旋毛虫赖氨酸乙酰化修饰组学的分析研究
2019-11-05刘晓雷王学林刘明远
刘 蕾,杨 勇,高 欣,刘晓雷,唐 斌,王学林,刘明远,白 雪
近年来,旋毛虫的基因组、转录组、表观基因组和蛋白质组的研究为深入了解旋毛虫生物学、旋毛虫-宿主相互作用和发病机理奠定了坚实的分子基础[1-2]。然而,对于旋毛虫蛋白质生物合成过程中或之后发生的转录后修饰(Post Transcriptional Modification, PTMs)至今没有相关报到,限制了旋毛虫侵袭过程中关键蛋白分子的功能研究。PTMs可以调控蛋白质的性质,包括折叠、稳定性、定位、结合亲和力和活性,它们在多种细胞途径和过程中发挥重要作用。由于它们在调节蛋白质功能及代谢和基因转录等细胞过程中发挥关键作用,因此,蛋白质PTMs的研究在后基因组时代变得更加重要[3]。迄今为止,已经鉴定了超过461种不同的PTM,例如磷酸化、甲基化、泛素化、丙二酰化、琥珀酰化、糖基化和乙酰化,其在许多原核和真核蛋白质的功能调节中发挥作用[4]。这些PTM形成一个复杂的监管系统,用于在各种条件下细胞的动态调控。
研究表明,乙酰化可以调节寄生虫的发育和抗原变异,这主要是由强选择压力决定的[5]。旋毛虫的完整生命周期发生在单一宿主体内,可分为3个主要阶段:肌幼虫期(ML),成虫期(AD)和新生幼虫期(NBL)。它们占据了2个不同的细胞内龛:肠细胞和骨骼肌细胞[6]。在这些发育阶段,旋毛虫已进化出适应不同环境并成功建立感染(例如寄生虫表面抗原的变异)的策略。因此,旋毛虫不同发育阶段的乙酰化蛋白质组的研究,有助于揭示赖氨酸乙酰化在调节旋毛虫发育转变及其在不同生活环境的选择压力下成功寄生中的作用。本研究第一次大规模分析旋毛虫肌幼虫期蛋白质中赖氨酸乙酰化程度,不仅可以拓展我们对蠕虫蛋白乙酰化的知识,而且为设计新的控制旋毛虫病的疫苗及药物靶点提供了重要信息。
1 材料与方法
1.1 材 料
1.1.1细胞、实验动物与旋毛虫虫种 Wistar大鼠,购自长春生物制品研究所,用于旋毛虫传代保种及旋毛虫肌幼虫收集。中国河南猪旋毛虫分离株T.spiralis(ISS534),由本室传代保种。
1.1.2主要试剂 胃蛋白酶、胰蛋白酶、三氟乙酸购自Sigma公司;三氯乙酸(TCA)、碘乙酰胺(IAA)、二硫苏糖醇(DTT)购自Merck公司;蛋白酶抑制剂购自碧云天生物技术有限公司;2-D Quant试剂盒购自GE Healthcare公司;EDTA购自索莱宝公司;抗乙酰赖氨酸抗体珠购自PTM Biolabs公司;C18 ZipTips购自美国Millipore公司。
1.2 方 法
1.2.1旋毛虫收集和蛋白质提取 旋毛虫(ISS534)在Wistar大鼠中通过连续传代保种。取旋毛虫ISS534感染后35 d(dpi)的Wistar大鼠1只,脱颈致死,37 °C人工消化液(10 g/L胃蛋白酶和10 mL/L浓盐酸)消化胴体2 h后,采用自然沉淀的方法收集感染性肌幼虫(ML)随后进行虫体蛋白的提取。用含有8 mol/L尿素,10 mmol/L二硫苏糖醇(DTT)和0.1%蛋白酶抑制剂混合物的裂解缓冲液裂解旋毛虫后,进行超声处理,并通过20 000×g、4 ℃离心10 min除去细胞碎片。离心后的上清液用预冷的15%三氯乙酸(TCA)在-20 ℃沉淀2 h后离心。沉淀用预冷的丙酮洗涤3次,随后溶解在含有8 mol/L尿素和100 mmol/L NH4CO3(pH 8.0)的缓冲液中,2-D Quant试剂盒(GE Healthcare)测量蛋白质浓度。
1.2.2样品制备和高效液相色谱(HPLC)分离肽段
用10 mmol/L DTT和20 mmol/L碘乙酰胺(IAA)对蛋白进行还原和烷基化处理,并用胰蛋白酶(1∶50质量比)过夜消化。为确保完全消化,以1∶100(质量比)再次消化4 h。随后使用高pH反相HPLC将胰蛋白酶肽分成6个部分并将分离的肽在Speed Vac(Thermo Scientific)中干燥,保存在-80 ℃,用于乙酰化富集分析。
1.2.3亲和富集和液相色谱电喷雾串联质谱(LC-ESI-MS/MS)分析 将分级分离的肽溶解在含有100 mmol/L NaCl,1 mmol/L EDTA,50 mmol/L Tris-HCl和0.5% NP-40(pH8.0)的NETN缓冲液中,并在4 ℃下与抗乙酰赖氨酸抗体珠(PTM Biolabs)孵育过夜。用NETN缓冲液和去离子H2O洗涤后,0.1%三氟乙酸洗脱肽段,真空干燥。在HPLC/MS/MS 分析前用C18 ZipTips(Millipore)清理洗脱的肽段。随后,利用EASY-nLC 1000超高效液相色谱(UPLC)系统上在Q Exactive 质谱仪(Thermo Fisher Scientific)对所富集的肽段进行LC-ESI-MS/MS分析,所得到的MS/MS数据使用MaxQuant软件和整合的Andromeda搜索引擎(v.1.5)进行处理。使用以下参数在UniProt上的T.spiralis数据库对串联质谱进行查询[7]:胰酶酶解肽段在谱图搜库时使用最大4个漏切位点,每个肽段最大4个修饰个数。Mass error 设置为母离子10-6,碎片离子0.02 Da。指定Cys上的氨甲酰甲基化为固定修饰,指定Met上的氧化,Lys上的乙酰化和蛋白质N-末端上的乙酰化为可变修饰。蛋白、肽段和乙酰化的FDR阈值设置为1%, 最小肽段长度为7。
1.2.4生物信息学分析 利用UniProt-GOA数据库对鉴定的蛋白进行GO(gene ontology)功能注释。利用Wolfpsort对每个乙酰化的蛋白进行亚细胞定位注释。在对目标蛋白质集合进行GO注释或KEGG通路注释的富集分析时,通过Fisher精确检验(Fisher’s Exact Test),来分析乙酰化蛋白GO或KEGG通路蛋白质富集度的显著性水平,P<0.05为差异有统计学意义。
2 结 果
2.1旋毛虫赖氨酸乙酰化蛋白的鉴定与分析 分析表明分子量偏差的分布接近零,大部分≤10-6(图1A)。此外,所有赖氨酸乙酰化肽的长度都在7到37个氨基酸之间,这与胰蛋白酶消化的肽一致(图1B)。这些结果表明样品制备方法是正确的,并且从MS获得的修饰的肽数据是高度准确的。本实验共鉴定出1 592个乙酰化蛋白及3 872个Kac位点,旋毛虫的乙酰化蛋白含有不同数目的乙酰化位点,范围从1至47(图1C),约一半(795/1 592)仅含有一个乙酰化位点,平均每个蛋白2.4个位点。

(A)所有鉴定的肽的质量误差分布;(B)乙酰化肽的长度分布;(C)乙酰化修饰蛋白修饰位点的分布图1 组学基于的旋毛虫赖氨酸乙酰化位点的鉴定 Fig.1 Proteome-wide identification of lysine acetylation sites in T. spiralis
2.2旋毛虫赖氨酸乙酰化(Kac)保守性分析 在旋毛虫的1 592个乙酰化蛋白中,408个(35.5%)和87个(30%)的蛋白分别与日本血吸虫和弓形虫乙酰化蛋白存在同源物,479个(35.2%)和471个(36.3%)的蛋白分别与小鼠和人类乙酰化蛋白存在同源物。159个(31.6%)和190个(39.8%)蛋白分别与酿酒酵母和白色念珠菌存在同源物,而仅43个(12.3%)与大肠杆菌乙酰化蛋白存在同源物(图2)。
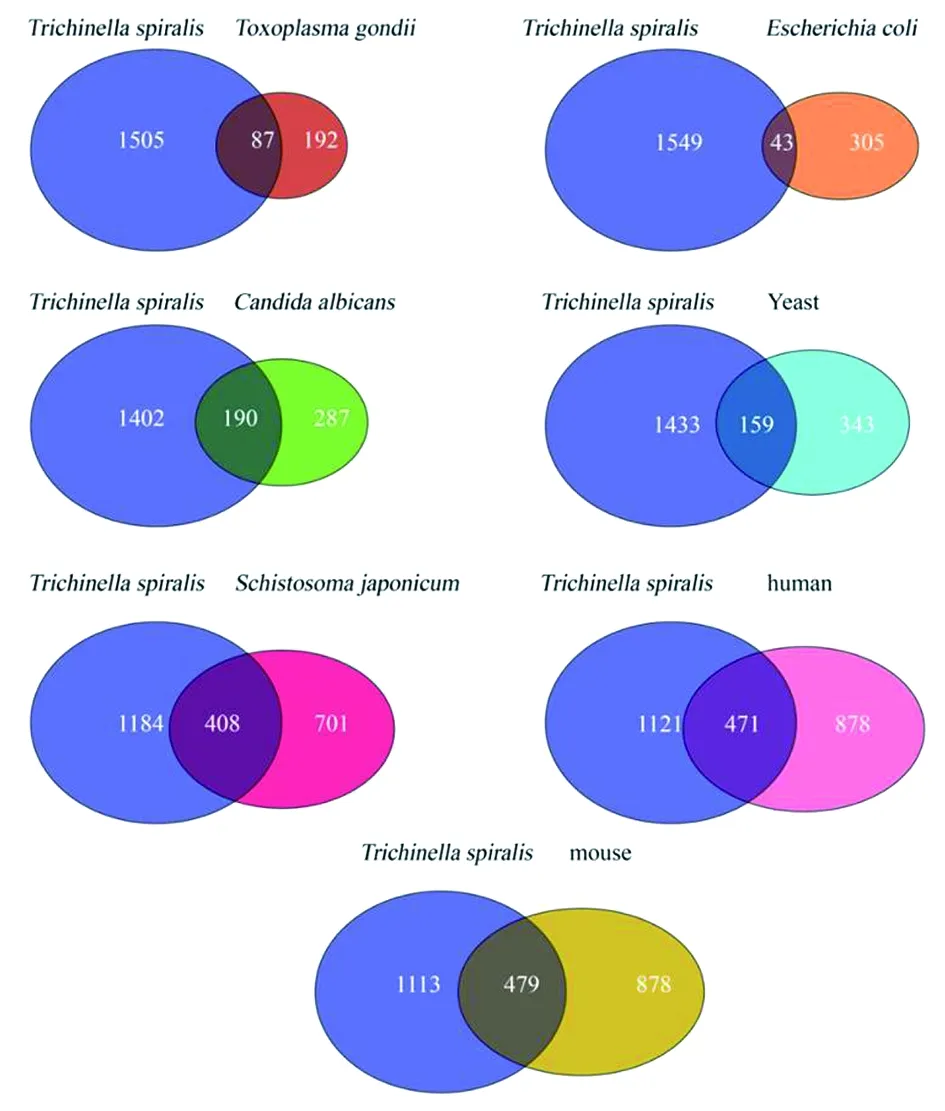
图2 分析比较旋毛虫乙酰蛋白质组与大肠杆菌,芽殖酵母(酿酒酵母),白色念珠菌,刚地弓形虫,日本血吸虫,小鼠和人类的乙酰化蛋白质组 Fig.2 Analysis comparing the T. spiralis acetyl proteome with those of E. coli, budding yeast (S. cerevisiae), C. albicans, T. gondii, S. japonicum, mouse, and human. Venn diagram describing the relationships among lysine-acetylated proteins from these species.
2.3乙酰化蛋白质在旋毛虫中的功能注释和细胞定位 我们对乙酰化的蛋白质组学数据进行了基因本体(gene ontology,GO)分析。细胞组件聚类分析显示旋毛虫乙酰化蛋白质主要分布在细胞(37%),细胞器(24%),大分子复合物(22%)和膜(13%)中。分子功能的聚类分析显示,最大一组乙酰化蛋白参与各种靶标结合,占所有鉴定乙酰化蛋白质的40%(图3B)。本文的结果与以前的研究结果一致,其中相当数量的具有催化活性的蛋白被乙酰化。GO生物过程的聚类分析显示乙酰化蛋白主要参与细胞和代谢过程,分别占识别蛋白的29%和28%(图3C)。本文也分析了1 592个乙酰化蛋白质在旋毛虫中的亚细胞定位(图3D),结果表明大多数鉴定的Kac蛋白质定位于细胞质(38%),其次是细胞核(17%)和线粒体(12%)。这一结果与生物过程分析结果一致,表明大部分乙酰化蛋白质参与细胞代谢和蛋白质合成。重要的是,还发现一些乙酰化蛋白质是细胞质膜(12%)或细胞外蛋白质(12%)定位,其可能调节旋毛虫表面蛋白和分泌蛋白,影响旋毛虫与宿主细胞之间的相互作用。
2.4功能富集分析 进一步对乙酰化数据进行了GO富集分析。基于生物过程的GO富集分析显示,乙酰化蛋白质富集于生物合成和代谢过程(图4A)。同时还进行了KEGG富集分析,数据表明,旋毛虫(T.spiralis)的乙酰化蛋白质组在蛋白质生物合成和3种类型的生物大分子的代谢中高度富集(图4B)。
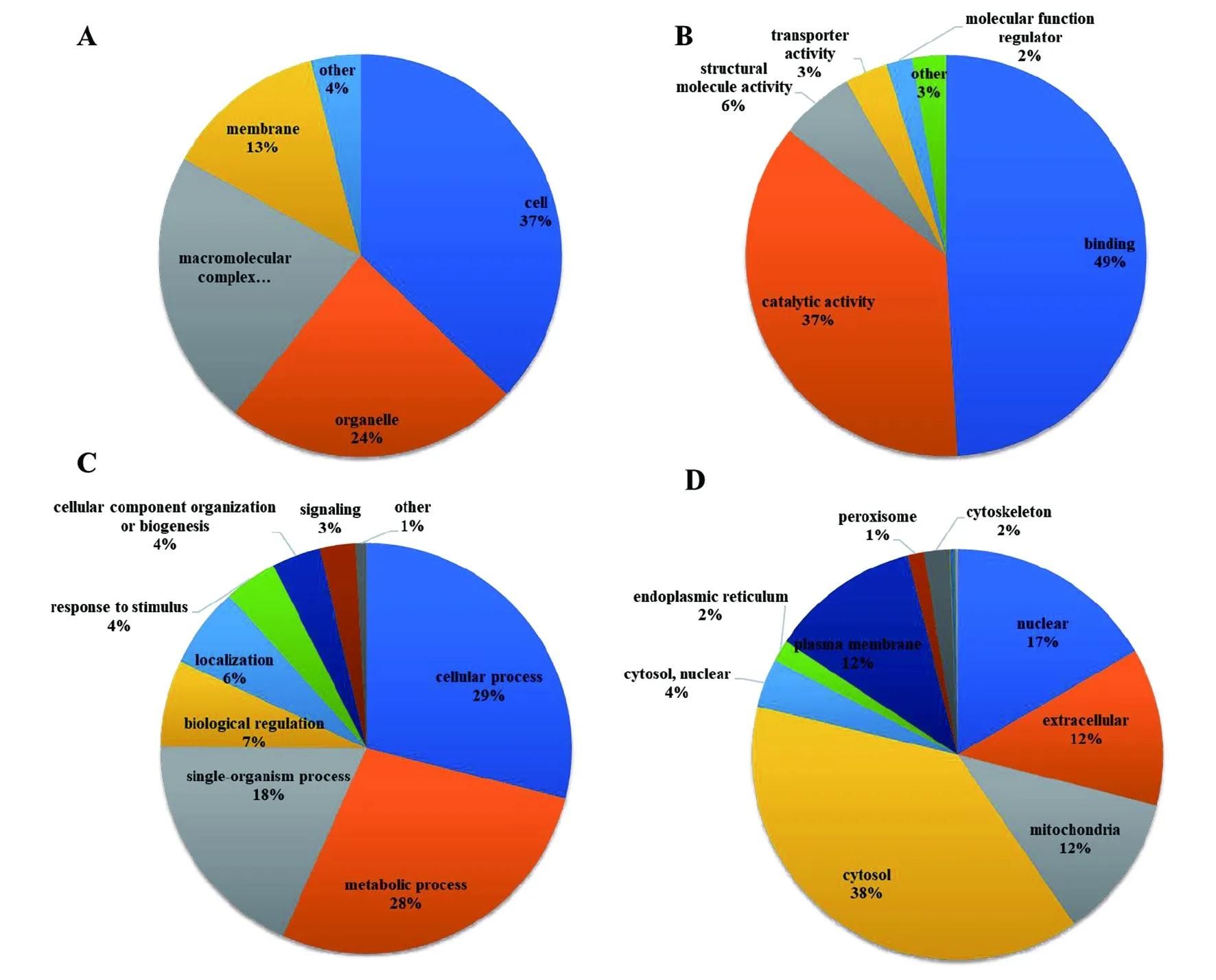
(A)根据细胞成分分类的乙酰化蛋白质;(B)根据生物学过程分类的乙酰化蛋白质;(C)根据分子功能分类的乙酰化蛋白质;(D)鉴定的乙酰化蛋白的亚细胞定位。图3 基因本体功能分类和亚细胞定位信息鉴定乙酰化蛋白质Fig.3 Gene Ontology functional classification and subcellular location information of identified acetylated proteins

(A)就分子功能,细胞组分和生物过程而言,乙酰化蛋白质的基于GO的富集分析;(B)KEGG途径富集分析。图4 旋毛虫乙酰化蛋白的富集分析 Fig.4 Enrichment analysis of acetylated proteins in T. spiralis
3 讨 论
赖氨酸乙酰化(Kac)是一种动态可逆且高度保守的翻译后修饰,是乙酰化转移酶 (lysineacetyltransferase, KAT) 和去乙酰化酶 (lysine deacetylase, KDAC)催化乙酰辅酶A(acetyl-CoA)提供的乙酰基团到蛋白质分子赖氨酸残基的可逆转移过程。Kac首先在真核组蛋白中被鉴定,它参与调节染色质结构和DNA转录[7]。除了在组蛋白中的作用外,Kac还存在于许多非核蛋白中参与不同的细胞生理过程,如细胞骨架动力学、凋亡、自噬和代谢[8]。另外,乙酰化也影响病原菌的毒力和抗生素抗性[9]。尽管Kac在许多生物体中发挥重要作用,但是迄今为止,蠕虫赖氨酸乙酰化的研究相对有限。旋毛虫作为重要的寄生蠕虫,其基因组的测序揭示了许多乙酰转移酶和脱乙酰酶同源物的存在,表明Kac可能在旋毛虫的发育和代谢中发挥重要作用[10]。Kac过程依赖氨酸乙酰转移酶和去乙酰酶(KDAC)调节,比其他PTMs的调控范围广泛并且更有可比性。KDAC已被确定为开发新型抗寄生虫治疗的药物靶点,并且一些KDAC配体显示出治疗寄生虫病的潜力[11]。
本项究中,我们利用抗赖氨酸乙酰化抗体富集技术结合高度灵敏的质谱鉴定在组学层面分析旋毛虫中的赖氨酸乙酰化程度,发现旋毛虫1 592种蛋白质872个赖氨酸位点发生了乙酰化修饰,超过了已报道的日本血吸虫中Kac水平[12],表明Kac在旋毛虫发育、寄生中的重要作用。与以前的研究结果一致,大量的乙酰化蛋白质涉及代谢和生物合成过程。进一步分析表面,大约一半的乙酰化蛋白存在2个以上的修饰位点。乙酰化转移酶通过修饰非组蛋白的多重位点利用不同的机制调节从细胞信号、蛋白转录、降解等不同的细胞进程[13]。这是旋毛虫蛋白的多重位点乙酰化修饰在调节蛋白功能中的作用需要进一步的研究。大量的证据表明,乙酰化修饰可能是进化保守在不同的种属[14]。
我们对乙酰化的蛋白进行了保守性分析,结果表明旋毛虫和大肠杆菌之间的同源性最低,这可能反应了真核与原核生物间不同的细胞结构及生物进程。而旋毛虫和白色念珠菌之间的同源性最高,这可能与它们之间的遗传性相关,也可能他们有着相似的亚细胞分布。在其他真核生物和原核生物中也报道了乙酰化蛋白质在各种途径(包括翻译,转录和代谢)中高度富集,我们的GO和KEGG富集结果进一步表明了Kac在各种生物体中的重要作用。乙酰化修饰在糖代谢已被表面发挥着重要的作用在调节寄生虫生长发育。研究发现在疟原虫糖酵解相关酶中仅仅葡萄糖六磷酸异构酶(GAPDH)没有发生乙酰化[15],而在血吸虫中,全部的糖酵解发生了乙酰化修饰。我们的研究发现,仅仅GAPDH没有发生乙酰化修饰。结果表明与其他寄生虫一样,乙酰化修饰在旋毛虫糖代谢调节中同样发挥着重要的作用。已表明吞噬在寄生虫获取营养,细胞增殖和寄生虫毒力中发挥重要的作用。有趣的是,根据功能富集分析,29个乙酰化蛋白与吞噬作用相关,表明赖氨酸乙酰化在这一过程中的重要作用[16]。总之,本研究首次对旋毛虫乙酰化的报道,提供了旋毛虫乙酰化图谱,为进一步探索赖氨酸乙酰化在蠕虫的寄生作用提供了重要数据。
利益冲突:无
