5-氟尿嘧啶对结肠癌干细胞干性和增殖的影响及相关机制
2019-12-20蔡尚党蔡满地娄宁郭艳萍殷涛
蔡尚党 蔡满地 娄宁 郭艳萍 殷涛
(1河南省中医院肛肠科,河南 郑州 450002;2河南大学医学院临床医学系;3中山大学附属肿瘤医院ICU;4河南省人民医院病理科;5华中科技大学同济医学院附属协和医院胰腺病研究所)
结肠癌是世界范围内第三常见的癌症,每年造成约70万人死亡〔1~3〕。在过去的二十年中,该病在发展中国家的发病率增加了2~4倍〔4,5〕,手术切除和辅助化疗是治疗结肠癌的主要方法,然而原发性或获得性药物抗性是该病成功治愈的主要挑战〔6〕。因此,亟待开发新的治疗策略或新的辅助药物以增强该病的治疗效果。5-氟尿嘧啶(FU)是一种细胞生长抑制剂,是最常用的治疗结肠癌的药物,然而,对晚期结肠癌5-FU化疗的有效率仅有20%左右,5-FU和如伊立替康及奥沙利铂等新的化疗药物的联合使用提高了5-FU对结肠癌的治疗效果〔7〕,在5-FU的代谢途径中,包括胸苷酸合成酶、甲基四氢叶酸还原酶、二氢嘧啶脱氢酶和胸苷磷酸化酶在内的四种酶被认为与5-FU的治疗敏感性和抗性密切相关,然而,许多研究结果相互矛盾〔8〕。最近,胰岛素样生长因子信号通路被证明对于结肠癌的发育至关重要,其能通过激活如PI3K/Akt等存活通路而有利于结肠癌细胞的存活、侵染、转移和化疗抗性〔9~12〕。本研究探讨5-FU对结肠癌干细胞(CCSCs)干性和增殖的影响及可能机制。
1 材料与方法
1.1CCSCs 标记CD133、CD44和ALDH1b1阳性的CCSCs购自美国Celprogen公司。为维持上述细胞的未分化状态,培养过程中使用CCSCs生长培养基和特异性包被的细胞培养皿,细胞培养于37℃,5%CO2湿润培养箱中,当细胞达到80%汇合度时,用于实验。所有实验所用细胞代次低于10代。细胞分为阴性对照组、DMSO组、舒林酸组、5-FU组。
1.2主要仪器及试剂 5-FU(Sigma,美国),兔抗人Bax、Bcl-2、cyclinD1和c-Myc抗体,鼠抗人α-tubulin抗体(Sigma,美国),兔抗人细胞色素c抗体(Sigma,美国),兔抗人Histone抗体(Cell Signaling Technology,美国),辣根过氧化物酶(HRP)标记羊抗兔/鼠IgG(Abcam,美国),嘌呤霉素(Sigma,美国),p53特异性shRNA质粒(Santa Cruz,美国),5-溴-2'-脱氧尿苷(BrdU)检测试剂盒(Cell Signaling Technology,美国),细胞死亡检测试剂盒(Roche,德国),电化学发光(ECL)显色试剂盒(碧云天生物技术研究所,中国),CO2培养箱(SANYO,日本),相差显微镜(Olympus,日本)。
1.3介导p53敲低表达的重组慢病毒短发夹RNA(shRNA)载体 CCSCs接种含靶向p53基因特异性shRNA的慢病毒颗粒,在含有5 μg/ml凝聚胺的CCSC培养基中培养的CCSCs接种10 MOI的慢病毒,慢病毒感染后24 h,更换新鲜培养基,细胞继续培养2 d后,在含有7.5 μg/ml嘌呤霉素的选择性培养基中加压筛选5 d。
1.4细胞增殖检测 BrdU检测试剂盒检测细胞活性,正常CCSCs或shRNA-p53 CSCs以1×105个/孔的密度铺板于12孔板中,培养24 h后,更换为不含血清的CCSCs培养基,并经10 μg/ml舒林酸或2.5 μg/ml 5-FU处理,处理24 h后,检测BrdU含量,实验重复3次,结果以均值表示。
1.5末端转移酶dUTP切口末端标记检测 使用荧光标记的核苷酸和末端脱氧核苷酸转移酶(TdT)鉴定细胞凋亡产生的DNA片段。6×104个正常CCSCs或shRNA-p53-CCSCs接种于玻璃载玻片上,经10 μg/ml舒林酸或2.5 μg/ml 5-FU处理12 h后,根据说明书的要求,使用原位细胞死亡检测试剂盒定量检测细胞凋亡情况,同时用不含TdT的孵育液孵育的样品作为阴性对照,计数5个视野内的染色细胞数目,计算凋亡细胞比例,每次至少计数50个细胞,实验重复3次,结果以均值表示。
1.6球体形成实验 1×104个CCSCs用干细胞特异性的不含血清的培养基培养于超低吸附性的6孔培养板中,然后将细胞置于正常细胞培养箱中,细胞接种于6孔板后6 h,加入10 μg/ml舒林酸或2.5 μg/ml 5-FU,处理10 d后,使用相差显微镜计数球体数量,实验重复3次,结果以均值表示。
1.7Western印迹检测 正常CCSCs或shRNA-p53-CCSCs以3.0×105个/孔的密度铺于6孔板中,用CCSCs培养基培养,24 h后,细胞转移至无血清培养基中孵育18 h,提取细胞样品的总蛋白,并将总蛋白转移至PVDF膜上,孵育一抗,一抗稀释比例为:兔抗人Bax、Bcl-2、cyclinD1和c-Myc抗体1∶1 000稀释,鼠抗人α-tubulin抗体1∶5 000稀释,兔抗人细胞色素c抗体1∶1 000稀释,兔抗人Histone抗体1∶2 000稀释。一抗4℃孵育过夜,然后,HRP标记的羊抗兔/鼠二抗以1∶2 000的比例稀释,室温孵育2 h,目的蛋白用增强型ECL化学发光液显色,并经BandScan软件进行灰度扫描分析,α-tubulin作为总蛋白检测时上样的内参蛋白,而Histone作为细胞核蛋白检测时上样的内参蛋白。
1.8统计学方法 采用GraphPad Prism5软件进行t检验或单因素方差分析。
2 结 果
2.15-FU抑制CCSCs增殖并诱导凋亡 对阴性照组相比,5-FU的浓度为2.5 μg/ml时,正常CCSCs的增殖速率下降了52%,而shRNA-p53-CCSCs增殖速率下降了73%,10 μg/ml的舒林酸处理导致正常CCSCs增殖速率下降了47%,然而,敲低CCSCs中p53表达后,舒林酸对CCSCs增殖的抑制效应较未敲低时强(51%),表明舒林酸对CCSCs的增殖抑制效应具有p53依赖性。使用TUNEL法检测细胞凋亡情况结果表明,5-FU处理后,正常CCSCs凋亡率为31%,shRNA-p53-CCSCs凋亡率为42%,表明5-FU能以p53非依赖性方式抑制结肠CSCs增殖,见表1,表2,图1。
2.25-FU抑制CCSCs球体形成能力 CCSCs经2.5 μg/ml 5-FU或10 μg/ml舒林酸处理后,5-FU和舒林酸均显著抑制CCSCs球体形成,与阴性对照组相比,5-FU和舒林酸处理组球体体积显著下降,结果表明,5-FU除了能抑制CCSCs增殖并促进凋亡外,其还能抑制CCSCs的自我更新能力,见图2。

表1 正常CCSCs与转染shRNA-p53对细胞增殖的影响
与阴性对照组比较:1)P<0.05,下表同

表2 正常CCSCs与转染shRNA-p53对细胞凋亡率的影响

图1 5-FU以p53非依赖性方式抑制CCSCs增殖并诱导细胞凋亡(×300)

图2 5-FU抑制CCSCs球体形成(×300)
2.35-FU促进线粒体介导的凋亡途径相关蛋白Bax/Bcl-2和细胞色素c表达 正常CCSCs和shRNA-p53-CCSCs经5-FU处理后,细胞裂解产物用于Western印迹检测,5-FU处理后正常CCSCs中Bax/Bcl-2比例显著升高,同时,5-FU处理也促进了正常CCSCs和shRNA-p53-CCSCs中细胞色素c表达,表明5-FU可能通过线粒体介导的凋亡途径诱导CCSCs凋亡,尽管舒林酸诱导CCSCs凋亡,但未使细胞中Bax/Bcl-2比例,明显改变但可以促进细胞色素c表达。见图3、表3。

1~4:阴性对照组,DMSO组,舒林酸组,5-FU组,下图同图3 5-FU上调线粒体介导的细胞凋亡途径蛋白表达
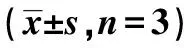
表3 5-FU上调线粒体介导的细胞凋亡途径蛋白表达
与阴性对照组比较:1)P<0.01,2)P<0.001
2.45-FU抑制Wnt信号通路活化 正常CCSCs和shRNA-p53-CCSCs中,5-FU抑制胞质和胞核中β-连环蛋白表达,且其抑制效应显著强于舒林酸的抑制效应(图4,表4)。5-FU能够抑制β-连环蛋白通路下游靶蛋白c-Myc和cyclinD1的表达(图5,表5),上述结果表明5-FU能抑制β-连环蛋白的核转位,进而导致CCSCs生长受到抑制。

图4 5-FU抑制正常CCSCs和shRNA-p53-CCSCs胞质和胞核内β-连环蛋白表达

表4 5-FU抑制正常CCSCs和shRNA-p53-CCSCs胞质和胞核内β-连环蛋白表达
与阴性对照组比较:1)P<0.05,2)P<0.01,3)P<0.001,下表同

图5 5-FU抑制正常CCSCs和shRNA-p53-CCSCs中β-连环蛋白的靶蛋白c-Myc和cyclinD1表达

表5 5-FU抑制正常CCSCs和shRNA-p53-CCSCs中β-连环蛋白的靶蛋白c-Myc和cyclinD1表达
3 讨 论
本研究表明5-FU可能以非p53依赖性方式发挥作用,5-FU也能上调与线粒体介导的凋亡途径相关的蛋白表达,同时下调与Wnt/β-连环蛋白信号通路有关的蛋白表达,5-FU的浓度为2.5 μg/ml时抑制CCSCs增殖并诱导细胞凋亡;然而舒林酸诱导的上述效应与p53有关。以往研究表明,在AOM诱导的p53功能缺陷型鼠模型中,舒林酸的功能具有p53依赖性〔14~16〕,与p53+/+小鼠相比,舒林酸在p53-/-缺陷型小鼠中不能恢复急性凋亡反应,上述现象是至关重要的,因为在结肠癌晚期或转移性阶段,p53会发生突变〔17〕。5-FU诱导CCSCs发生凋亡的同时,Bax/Bcl-2比例和细胞色素c的表达也升高。Bax是一种促凋亡蛋白,其能结合于Bcl-2并促进细胞色素c释放,细胞色素c是线粒体介导的凋亡途径的重要分子〔18,19〕。这些结果表明,5-FU通过线粒体介导的凋亡途径诱导CCSCs凋亡,本研究结果也证实5-FU抑制球体形成,因为克隆球体的形成是干细胞干性的重要特征,本研究表明5-FU具有潜在的靶向CCSCs自我更新的潜能。本文中5-FU处理导致正常CCSCs和shRNA-p53-CCSCs中β-连环蛋白的表达受到抑制,β-连环蛋白的稳定表达及其在胞核中的积累伴随着c-Myc和cyclinD1的转录激活,这些基因的转录激活能促进细胞增殖而增加致癌性〔20~22〕,5-FU处理后的CCSCs能抑制c-Myc和cyclinD1表达,且这种抑制效应不依赖于p53。
CCSCs的一些特征可能解释5-FU抑制干细胞干性的原因,干细胞表达高水平的干性因子,其中包括肿瘤蛋白c-Myc〔23〕,该蛋白在CCSCs中过度表达〔24〕。本研究表明,体外5-FU能抑制CCSCs中Wnt效应因子β-连环蛋白和下游靶蛋白c-Myc和cyclinD1表达。因此,相对于可分化细胞的致癌性变化,干细胞致癌性变化,如β-连环蛋白积累,可能对5-FU诱导的凋亡更敏感。
