不同修复模式边坡土壤微生物量季节动态
2019-10-25张琳瑶杜祥运刘大翔
张琳瑶 杜祥运 刘大翔


摘要:通过对四川省向家坝工程扰动区3种不同坡位土壤微生物量碳(MBC)和氮(MBN)含量进行季节动态监测,对其主要影响因子进行相关性分析,揭示土壤微生物量的季节变化特征及其主要影响因子。结果表明:3种边坡MBC、MBN含量排序为天然林>植被混凝土>客土;土壤MBC、MBN表现出较高的空间异质性;土壤MBC含量与有机碳含量、全氮含量及含水量呈极显著正相关(P<0.01),土壤MBN含量与有机碳、全氮及微生物量碳含量呈极显著正相关(P<0.01),与pH值呈极显著负相关(P<0.01)。研究区土壤微生物量的季节变化可能与气候条件、植物生理过程、土壤环境因子、养分资源有关。
关键词:修复模式;土壤微生物量碳;土壤微生物量氮;空间异质性;季节动态
中图分类号: S154.3;X53 文献标志码: A 文章编号:1002-1302(2019)13-0320-05
在土壤生态系统中,土壤微生物量尽管只占小于 5 000 μm3 土壤体积,但既是养分的库,又是养分的源,作为土壤中极为活跃的因子,参与了有机质分解以及腐殖质的形成等诸多生态过程,对陆地生态系统的能量流动以及碳氮循环有着重要的意义,并且影响了生态系统中的植物营养土壤结构和土壤肥力等变化,能综合反映土壤质量状况[1-2]。在土壤碳氮循环过程以及对自然和人为干扰的响应中,土壤微生物量能准确反映出微生物群落在土壤中的实际意义和作用潜力,被认为是土壤环境质量的灵敏生物指标[3]。土壤生态系统中土壤微生物量季节性波动模式复杂,受多种生态因子综合作用的影响,主要与土壤中可利用碳和养分资源的限制、植物生长节律、土壤温度及土壤湿度等因素有关[4-5]。而在边坡生境中,地形因素可以通过影响土壤的微气候、土壤的理化性质、植被的生长以及地下碳输入等途径影响土壤微生物的生长及生物量[6-7]。
近年来,我国大力加强大型水利水电工程的建设,而在大型工程建设的同时伴随如大规模土石开挖形成高陡边坡、碎渣堆弃、改变地形及掩埋原有植被等,解决此类生态环境问题一般须要采用人工修复方式[8]。而我国西南地区是典型的干热河谷地带,水利资源丰富,山高坡陡沟壑纵横,是国家水利水电重点规划与开发区。大型水利水电工程的兴建必然会对工程区极其脆弱的生态环境产生严重的干扰,导致生态系统退化、物种多样性丧失、滑坡、水土流失等一系列严重的生态环境问题[8-9]。目前,常采用人工方式修复被毁损的陆地植被[10]。相关研究大多集中于先锋物种生理特性[11]、修复后不同修复土壤在同一地区肥力的对比分析[8]和生态环境监测与评价[12],对恢复生态系统微生物生物量的研究主要集中在农业措施和环境条件[13-15]上,而对工程扰动区不同修复土壤中微生物量季节动态研究鲜有报道。
因此,本研究选取西南地区大型水利水电站——向家坝水电站工程扰动区不同恢复模式边坡,进行土壤微生物碳氮含量的季节动态研究,以期了解不同生境土壤微生物量碳、氮含量及其季节变化,并从土壤微生物量角度分析不同生境土壤肥力状况,为工程扰动区人工植被修复技术的筛选和应用提供一定的理论基础。
1 研究地点与研究方法
1.1 研究地域概况
金沙江向家坝水电站位于滇东北最北端,地处金沙江与横江交汇的三角地带,地理坐标位于28°22′15″~28°39′16″N、104°03′27″~104°25′25″E之间。该地区属南亚热带干热季风气候,是典型的干热河谷地区,干热少雨,干湿季分明。年均气温21~23 ℃;年均降水量896.2 mm,其中6—11月降水量占全年降水量的90%以上,年蒸发量在2 500~3 000 mm;年均相对湿度为81%左右;年日照时数2 200 h左右。
1.2 研究方法
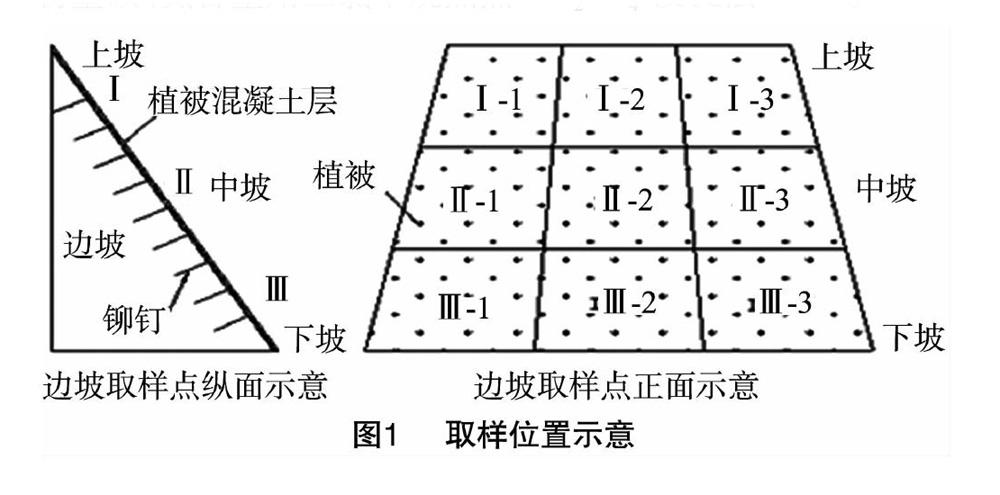
于2014年3—12月期间,选取金沙江向家坝水电站工程扰动区的1个天然林边坡作(对照)、1个植被混凝土基材边坡、1个客土喷播边坡这3个代表性边坡样地为研究对象(表1)。植被混凝土边坡2005年施工,人工修复土壤为当地开挖原土壤、腐殖质、复合肥、水泥、混凝土绿化添加剂和保水剂的混合物[16]。客土喷播边坡2004年施工,人工修复土壤为当地开挖原土壤、腐殖质及复合肥的混合物[17]。
1.3 数据处理
数据经Excel 2007整理后,采用SPSS统计分析软件进行单因素方差分析(ANOVA)方法分析,再用Pearson分析方法进行相关性分析。
2 研究结果与分析
2.1 土壤微生物量碳时间动态
如图2所示,天然林、植被混凝土和客土喷播土层下坡位土壤微生物量碳(MBC)含量的变化具有明显的季节动态(P<0.05),在整个生长季内,天然林、植被混凝土和客土喷播MBC含量均呈现先上升后下降的变化动态,在9月时出现峰值,四季MBC含量变幅分别为462.69~694.04、210.35~567.85、168.25~420.63 mg/kg,除9月外,天然林MBC在四季均显著高于植被混凝土、客土喷播,是植被混凝土、客土喷播的1.22~2.50、1.65~3.13倍。
如图3所示,中坡位土壤MBC含量的变化具有明显的季节动态(P<0.05),3种边坡生境MBC含量呈现先上升后下降的趋势。其中,天然林MBC峰值出现在6—9月;植被混凝土峰值出現在9—12月;客土喷播出现在9月。在整个生长季内,天然林中坡位土壤MBC含量(231.35~630.95 mg/kg)是植被混凝土(210.32~420.63 mg/kg)、客土喷播(168.25~483.73 mg/kg)含量的1.10~2.00、 1.30~2.30倍。
如图4所示,上坡位土壤MBC含量的变化具有明显著的季节动态(P<0.05),3种边坡生境MBC含量呈现先上升后下降的趋势,其中天然林和指标混凝土在9月出现峰值,客土喷播峰值出现在9—12月。在整个生长季内,植被混凝土上坡位土壤MBC含量是天然林、客土喷播含量的1.05~1.25、1.33~1.67倍。
将各边坡所对应不同坡位、不同季节的土壤MBC含量求和后计算,得出不同类型边坡土壤MBC含量的平均值。如图5所示,天然林、植被混凝土和客土喷播土壤MBC含量均值分别为476.72、368.05、280.42 mg/kg。天然林的MBC坡位均值最高,分别是混凝土、客土的1.30、1.70倍;客土喷播的MBC含量均值最低,是植被混凝土的0.76倍。
2.2 土壤微生物量氮时间动态
3种边坡生境下坡位土壤MBN含量的变化具有明显的季节动态,结果如图6所示,在整个生长季内,天然林、植被混凝土和客土喷播MBN含量均呈现逐步上升的变化动态,且在12月出现峰值,四季变幅较大,分别为20.74~35.26、9.68~13.83、6.91~11.75 mg/kg,且不同样地之间差异显著,天然林MBN含量在四季均显著高于植被混凝土和客土喷播,是植被混凝土、客土喷播的1.57~2.50、1.83~5.00倍。
天然林与客土喷播生境中坡位MBN含量的变化具有显著的季节动态,如图7所示,2种生境MBN含量变化趋势与上坡位大体一致,呈现逐步上升的趋势,在12月出现峰值,MBN含量的变幅为20.74~37.33、8.30~10.37 mg/kg,植被混凝土生境MBN含量变幅为8.30~14.52 mg/kg。天然林生境MBN含量在四季均显著高于植被混凝土和客土喷播,天然林中坡位MBN含量是植被混凝土、客土喷播含量的1.43~3.50、2.5~3.60倍。
客土喷播生境上坡位MBN含量变幅为8.30~14.52 mg/kg。天然林、植被混泥土生境MBN含量的变化具有显著的季节动态,如图8所示,2种生境边坡MBN含量变化趋势与上坡位大体一致,呈逐步上升的趋势,变幅为20.74~37.33、6.22~12.44 mg/kg,在整个生长季内,天然林生境较植被混凝土、客土喷播2种生境上坡位土壤存在显著性差异,天然林生物量氮含量在四季均显著高于植被混凝土、客土喷播,其上坡位MBN是植被混凝土、客土喷播含量的2.17~3.60、2.50~3.40 倍。
将各边坡所对应不同坡位、不同季节的土壤MBN含量求和后计算,得出不同类型边坡土壤MBN含量平均值。如图9所示,天然林、植被混凝土、客土喷播土壤MBN含量平均值分别为28.86、11.75、10.20 mg/kg。天然林的MBN含量平均值最高,分别是混凝土、客土的2.46、2.83倍;客土喷播MBN含量坡位平均值最低,是植被混凝土的0.87倍。
2.3 土壤微生物量含量与土壤环境因子相关性
土壤微生物量碳、氮含量与土壤环境因子相关分析(表2)表明,土壤MBC含量与有机碳含量、全氮含量及含水量呈极显著正相关(P<0.01);土壤MBN含量与有机碳含量、全氮含量及微生物量碳含量呈极显著正相关,与pH值呈极显著负相关。
3 讨论
3.1 不同修复类型土壤微生物量的差异
土壤微生物量作为土壤环境质量的生物指标[3],可以灵敏地反映环境因子变化,对不同土地利用方式和生态功能的变化作出评价[20]。本研究中MBC、MBN含量的范围分别为168.25~694.04、6.22~37.33 mg/kg,天然林MBC、MBN含量大多都显著大于植被混凝土和客土喷播,植被混凝土MBC、MBN含量大于客土喷播,但有时差异不显著。总体来看,植被混凝土肥力水平略优于客土喷播,低于天然林。
土壤微生物量的变化极为复杂,并受土壤理化性质、土壤利用方式及植被类型差异等的影响。对于同一气候条件下不同区域的同种、异种土壤,其土壤内凋落物、根活性及微气象因子也明显不同,这使得这些区域的土壤微生物量也会存在一定差异[21-23]。薛萐等研究发现,人工林生境的MBC含量显著低于自然林生境[24],人工刺槐林虽然可以显著增加土壤MBC含量,但仍显著低于该区顶级群落的土壤MBC[25],本研究结果与之一致。植被混凝土与客土喷播作为人工技术修复边坡,一方面人工修复边坡由于修建时间短,植被演替时间也较短于天然林样地,植被多样性与空间格局都远不及天然林样地。张于光等研究发现,即使土壤性质相似,不同植物群落能够维持的微生物量差異也较大,其中归还土壤的植物残体数量和质量不同,也是造成土壤有机碳、土壤微生物量差异较大的原因之一[26]。另一方面人工技术修复边坡中为了使边坡结构稳定加入了一些物料,这些也可能不利于土壤微生物的生长和繁殖[27],导致其土壤微生物量较低于天然林边坡。而植被混凝土中添加的绿化添加剂等物料能适当提高土壤中微生物量含量,使得植被混凝土微生物量略高于客土喷播。进一步说明植被类型、土地利用方式是影响土壤微生物量的重要因素。
3.2 不同修复类型土壤微生物量碳氮含量的季节变化
土壤微生物量受季节影响明显,这不仅与植物生长节律的自身因素有关,还与气候条件、土壤环境因子及可利用碳和养分资源的赋存环境有关[28]。向家坝工程扰动区3种边坡土壤微生物量碳含量存在明显的季节变化,且3种边坡变化趋势基本一致,表现为MBC含量在秋夏季高于春冬季,峰值出现在9月,这与之前的研究结果[5,29-31]一致。土壤MBC含量与有机碳含量、全氮含量以及含水量呈极显著正相关,说明土壤有机碳含量、全氮含量以及含水量可能是影响本研究区土壤微生物量碳含量季节变化的主要因子,与大多数研究结论[15,32]相同。直接调控森林土壤MBC含量及其季节变化的主导因子不是土地利用方式或地上植被类型,而可能是由于5—9月土温升高、降雨增多使得植物生长旺盛、根系活动能力的增强,根系分泌物含量增加,植物根系的影响又造成土壤有机碳矿化作用增大,进一步形成了利于微生物生长的环境,加上土壤微生物本身的活性随温度而增加,分解利用能源物质的速率加快,维持了较快的周转速率,这些综合因素最终使得土壤微生物量在该段时间出现峰值。
微生物量氮含量也存在明显的季节变化规律,但天然林与2种人工修复边坡变化趋势不同,表现为天然林MBN含量在春季最低,随着植物的生长MBN含量逐渐增加,而植被混凝土边坡与客土喷播边坡则表现为MBN含量在春夏季较低,其他季节中MBN含量相对稳定,这与Franzluebbers等的研究结果[32-33]一致。笔者猜测,雨热同季的环境影响了土壤温度和湿度以及能源物质的输入,为微生物提供了良好的代谢环境,促进了其生长和繁殖。之后,随着土壤温度下降,光照时间减少等因素,不同植被类型下的土壤微生物群落组成受到影响,使不同边坡类型下MBN含量的季节变化趋势的不同。
3.3 不同边坡坡位的土壤微生物量的差异
众多的环境因子中,边坡地形因子影响了光、热、水、土的分布状况,也可导致土壤微生物量发生季节变化[34]。本研究发现,不同季节坡位间MBC、MBN含量存在明显差异,这与张地等运用通径分析模型得到的不同坡位间土壤MBC无显著差异的结果[35]不一致,但与许多研究[36-37]一致。由于不同生境的主要影响因子不同,而土壤微生物量通常受这些因子的交互影响,使得土壤微生物量的季节变化存在差异[38]。白爱芹等在对微地形对土壤微生物特性的影响研究中指出,坡度、坡向影响了土壤养分和水分条件,进而影响了土壤微生物的生物量,使得坡地土壤MBC含量低于平地土壤,使得南坡和西坡土壤MBC含量显著不同[36]。张地等发现,草本丰富度在下坡位对土壤微生物量的负作用突显,而含水量和土壤有机碳含量是影响上坡位和中坡位的主要因素[35]。而秦华军等认为,调节土壤微生物量碳氮含量沿坡位梯度变化的主要生态因子是过氧化氢酶等酶活性,因而土壤MBC含量在下坡与上坡间差异显著[37]。各边坡在不同时期内MBC与MBN含量沿坡位变化没有明显规律,没有出现明显的在坡底富集和坡面流失的趋势,表现出较高的空间异质性,这可能是因为所选取坡地均属于公路两侧的人工修复边坡,其坡度较大且厚度只有10~20 cm,而研究区域多雨和湿润气候,长期的冲刷侵蚀作用使许多矿化产物随着地表和地下径流由高处向坡脚淋洗迁移,组内异质性较为明显,加上在较小尺度下取样,土壤微生物对于环境反应敏感,致使MBC、MBN含量在沿坡位梯度上分布无明显规律。由于本试验没有对凋落物、微生物群落、温度以及速效养分分布进行研究,因此,关于在不同季度下对MBC、MBN含量分布格局沿坡面的响应有待进一步研究。
4 结论
天然林MBC、MBN含量平均值分别是植被混凝土的 1.30、2.46倍,是客土1.70、2.83倍,植被混凝土MBC、MBN含量均高于客土喷播,植被混凝土修复边坡肥力水平高于客土喷播而低于天然林。
3种边坡MBC、MBN含量随季节变化的趋势基本一致,且季节差异明显,表现为MBC含量在秋夏季高于春冬季,峰值出现在9月,MBN含量在秋冬季高于春夏季。表明向家坝工程扰动区土壤微生物生物量碳、氮含量存在明显的季节变化格局差异和空间异质性。
土壤MBC、MBN含量沿坡面空间分布规律性不明显,表现出较高的空间异质性。其中,土壤MBC、MBN含量与有机碳、全氮含量呈极显著正相关,土壤MBC含量与含水量呈极显著正相关。
土壤微生物量碳、氮含量的季节性变化可能受不同机制的驱动,与气候条件、植物生理过程、土壤环境因子与可利用碳和养分资源有关。其中,植被类型与修复模式会引起微生物量氮含量季节性变化差异。
参考文献:
[1]何振立. 土壤微生物量及其在养分循环和环境质量评价中的意义[J]. 土壤,1997,29(2):61-69.
[2]Aison S D,Wallenstein M D,Bradford M A.,Soil-carbon response to warming dependent on microbial physiology[J]. Nature Geoscience,2010,3:336-340.
[3]刘秉儒. 贺兰山东坡典型植物群落土壤微生物量碳、氮沿海拔梯度的变化特征[J]. 生态环境学报,2010,19(4):883-888.
[4]王国兵,阮宏华,唐燕飞,等. 北亚热带次生栎林与火炬松人工林土壤微生物量碳的季節动态[J]. 应用生态学报,2008,19(1):37-42.
[5]Ruan H H,Zou X M,Zimmerman J K,et al. Asynchronous fluctuation of soil microbial biomass and plant litter fall in a tropical wet forest[J]. Plant and Soil,2004,260(1/2):147-154.
[6]Groffman P M,Zak D R,Christensen S,et al. Early spring nitrogen dynamics in a temperate forest landscape[J]. Ecology,1993,74(5):1579-1585.
[7]Burke I C,Lauenroth W K,Riggle R,et al. Spatial variability of soil properties in the shortgrass steppe:the relative importance of topography,grazing,microsite,and plant species in con trolling spatial patternsvz[J]. Ecosystems,1999,2(5):422-438.
[8]周明涛,许文年,夏 栋. 向家坝水电站工程扰动区不同类型边坡土壤酸碱度与肥力分析[J]. 应用生态学报,2010,21(4):1031-1037.
[9]黄真理. 国内外大型水电工程生态环境监测与保护[J]. 长江流域资源与环境,2004(2):101-108.
[10]陈凯麒,芮建良. 中国水电环境影响评价的十年回顾与展望[J]. 环境保护,2012(22):21-24.
[11]曾 旭,陈芳清,郄光武,等. 向家坝工程扰动区3种先锋灌木的光合生理特性[J]. 福建林业科技,2010,37(1):22-26.
[12]李桂媛,陳 静,段中元,等. 水电建设区的生态环境监测与评价水电建设区的生态环境监测与评价——以向家坝水电站为例[J]. 长江流域资源与环境,2013,22(12):1573-1579.
[13]臧逸飞,郝明德,张丽琼,等. 26年长期施肥对土壤微生物量碳、氮及土壤呼吸的影响[J]. 生态学报,2015,35(5):1445-1451.
[14]王春阳,周建斌,夏志敏,等. 黄土高原区不同植物凋落物搭配对土壤微生物量碳、氮的影响[J]. 生态学报,2011,31(8):2139-2147.
[15]魏亚伟,苏以荣,陈香碧,等. 人为干扰对桂西北喀斯特生态系统土壤有机碳、氮、磷和微生物量剖面分布的影响[J]. 水土保持学报,2010,24(3):164-169.
[16]叶建军,周明涛,许文年. 谈喷射护坡绿化技术[J]. 水土保持研究,2004,11(6):194-197.
[17]章梦涛,邱金淡,颜 冬. 客土喷播在边坡生态修复与防护中的应用[J]. 中国水土保持科学,2004,2(3):10-12.
[18]李振高,骆永明,滕 应. 土壤与环境微生物研究法[M]. 北京:科学出版社,2008:28-29.
[19]林启美,吴玉光,刘焕龙. 熏蒸法测定土壤微生物量碳的改进[J]. 生态学杂志,1999,18(2):63-66.
[20]张海燕,肖延华,张旭东,等. 土壤微生物量作为土壤肥力指标的探讨[J]. 土壤通报,200 6,37(3):22 -425.
[21]Kushwaha C P,Tripathi S K,Singh K P. Variations in soil microbial biomass and N availability due to residue and tillage management in a dryland rice agroecosystem[J]. Soil and Tillage Research,2000,56(3/4):153-166.
[22]Wardle D. A comparative assessment of factors which influence microbial biomass carbon and nitrogen levels in soil[J]. Biological Reviews,1992,67(3):321-358.
[23]朱志建,姜培坤,徐秋芳. 不同森林植被下土壤微生物量碳和易氧化态碳的比较[J]. 林业科学研究,2006,19(4):523-526.
[24]薛 萐,刘国彬,戴全厚,等. 侵蚀环境生态恢复过程中人工刺槐林土壤微生物量演变特征[J]. 生态学报,2007,27(3 ):909-917.
[25]王风芹,田丽青,宋安东,等. 华北刺槐林与自然恢复植被土壤微生物量碳氮含量四季动态[J]. 林业科学,2015,19(1):16-24.
[26]张于光,张小全,肖 烨. 米亚罗林区土地利用变化对土壤有机碳和微生物量碳的影响[J]. 应用生态学报,2006,17(11):2029-2033.
[27]吴 彬,夏振尧,赵 娟,等. 植被混凝土基材微生物活性对不同水泥含量的响应[J]. 水土保持通报,2014,34(3):6-9.
[28]王国兵,阮宏华,唐燕飞,等. 森林土壤微生物量动态变化研究进展[J]. 安徽农业大学学报,2009,36(1):100-104.
[29]刘 颖,韩士杰. 长白山四种森林土壤呼吸的影响因素[J]. 生态环境学报,2009,18(3):1061-1065.
[30]胡婵娟,刘国华,陈利顶,等. 黄土丘陵沟壑区坡面上土壤微生物量碳、氮的季节变化[J]. 生态学杂志,2011,30(10):2227-2232.
[31]王风芹,田丽青,宋安东,等. 华北刺槐林与自然恢复植被土壤微生物量碳、氮含量四季动态[J]. 林业科学,2015,51(3):16-24.
[32]Franzluebbers A J,Stuedemann J A. Temporal dynamics of total and particulate organic carbon and nitrogen in cover crop grazed cropping systems[J]. Soil Science Society of America Journal,2014,78(4):1404-1413.
[33]Zak D R,Holmes W E,Tomlinson M J,et al. Microbial cycling of C and N in northern hardwood forests receiving chronic atmospheric NO3-deposition[J]. Ecosystems,2006,9(2):242-253.
[34]李志安,邹 碧,曹裕松,等. 南方典型丘陵退化荒坡地土壤养分特征分析[J]. 生态学报,2003,23(8):1648-1656.
[35]张 地,张育新,曲来叶,等. 坡位对东灵山辽东栎林土壤微生物量的影响[J]. 生态学报,2012,32(20):6412-6421.
[36]白爱芹,傅伯杰,曲来叶,等. 重度火烧迹地微地形对土壤微生物特性的影响——以坡度和坡向为例[J]. 生态学报,2013,33(17):5201-5209.
[37]秦华军,何丙辉,赵旋池,等. 西南喀斯特山区寿竹林地土壤微生物量与酶活性在不同坡位和剖面层次的分布特征[J]. 环境科学,2014,35(9):3580-3586.
[38]王国兵,阮宏华,唐燕飞,等. 北亚热带次生栎林与火炬松人工林土壤微生物量碳的季节动态[J]. 应用生态学报,2008,19(1):37-42.
