群间的亲缘关系,多数地理位置或者海拔相近的居群能够聚在同一分支,但有些同一产地的不同居群却分布在不同的分支中。
2.2 基于ISSR分子标记的遗传多样性分析
2.2.1 黄杨叶栒子 ISSR多样性分析 24条ISSR引物对27个黄杨叶栒子居群共扩增出228条带,其中,多态性条带有153条,每条引物扩增得到的总条带数为5~14,平均9.50条;多态性条带数为3~13,平均6.38条;多态百分率为58.33%~100.00%,多态百分率达81.56%(表2)。
将采自滇西北、宣威、嵩明、石林、昆明、会泽地区的27个居群按产区进行归类后用PopGene 32 软件分析,得表3。6个产区平均扩增出151.67多态性位点,多态性位点比率达到66.52%,其中,嵩明产区多态性位点最多,达170,多态性位点比率达74.56%;会泽产区的多态性位点最少,只有122,比率53.51%。群体间平均Nei`s遗传多样性指数为0.413 2,Shannon多样性指数为0.595 9,群体内多态性位点比率为53.51~74.56%。Nei`s遗传多样性指数(H)为0.156 2~0.432 5,平均为0.352 0;Shannon多样性指数(I)为 0.216 6~0.621 2,平均 0.505 9。6个产地中,HZ群体(H=0.156 2,I=0.216 6)的遗传多样性水平最低,KM群体(H=0.432 5,I=0.621 2)的遗传多样性水平最高。Nei`s遗传多样性指数、Shannon多样性指数和多态性位点比率3个指标所表现的多样性趋势基本一致,6个群体遗传多样性由高到低依次为:KM 群体 >XW 群体 > SL 群体 >SM 群体>DXB群体>HZ群体。经计算可知,遗传多样性指数(Ht)为0.413 2±0.011 0。XW群体与KM群体的遗传距离最大(0.980 8),XW群体与DXB群体遗传距离相对较近(0.330 2);XW群体与KM群体遗传一致度最低(0.375 0),XW群体与DXB群体遗传一致度最高(0.718 8)。其余遗传一致度基本在0.48左右,4个群体间的遗传一致度变幅为 0.375 0~0.718 8。
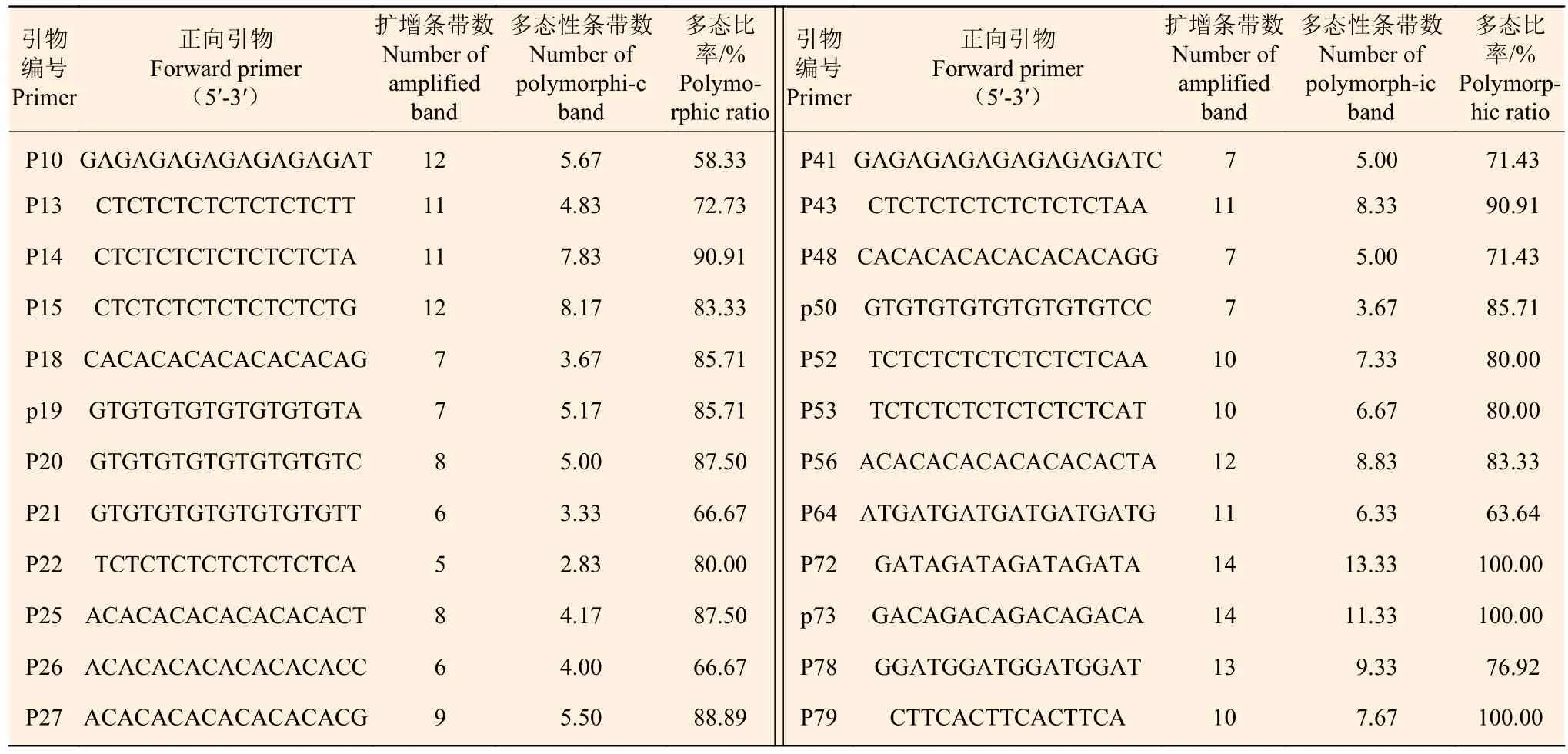
表 2 24条引物的扩增结果Table 2 Amplification results of 24 primers
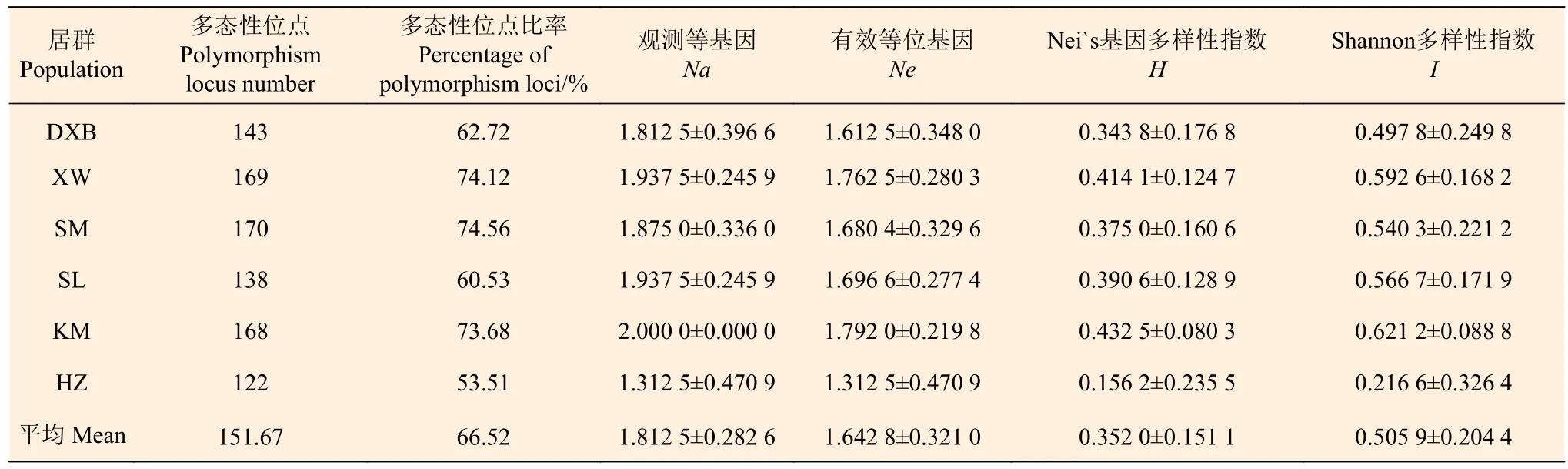
表 3 栒子群体的遗传多样性Table 3 Genetic diversity of populations of C. buxifolius
2.2.2 基于ISSR分子标记的黄杨叶栒子居群间亲缘关系 用SPSS 16.0软件计算27个黄杨叶栒子居群间的欧氏平方距离系数,采用 UPGMA 聚类方法,得到27个黄杨叶栒子居群的聚类树系图(图2)。从图2可以看出:以距离系数23为界,可以将27个居群划分为3大分支:分支I包含13个居群,又可进一步细分为昆明、嵩明组(I-1),5个居群基本分布在海拔2 050 m左右,石林等组(I-2)6个居群分布在海拔1 950~2 100 m;分支II包含3个居群,包括了全部来自滇西北的居群;分支III包含11个居群,可进一步细分为昆明、宣威等组(III-1),包括了9个居群,昆明与会泽(III-2、III-3)2个分支,各自包括1个居群,其均分布在海拔2 400 m的地区。
3 讨论
3.1 黄杨叶栒子的遗传多样性丰富
表型性状是种质资源遗传多样性的一种表现方式,与分子标记有着同等重要的作用,同时也是育种和种质保护研究的基础工作[14-15]。本研究选取33个表型性状,对51个野生黄杨叶栒子居群的遗传多样性进行了分析,发现有22个性状均表现出不同程度的变异,且变异主要存在于居群之间,变异系数较大的性状有萼裂片形状(69.02%)、萼裂片毛被情况(35.71%)、叶片宽度(28.78%)、叶片长度(23.19%)等,说明黄杨叶栒子居群间在果实萼裂片和叶片形态上存在较大的差异,大于野生玫瑰(18.48%)[16]、假俭草(25.59%)[17]、垂珠花(22.64%)[18],但这些差异均没有准噶尔山楂(86.10%)[19]的大。

图 2 基于ISSR标记分析27份黄杨叶栒子资源UPGMA聚类图Fig. 2 UPGMA dendrogram of 27 C. buxifolius resources based on the ISSR marker
ISSR分子标记可以产生丰富的多态位点,从而揭示种间、种内遗传变异情况[4]。本文利用ISSR分子标记对27个野生黄杨叶栒子居群进行遗传多样性分析,发现多态百分率达81.56%,说明云南的黄杨叶栒子资源存在较丰富的遗传多样性,其丰富程度与枣庄石榴(82.86%)[7]相近,但不如大别山区的白檀(92.62%)[20]和杜鹃花(98.94%)[21]高。
3.2 黄杨叶栒子居群间的亲缘关系
本文用33个表型性状构建了51个黄杨叶栒子居群的树系图(图1),用24个ISSR引物的228条条带构建了27个黄杨叶栒子居群的树系图(图2)。2张图对比分析可知:在ISSR聚类图中I-1分支中的5个居群,有 3个居群(SM1、SM2、KM8)在表型性状聚类图中都属于I-1-1这个分支,表型性状和ISSR分子标记所得结果都表明这3个居群的亲缘关系近;而其中的KM3居群在表型性状图中属于I-2分支,2种聚类方法结果不一致。分析此现象出现的原因:一是黄杨叶栒子居群间的亲缘关系与地理位置密切相关,分布在同一地区的各个居群多数能聚在一起,如昆明群体、滇西北群体、石林群体和会泽群体的部分材料可分别聚为一类,说明分布位置的隔离促进了黄杨叶栒子居群间的遗传分化[22-23];二是个别居群未能与同一产区的其他居群聚在一起,而聚进了其他产区的居群之中,其原因可能是同一产区内各居群的小生境不完全相同,在不同产区内也有相似的小生境,生境的不同导致不同居群间发生遗传分化和趋同进化。在材料采集时发现,虽然有些居群采集自同一产地,但它们的生境(土壤、水分、坡度、坡向、光照、海拔高度、伴生植物等)各不相同;但有些居群虽然采自不同的产地,但却拥有极为相似的生境,导致黄杨叶栒子呈现地域性趋同进化[24]。
3.3 地理隔离促进栒子居群间的遗传分化
正如沙棘,由于其分布区广泛,分布区内气候、土壤和海拔等生态因子的差异,地理隔离促进其高度种群遗传分化,形成11亚种[1,25]。地理隔离是物种间基因交流的天然屏障,也是影响居群间遗传分化的重要因素[26]。在聚类分析中,昆明群体(I-1分支中 3个居群和III-1亚分支中3个居群)、石林群体(I-2亚分支中6个居群)、滇西北群体(II分支中3个居群)可分别聚为一类。昆明群体和石林群体的栒子分布于滇中盆地区域,但2个地区相距70 km,受虫媒和鸟媒飞行距离的影响,依靠其在居群间传播花粉的能力必定受到地理距离的限制[27],空间隔离是居群间遗传分化的因素;而滇西北群体则分布在三江并流区域,群体间在地理分布上有横断山脉阻隔,使得物种区域分布的山脉阻隔特点突出,对黄杨叶栒子不同的居群具有天然的隔离作用,山脉阻隔严重影响了居群间花粉及种子散布,也使居群之间的基因交流受到了一定程度的阻碍[28],是引起居群间遗传分化的重要因素。
表型性状是植物的外部特征,标记数量少,且受到生境、生长状况和时期等方面因素的影响,利用受限。表型性状与ISSR分子标记所提供的DNA片段信息相结合,可以揭示黄杨叶栒子遗传的本质特征。同时通过遗传聚类了解栒子遗传变异在地理上的分布格局,对于制定科学的保护策略起着极为重要的作用。上述黄杨叶栒子遗传多样性分析将为今后黄杨叶栒子种质资源分类、起源、保存和利用奠定基础。
4 结论
本研究表明:滇产黄杨叶栒子表型性状和分子遗传多样性丰富。根据分子标记聚类结果可以将27个居群分为3个分支。黄杨叶栒子居群间的亲缘关系及分布与地理位置、海拔位置有密切关系。
