温度与溶解态有机磷源对中肋骨条藻生长的影响
2020-02-07张小华刘东艳
张小华, 刘东艳
温度与溶解态有机磷源对中肋骨条藻生长的影响
张小华1, 2, 3, 刘东艳4
(1. 中国科学院 烟台海岸带研究所 海岸环境过程重点实验室, 山东 烟台 264003; 2. 中国科学院大学, 北京 100049; 3. 滨州医学院 药学院, 山东 烟台 264003; 4. 华东师范大学 河口与海岸学国家重点实验室, 上海 200241)
为探讨温度与溶解态有机磷(Dissolved organic phosphorus, DOP)对浮游植物生长的共同影响, 本文以中肋骨条藻为研究对象, 选取含有C-O-P键的三磷酸腺苷(Adenosine triphosphate, ATP)、甘油磷酸钠(Sodium glycerophosphate, SG-P)、6-磷酸葡萄糖(Glucose 6-phosphate, G-6-P)和含有C-P键的草甘膦(Glyphosate, G-P)为不同类型的DOP, 检测了4种温度条件下(20、23、26、29℃)藻细胞的生长状况以及培养液中总溶解态磷(Total dissolved phosphorus, TDP)、溶解态无机磷(Dissolved inorganic phosphorus, DIP)浓度与藻细胞内碱性磷酸酶(Alkaline phosphatase, AP)的变化。研究结果表明: 含有C-O-P键的ATP、SG-P和G-6-P均能较好地维持中肋骨条藻的生长, 并且升温可进一步促进各组藻细胞的生长, 最大藻细胞密度分别达到176.7×104、218.8×104和 178×104cell/mL; 此外升温还可加速ATP、SG-P和G-6-P组培养液中TDP浓度的下降, 以及ATP组培养液中DIP浓度的升高; 但各组藻细胞内AP活性在不同温度条件下均处于较低水平(< 10 fmol pNP /(cell· h))。与含有C-O-P键的DOP相比, G-P组的藻细胞在各温度条件下生长密度均较低, 最大藻细胞密度仅为40.7×104cell/mL ; 但其藻细胞内AP活性显著高于ATP、SG-P和G-6-P组[最高达83 fmol pNP /(cell·h)]且随温度升高而降低。以上结果表明, 不同类型DOP会显著影响中肋骨条藻的生长, 其中ATP、SG-P和G-6-P能更为有效地被中肋骨条藻生长所利用, 并且温度升高(20~29℃)是促进3种DOP条件下中肋骨条藻生长的重要环境因素, 但该效应可能并不依赖于AP活性。
中肋骨条藻; 温度; 生长; 溶解态有机磷; 溶解态无机磷; 碱性磷酸酶
溶解态无机磷(Dissolved inorganic phosphorus, DIP)是海洋浮游植物生长的必需营养元素, 然而, 很多海域存在季节性磷限制的现象[1-2]。已有研究表明, 在DIP限制的条件下, 部分浮游植物可以利用溶解态有机磷(Dissolved organic phosphorus, DOP), 大量繁殖生长, 甚至形成赤潮[3]。DOP是近岸海域中广泛存在的一种重要磷源, 在海洋总溶解态磷(Total dissolved phosphorus, TDP)中占有很大比例[4-6]。例如, 有调查表明, 黄河口及邻近海域DOP的含量大约贡献了TDP的85%左右[7], 是导致水体富营养化的重要因素之一[8]。DOP结构复杂, 由众多复杂混合物组成。根据其所含C、P键型不同, 基本可分为两大类有机化合物: 含有C-O-P键的磷酸酯和含有C-P键的膦酸酯[9]。大量研究表明DOP需要先被转化为DIP才能被浮游植物吸收利用[10-11], 其转换利用能力一方面受到DOP的结构影响, 另一方面受到海洋环境因子的影响。例如, 海洋中痕量金属元素(铝、锌等)的存在可以促进浮游植物对DOP的利用[12-13]; 较高的海水温度也会促进浮游植物对DOP的转化吸收速率[14]。
有研究表明, 海洋变暖会加剧海水的富营养化效应,促进浮游植物对营养物质的吸收利用, 加剧赤潮的爆发频率和规模[15-16]。DOP是富营养化海域的重要磷源, 浮游植物在海水暖化条件下, 对不同结构DOP的吸收利用能力可显著影响到生物生产量, 乃至赤潮的发生, 因此, 有必要深入探讨温度与DOP对浮游植物生长的影响效应。本研究选取我国东部沿海的一种常见赤潮藻-中肋骨条藻()[17-18]为研究对象, 在四个不同的温度条件下(20、23、26、29℃), 分别添加三磷酸腺苷(Adenosine triphosphate, ATP)、6-磷酸葡萄糖(Glucose 6-phosphate, G-6-P)、甘油磷酸钠(Sodium glycerophosphate, SG-P)和草甘膦(Glyphosate, G-P)四种不同形态的DOP, 通过分析藻细胞生长特征, 以及培养液中TDP、DIP浓度和碱性磷酸酶(alkaline phosphatase, AP)活性变化, 探讨了升温与不同DOP对中肋骨条藻生长的影响特征, 研究结果将为揭示海水暖化条件下DOP丰富海区浮游植物的生长状态提供一定的科学依据。
1 材料与方法
1.1 藻种培养及实验设计
中肋骨条藻()分离自我国黄海海域。实验所用玻璃器皿均使用10% HCl浸泡24 h, 纯净水冲洗干净后, 放于烘箱烘干备用。培养所用海水为人工海水, 参照Harrison等[19]的方法配制, pH值为8.0±0.1, 盐度为30±1, 经121℃高压湿热灭菌30 min,室温冷却后备用。实验室中肋骨条藻的培养条件光照为55 μmol/(m2· s), 光暗周期为12 h/12 h, 温度为20℃, 培养液为f/2营养盐配方[20]。
实验选择4种不同类型的DOP, 分别代表海洋中存在的两种P键类别: G-6-P、ATP和SG-P代表含有C-O-P键的磷酸酯, G-P代表含有C-P键的膦酸酯。参照f/2培养基中NaH2PO4的浓度(36 μmol/L), 分别添加36 μmol/L G-6-P、12 μmol/L ATP(约为36 μmol/L P)、36 μmol/LSG-P和36 μmol/L G-P配制不同类别磷源的培养液, 其他各种元素依旧按f/2配方添加; 对照组使用f/2培养基。中肋骨条藻生长最适温度在20~30℃之间[21-22], 因此本次实验在此区间设定4个温度条件, 分别为20、23、26、29℃。接种指数生长期的中肋骨条藻到各含有600 mL不同磷源培养液的1 L锥形瓶中(表1), 起始藻细胞密度为5×104cell/mL, 然后分别置于预设4个温度条件的恒温光照培养箱中, 连续培养7 d, 每个处理组合均设3个重复。每天固定时间从各组中取50 mL样品, 用于藻细胞计数、培养液中TDP、DIP浓度以及细胞AP活性的测定。
1.2 藻细胞计数
取100 μL样品用于藻细胞计数, 取样前混匀藻液, 将卢戈氏液加入取样样品中固定藻细胞, 使用0.1 mL浮游植物计数框(20 mm×20 mm)于显微镜下对每个样品进行计数, 计算每mL样品中含有的藻细胞个数。

表1 实验分组情况列表
1.3 生长速率计算
根据每天获得的藻细胞密度, 采用下列公式[23]计算本实验中肋骨条藻的生长速率:
= (ln2– ln1)/(2–1),
式中,2和1分别代表时间2和1的细胞密度。
1.4 培养液中DIP与TDP浓度的测定
50 mL的培养液离心后, 上清经醋酸纤维膜过滤, 用于测定培养液中DIP和TDP的浓度。DIP采用以抗坏血酸为还原剂的磷钼蓝比色法测定(海洋调查规范第4部分: 海水化学要素调查GB17378.4- 2007)。TDP根据Jeffries等[24]的方法进行测定, 过滤培养液加入50 g/L的过硫酸钾, 121℃消解30 min, 冷却至室温, 再采用以抗坏血酸为还原剂的磷钼蓝比色法进行测定。
1.5 胞内AP活性的测定
离心收集的藻细胞用于胞内AP活性的测定。参照Lin等[25]的方法, 以对硝基苯磷酸二钠(4-Nitrophenyl phosphate, pNPP)为底物测定AP活性变化。收集到的藻细胞加入1 mL 0.05 mol/L的Tris-HCl (pH=9)的缓冲液, 在冰浴条件下匀浆1~2 min, 匀浆后离心取上清, 与50 μL 20 mmol/L的pNPP混匀, 对应培养温度条件下黑暗中放置2 h, 随后将样品放置冰上终止酶反应, 10 000离心2 min, 取上清于405 nm处测定胞内AP活性。酶活计算单位为单个藻细胞含有的AP 1 h降解pNPP所产生对硝基苯酚(p-nitrophenol, pNP)的fmol数。
1.6 数据处理与分析
所有实验测定结果均表示为平均值±标准方差(=3)。采用三因子方差分析DOP(ATP、G-6-P、SG-P和G-P)、温度(20、23、26、29℃)和取样时间对藻细胞密度、TDP、DIP浓度和AP活性的影响, 如果交互作用因子<0.05, 则使用单因素重复方差分析来确定差异性。重复测量方差分析时, 如果重复性方差分析的方差-协方差矩阵的球形假设被违反, 则对自由度(df)进行Greenhouse-Geisser校正。所有统计分析均采用SPSS 17.0软件进行。
2 结果与分析
2.1 中肋骨条藻生长特征比较
中肋骨条藻在不同温度与DOP组中表现出不同的生长特征(图1; 表2)。在ATP组中, 藻细胞在4 d后快速生长, 最大藻细胞密度与平均生长率出现在29℃条件下 (图1a; 表2); SG-P和G-6-P组的藻细胞生长变化趋势相似, 5 d后快速生长, 最高藻细胞密度与平均生长率均出现在26℃条件下(图1b, c; 表2)。相比之下, G-P组的藻细胞密度维持在较低水平, 没有出现显著生长, 生长速率明显低于其它组, 29℃条件下的生长略高于其他温度条件(图1d; 表2)。NaH2PO4组随着温度的升高细胞密度显著上升, 29℃生长最好(图1e)。以上结果可以看出, 含有C-O-P键的磷酸酯(ATP, SG-P和G-6-P )表现出与NaH2PO4组相似的生长趋势, 较高的温度条件可以显著促进中肋骨条藻的生长; 而含有C-P键的膦酸酯(G-P)对中肋骨条藻生长的影响不显著。

图1 不同温度和DOP条件下中肋骨条藻的生长特征

表2 中肋骨条藻在各组中的最高生物量(×104 cell/ mL)及平均生长速率(d–1)
三因子方差分析表明, 温度与DOP类型均对中肋骨条藻生长有显著影响, 并且两者之间存在显著的交互效应(表3)。单因素重复性方差分析发现, 在相同的DOP类型下, 温度对藻细胞生长均有显著影响(<0.01); 在相同的温度下, SG-P和G-6-P组之间的生长差异不显著外(>0.05), 其余各DOP组之间均具有显著差异(<0.01)。
2.2 培养液中磷酸盐的变化特征
培养液中TDP与DIP的浓度变化在不同DOP组中显著不同(图2)。ATP组的培养液中, TDP在实验过程中迅速下降, 29℃下降最为显著(图2a); 培养液中DIP浓度在实验前5 d显著上升, 随后下降至实验结束(图2b)。SG-P和G-6-P组培养液中的TDP均在29℃条件下下降最为显著(图2c, e); 两组培养液中DIP浓度变化相似, 均在实验后期有少量DIP产生(<10 μmol/mL)(图2d, f)。相比之下, G-P组培养液中TDP浓度到实验结束时均呈现上升趋势(图2g); 整个实验过程均难以检测到DIP(图2h)。三因子方差分析发现, 温度和DOP类型对TDP与DIP浓度均有显著影响(表3), 且DOP类型对TDP和DIP浓度的影响较温度更为显著。在相同温度条件下, G-P组的TDP浓度与其他各DOP组之间差异显著(<0.01); ATP组的DIP浓度与其他各DOP组之间差异显著(<0.01)。
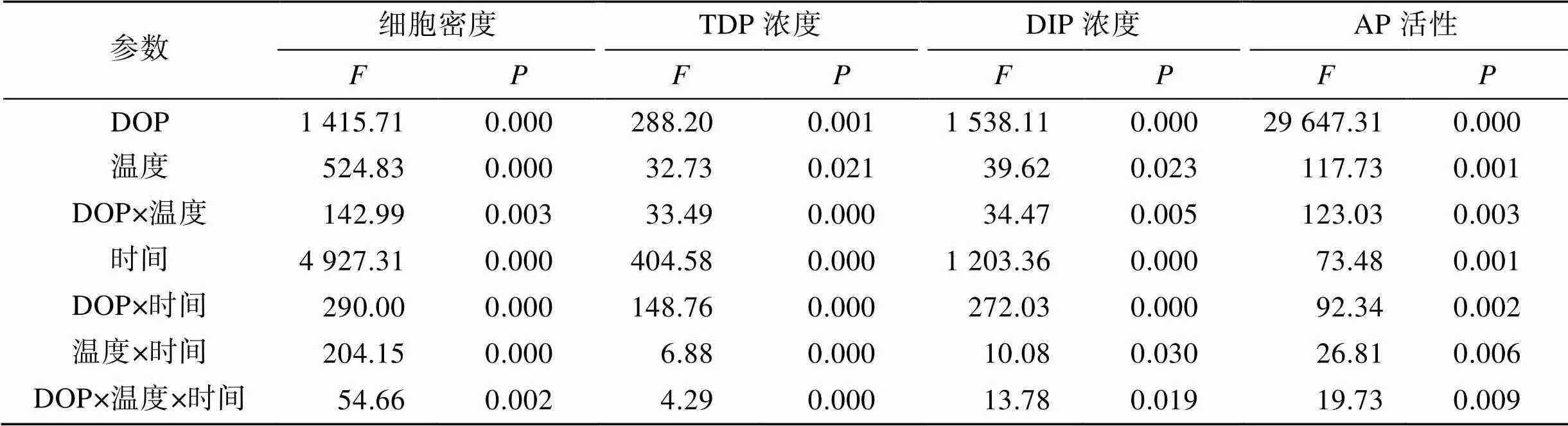
表3 三因子方差分析DOP类型、温度和取样时间对细胞密度、TDP和DIP浓度以及AP活性的影响
注:表示这些因素或其相互作用的显著性,<0.05时表示差异显著.
2.3 细胞碱性磷酸酶活性变化特征
不同温度条件与DOP组中, 中肋骨条藻胞内AP活性表现出显著不同的变化特征(图3)。AP 活性在ATP组中不活跃(<10 fmol pNP/(cell·h)), 仅在26℃条件下培养的第6, 7 d出现峰值(图3a); SG-P和G-6-P组中, AP活性的变化特征相似(图3b, c), 培养期间变化不显著。各温度条件下G-P组的AP活性均显著升高, 其中酶活最高值出现在20℃第5 d(83 fmol pNP/ (cell·h))(图3d)。三因子方差分析结果表明, 温度和DOP类型均对AP活性有显著影响(表3)。相同温度条件下, G-P组AP活性与其他各DOP组之间均存在显著差异(<0.01); 此外G-P组AP活性在26℃与29℃条件下差异不显著(>0.05), 其余各温度条件下的AP活性均有显著差异(<0.01)。
3 讨论
DOP是富营养海岸水域存在的重要磷源之一, 其中, 磷酸酯类DOP可以占到总DOP类型的75%左右[26-28]。目前已有多种磷酯化合物被鉴定出来, 其中,核酸、游离核苷酸(如ATP)、甘油磷酸脂(如SG-P)、糖类衍生物(如SG-P)等是其主要组成成分[29-30]。ATP、G-6-P、SG-P都属于小分子磷酸酯, 含有C-O-P键, 易于被降解, 因此常被用来作为研究浮游植物对海水DOP利用的代表性对象。已有研究表明东海原甲藻()[31-32]、球形棕囊藻()[33]、具槽帕拉藻()[34]、塔玛亚历山大藻()[35]等可利用ATP、G-6-P、SG-P等DOP化合物维持生长。膦酸酯约占海洋中DOP的25%[30], 广泛分布于许多海洋生命形式中。近年来由于G-P的广泛应用导致其在沿海海域中积累, 成为水体膦酸盐的重要组成部分[36]。G-P属于小分子膦酸盐, 含有化学稳定的C-P键, 从能量上讲, 它比水解C-O-P键更困难, 因此, 只有少数细菌和海洋浮游植物被报道可以利用G-P作为磷营养源, 例如小定鞭藻()[37]、铜绿微囊藻()[38]以及丝状蓝细菌鱼腥藻(sp.)[39]等。不同浮游植物对不同DOP的利用能力不同[35], 且易受环境因子影响。

图2 不同温度和DOP条件下培养液中TDP和DIP浓度变化特征

图3 不同温度和DOP条件下中肋骨条藻AP酶活性变化特征
本研究发现中肋骨条藻可以在ATP、G-6-P和SG-P条件下快速生长, 而在G-P条件下, 中肋骨条藻的生长显著低于其他DOP组。这表明中肋骨条藻对不同结构类型DOP的利用能力不同, 更易利用磷酸酯。在相同DOP处理条件下, 中肋骨条藻的生长响应了较高的温度条件, 这与Tian等[40]报道中肋骨条藻在较高温度下分裂更快, 更易引起赤潮的结果一致。通过监测藻细胞生长过程中培养液里TDP、DIP的变化, 进一步了解中肋骨条藻对不同DOP的利用过程。实验发现, 在ATP、G-6-P和SG-P培养液中TDP浓度迅速下降, 表明中肋骨条藻可以有效利用磷酸酯ATP、G-6-P和SG-P维持自身生长, 在较高温度培养液中的TDP消耗更为显著。G-P组培养液中TDP浓度没有下降反而略有升高, 表明中肋骨条藻不能直接利用G-P, 培养液中TDP浓度升高可能是由于部分细胞死亡后, 细胞破裂导致含磷内容物释放到培养液中。有研究报道表明, DOP的吸收利用是通过被酶水解成DIP的途径来进行的[11, 41], 但也有研究发现一些小分子DOP可以被藻类直接吸收利用, 例如, 米氏凯伦藻()可以直接吸收利用G-6-P[42]。本研究发现ATP组培养液中有大量DIP, 而SG-P和G-6-P组中仅有少量DIP, 表明中肋骨条藻对ATP与SG-P和G-6-P的利用途径可能不同。温度对ATP组DIP浓度变化有显著影响(<0.01), 培养液中DIP浓度随温度的升高显著增多, 表明温度升高可能加速ATP水解产生更多的DIP, 从而促进了中肋骨条藻的生长。
AP被认为是最重要的DOP利用酶, 在DIP缺失的环境被诱导合成, 通过分解水体中的DOP 为浮游植物提供磷源[10, 25]。本研究发现, 不同类型DOP条件下的藻细胞内AP活性存在较大差异, 其中ATP、SG-P和G-6-P组的AP活性均较低且各组间无显著差异(>0.05), 而G-P组的AP活性显著高于ATP、SG-P和G-6-P组的AP活性(<0.01), 表明在含有C-P键的DOP条件下中肋骨条藻的AP活性较高。有研究报道, 当环境中可利用磷匮乏时, 浮游植物会通过合成AP利用细胞内储存的多聚磷酸盐维持细胞较低水平的生长[31]。因为G-P较难被水解, 中肋骨条藻可能会通过合成较高AP利用胞内多聚磷酸盐, 这可能是本实验在G-P营养条件下, 中肋骨条藻细胞生长率较低及培养液中TDP没有降低现象的一种解释。ATP、SG-P和G-6-P组的AP活性均较低, 可能是小分子磷酸酯较易被水解, 藻细胞保持较低的AP水平就能维持较好生长, 也可能是藻细胞存在其他酶利用DOP维持生长[41, 43]。Healey等[44]研究表明温度会不同程度上影响浮游植物AP活性。本研究中, 温度变化对G-P条件下的细胞AP活性有显著影响, AP活性在20℃和23℃下活性更高。
4 结论
1) 不同类型DOP会显著影响中肋骨条藻的生长。与G-P相比, ATP、SG-P和G-6-P能更为有效地被中肋骨条藻生长所利用。
2) 本实验温度范围内(20~29℃), 温度升高能显著促进中肋骨条藻在ATP、SG-P和G-6-P三种DOP条件下的生长, 但该效应可能并不依赖于AP活性。
[1] Xu S, Song J, Li X, et al. Changes in nitrogen and phosphorus and their effects on phytoplankton in the Bohai Sea[J]. Chinese Journal of Oceanology and Limnology, 2010, 28(4): 945-952.
[2] Turner R E, Rabalais N N. Nitrogen and phosphorus phytoplankton growth limitation in the northern Gulf of Mexico[J]. Aquatic Microbial Ecology, 2013, 68(2): 159-169.
[3] Ou L, Qin X, Shi X, et al. Alkaline phosphatase activities and regulation in three harmfulspecies from the coastal waters of the East China Sea[J]. Microbial Ecology, 2020, 79(2): 459-471.
[4] Benitez-Nelson C R. The biogeochemical cycling of phosphorus in marine systems[J]. Earth-Science Reviews, 2000, 51(1-4): 109-135.
[5] Sebastián M, Arístegui J, Montero M F, et al. Kinetics of alkaline phosphatase activity, and effect of phosphate enrichment: a case study in the NW African upwelling region[J]. Marine Ecology Progress Series, 2004, 270: 1-13.
[6] Karl D M, Yanagi K. Partial characterization of the dissolved organic phosphorus pool in the oligotrophic North Pacific Ocean[J]. Limnology and Oceanography, 1997, 42(6): 1398-1405.
[7] 张欣泉, 邓春梅, 魏伟, 等. 黄河口及邻近海域溶解态无机磷、有机磷、总磷的分布研究[J]. 环境科学学报, 2007, 27(4): 660-666. Zhang Xinquan, Deng Chunmei, Wei Wei, et al. The distribution of dissolved inorganic phosphorus, dissolved organic phosphorus, dissolved total phosphorus in the Yellow River estuary and adjacent water[J]. Acta Scientiae Circumstantiae, 2007, 27(4): 660-666.
[8] Ni Z, Wang S, Wang Y. Characteristics of bioavailable organic phosphorus in sediment and its contribution to lake eutrophication in China[J]. Environmental Pollution, 2016, 219: 537-544.
[9] Lin S, Litaker R W, Sunda W G. Phosphorus physiological ecology and molecular mechanisms in marine phytoplankton[J]. Journal of Phycology, 2016, 52(1): 10-36.
[10] Labry C, Delmas D, Herbland A. Phytoplankton and bacterial alkaline phosphatase activities in relation to phosphate and DOP availability within the Gironde plume waters (Bay of Biscay)[J]. Journal of Experimental Marine Biology and Ecology, 2005, 318(2): 213-225.
[11] Nicholson D, Dyhrman S, Chavez F, et al. Alkaline phosphatase activity in the phytoplankton communities of Monterey Bay and San Francisco Bay[J]. Limnology and Oceanography, 2006, 51(2): 874-883.
[12] Zhou L, Tan Y, Huang L, et al. Enhanced utilization of organic phosphorus in a marine diatom: A possible mechanism for aluminum effect under P limitation[J]. Journal of Experimental Marine Biology and Ecology, 2016, 478: 77-85.
[13] Shi Y, Hu H, Ma R, et al. Improved use of organic phosphate bythrough regulation of Zn2+concentrations[J]. Biotechnology Letters, 2004, 26(9): 747-751.
[14] White A E, Watkins-Brandt K, Engle M, et al. Characterization of the rate and temperature sensitivities of bacterial remineralization of dissolved organic phosphorus compounds by natural populations[J]. Frontiers in Microbiology, 2012, 3: 276.
[15] Hessen D O, Hafslund O T, Andersen T, et al. Changes in stoichiometry, cellular RNA, and alkaline phosphatase activity ofin response to temperature and nutrients[J]. Frontiers in Microbiology, 2017, 8: 18.
[16] Lee K H, Jeong H J, Lee K, et al. Effects of warming and eutrophication on coastal phytoplankton production[J]. Harmful Algae, 2019, 81: 106-118.
[17] 赵行行, 刘妍忻, 宋明, 等. 东港赤潮监控区中肋骨条藻生长的主要环境因子影响分析[J]. 环境与发展, 2018, 30(11): 13-15. Zhao Hanghang, Liu Yanxin, Song Ming, et al. Influence of environmental factors on the growth ofin the Donggang red-tide-monitoring area of Liaoning[J]. Environment and Development, 2018, 30(11): 13-15.
[18] 朱根海, 许卫忆, 朱德第, 等. 长江口赤潮高发区浮游植物与水动力环境因子的分布特征[J]. 应用生态学报, 2003(7): 1135-1139.Zhu Genhai , Xu Weiyi , Zhu Dedi, et al. Distribution of phytoplankton and water dynamical environmental factors in high red tide occurrence area of Changjiang River estuary[J]. Chinese Journal of Applied Ecology, 2003(7): 1135-1139.
[19] Harrison P J, Waters R E, Taylor F J R. A broad spectrum artificial sea water medium for coastal and open ocean phytoplankton[J]. Journal of Phycology, 1980, 16(1): 28-35.
[20] Guillard R R L, Ryther J H. Studies of marine planktonic diatoms: I.Hustedt, and(Cleve) Gran[J]. Canadian journal of microbiology, 1962, 8(2): 229-239.
[21] 霍文毅, 俞志明, 邹景忠, 等. 胶州湾中肋骨条藻赤潮与环境因子的关系[J]. 海洋与湖沼, 2001, 32(3): 311-318. Huo Wenyi, Yu Zhiming, Zou Jingzhong, et al. Outbreak ofred tide and its relations to environmental factors in Jiaozhou Bay[J]. Oceanologia et Limnologia Sinica, 2001, 32(3): 311-318.
[22] 黄秀清, 蒋晓山, 王桂兰, 等. 长江口中肋骨条藻赤潮发生过程环境要素分析: 水温, 盐度, DO和pH特征[J]. 海洋通报, 1994, 4: 35-40.Huang Xiuqing, Jiang Xiaoshan, Wang Guilan, et al.[J]. Marine Science Bulletin, 1994, 4: 35-40.
[23] Guillard R R L. Division rates[C]// Stein J R. Handbook of phycological methods: culture methods and growth treatment. Cambridge: Cambridge University Press, 1973: 289-311.
[24] Jeffries D S, Dieken F P, Jones D E. Performance of the autoclave digestion method for total phosphorus analysis[J]. Water Research, 1979, 13(3): 275-279.
[25] Lin X, Zhang H, Huang B, et al. Alkaline phosphatase gene sequence characteristics and transcriptional regulation by phosphate limitation in(Dinophyceae)[J]. Harmful Algae, 2012, 17: 14-24.
[26] Yamaguchi H, Nishijima T, Oda A, et al. Distribution and variation of alkaline phosphatase activity and phosphatase-hydrolyzable phosphorus in coastal seawaters[J]. Bulletin of the Japanese Society of Scientific Fisheries (Japan), 2005, 70(3): 333-342.
[27] Suzumura M, Ishikawa K, Ogawa H. Characterization of dissolved organic phosphorus in coastal seawater using ultrafiltration and phosphohydrolytic enzymes[J]. Limnology and Oceanography, 1998, 43(7): 1553-1564.
[28] Kolowith L C, Ingall E D, Benner R. Composition and cycling of marine organic phosphorus[J]. Limnology and Oceanography, 2001, 46(2): 309-320.
[29] Azam F, Hodson R E. Dissolved ATP in the sea and its utilisation by marine bacteria[J]. Nature, 1977, 267(5613): 696-698.
[30] Young C L, Ingall E D. Marine dissolved organic phosphorus composition: insights from samples recovered using combined electrodialysis/reverse osmosis[J]. Aquatic Geochemistry, 2010, 16(4): 563-574.
[31] Huang B, Ou L, Hong H, et al. Bioavailability of dissolved organic phosphorus compounds to typical harmful dinoflagellateLu[J]. Marine pollution bulletin, 2005, 51(8-12): 838-844.
[32] 李英, 吕颂辉, 徐宁, 等. 东海原甲藻对不同磷源的利用特征[J]. 生态科学, 2005, 24(4): 314-317. Li Ying, Lü Songhui, Xu Ning, et al. The utilization ofto four different types of phosphorus[J]. Ecologic science, 2005, 24(4): 314-317.
[33] 覃仙玲, 陈波, 赖俊翔, 等. 球形棕囊藻北部湾株对不同形态磷源的利用及碱性磷酸酶特性研究[J]. 广西科学, 2018, 25(1): 80-86. Qin Xianling, Chen Bo, Lai Junxiang, et al. Study on the utilization of different phosphorus and alkaline phosphatase characteristics ofcultivated from Beibu Gulf[J]. Guangxi Sciences, 2018, 25(1): 80-86.
[34] 于倩, 王清, 袁泽轶, 等. 不同形态磷源对具槽帕拉藻 () 生长和磷酸酶活性的影响[J]. 海洋与湖沼, 2015, 46(5): 1018-1023. Yu Qian, Wang Qing, Yuan Zeyi, et al. Effects of different phosphorus substrates on growth and phosphatase activity of algae[J]. Oceanologia et Limnologia Sinica, 2015, 46(5): 1018-1023.
[35] Wang Z, Liang Y, Kang W. Utilization of dissolved organic phosphorus by different groups of phytoplankton taxa[J]. Harmful Algae, 2011, 12: 113-118.
[36] Annett R, Habibi H R, Hontela A. Impact of glyphosate and glyphosate-based herbicides on the freshwater environment[J]. Journal of Applied Toxicology, 2014, 34(5): 458-479.
[37] Dabney B L, Patiño R. Low-dose stimulation of growth of the harmful alga,, by glyphosate and glyphosate-based herbicides[J]. Harmful Algae, 2018, 80: 130-139.
[38] Ren L, Wang P, Wang C, et al. Algal growth and utilization of phosphorus studied by combined mono- culture and co-culture experiments[J]. Environmental Pollution, 2017, 220 (part A): 274-285.
[39] Forlani G, Pavan M, Gramek M, et al. Biochemical bases for a widespread tolerance of cyanobacteria to the phosphonate herbicide glyphosate[J]. Plant and Cell Physiology, 2008, 49(3): 443-456.
[40] Tian Y, Mingjiang Z, Peiyuan Q. Combined effects of temperature, irradiance and salinity on growth of diatom[J]. Chinese Journal of Oceanology and Limnology, 2002, 20(3): 237-243.
[41] Luo H, Lin X, Li L, et al. Transcriptomic and physiological analyses of the dinoflagellatereveal non- alkaline phosphatase based molecular machinery of ATP utilisation[J]. Environmental Microbiology, 2017, 19(11): 4506-4518.
[42] Zhang C, Luo H, Huang L, et al. Molecular mechanism of glucose-6-phosphate utilization in the dinoflagellate[J]. Harmful Algae, 2017, 67: 74-84.
[43] Yamaguchi H, Arisaka H, Otsuka N, et al. Utilization of phosphate diesters by phosphodiesterase-producing marine diatoms[J]. Journal of Plankton Research, 2014, 36(1): 281-285.
[44] Healey F P, Hendzel L L. Fluorometric measurement of alkaline phosphatase activity in algae[J]. Freshwater Biology, 1979, 9(5): 429-439.
Effects of temperature and dissolved organic phosphorus on the growth characteristics of
ZHANG Xiao-hua1, 2, 3, LIU Dong-yan4
(1. Key Laboratory of Coastal Zone Environmental Processes and Ecological Remediation, Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, Yantai 264003, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. Department of pharmacy, Binzhou Medical University, Yantai 264003, China; 4. State Key Laboratory of Estuarine and Coastal Research, East China Normal University, Shanghai 200241, China)
The effects of temperature and dissolved organic phosphorus (DOP) on the growth of phytoplankton were investigated in this study. We selectedas the study species and used adenosine triphosphate (ATP), sodium glycerophosphate (SG-P), glucose 6-phosphate (G-6-P) that contains the C-O-P ester bond, and glyphosate (G-P) that contains the C-P bond as DOP sources to measure cell density, total dissolved phosphorus (TDP), and dissolved inorganic phosphorus (DIP) concentrations in the culture medium and alkaline phosphatase (AP) activity in algal cells under four temperature conditions (i.e., 20℃, 23℃, 26℃, and 29℃). Results showed that ATP, SG-P, and G-6-P with a C-O-P ester bond could maintain the growth of, and warming could promote the growth of algal cells in each group. The maximum cell concentrations in the ATP, SG-P, and G-6-P groups reached 176.7 × 104, 218.8 × 104, and 178 × 104cells/mL, respectively. Warming could also accelerate the decrease of TDP concentration in the ATP, SG-P, and G-6-P groups and the increase of DIP concentration in the ATP group. The AP activity in the algal cells of each group was low at different temperatures (<10 fmol pNP/(cell·h)). Compared with that in G-6-P with C-O-P ester bond, the algal cells in G-P with C-O-P bond were lower under the four temperature conditions with maximum cell concentration of 40.7 × 104cells/mL. However, the AP activity in the algal cells was higher than that in the ATP, SG-P, and G-6-P groups (up to 83 fmol pNP/(cell·h)) and decreased with warming. These results showed that different types of DOPs could significantly affect the growth of, among which ATP, SG-P, and G-6-P could be more effectively used by algal cells. Moreover, warming (20~29℃) was an important environmental factor that promoted the growth of algal cells under ATP, SG-P, and G-6-P conditions but did not depend on the AP activity.
; temperature; growth; dissolved organic phosphorus; dissolved inorganic phosphorus; alkaline phosphatase
Feb. 25, 2020
X173
A
1000-3096(2020)11-0036-09
10.11759/hykx20200225001
2020-02-25;
2020-07-20
国家自然科学基金项目(41876127); 国家重点研发计划项目 (2016YFE0133700, 2016YFA0600904)
[National Natural Science Foundation, No. 41876127; Ministry of Science and Technology, Nos. 2016YFE0133700, 2016YFA0600904]
张小华(1980-), 女, 山东荣成人, 博士研究生, 研究方向为藻类生理学, E-mail: 17062997@qq.com; 刘东艳,通信作者, 研究员, 博士生导师, E-mail: dyliu@sklec.ecnu.edu.cn
(本文编辑: 康亦兼)
