稀土尾砂干旱胁迫对2种牧草种子萌发与幼苗生理特性的影响
2019-10-25张海娜鲁向晖王瑞峰
张海娜 鲁向晖 王瑞峰

摘要:稀土矿尾砂保水能力差,干热化严重,常采用冷季草与暖季草混播的方式进行植被恢复。采用不同浓度梯度(0~30%)聚乙二醇(PEG-6000)溶液模拟稀土尾砂干旱,研究常见冷季草多年生黑麦草(Lolium perenne)和高羊茅(Festuca elata)种子萌发和幼苗生长的抗旱性,以期为稀土尾砂冷季草种的选择提供理论依据。研究结果表明,随干旱胁迫加剧,多年生黑麦草和高羊茅种子的发芽率、发芽势、发芽指数呈降低趋势,分别在20%、15%胁迫下显著降低;幼苗期,多年生黑麦草和高羊茅的叶绿素a、叶绿素b、叶绿素a+b的含量呈现先降低后升高的趋势;在中度和重度胁迫下,多年生黑麦草的叶绿素含量、丙二醛(MDA)含量、可溶性蛋白含量、游离氨基酸含量显著高于高羊茅;相关性分析表明,MDA含量、可溶性蛋白含量和游离氨基酸含量之间存在极显著正相关关系;隶属函数评价显示,多年生黑麦草较高羊茅对稀土尾砂干旱胁迫具有更强的适应性。在稀土矿尾砂区植被恢复中可优先选择冷季草多年生黑麦草。
关键词:稀土尾砂;多年生黑麦草;高羊茅;干旱胁迫;渗透调节物质
中图分类号:Q945.78;S540.1 文献标志码: A 文章编号:1002-1302(2019)13-0204-05
离子型稀土矿是我国重要的战略资源,南方红壤区是主要产区。长期以来,稀土矿开采尤其早期多采用堆浸工艺,遗留大面积裸露尾砂地,形成类似“沙漠化”的景观,水土流失严重[1-2]。采取有效的矿山植被修复措施来改善稀土尾砂生态环境是迫切需要解决的问题。然而,稀土尾砂地表裸露、干热化强烈,土壤功能低下[3],尤其季节性干旱期,植物生长受到严重的水分胁迫,甚至威胁到植物幼苗的存活。常见的方法是将冷季草和暖季草混合播种。
多年生黑麦草(Lolium perenne)和高羊茅(Festuca elata)是典型的冷季草种类,分蘖能力强,覆绿速度快,是我国广大温带地区地表覆绿的重要材料,关于这2种植物抗旱性的研究较多。在研究PEG模拟干旱胁迫对冷季型草坪草种子萌发和幼苗生长的影响中发现,高羊茅的抗旱性高于多年生黑麦草[4-5]。也有研究采用盆栽育苗法对冷季型草种生长期的生理特性进行比较,表明高羊茅抗旱性高于多年生黑麦草[6-8];高涵等对几种冷季草苗期的抗旱性研究显示,多年生黑麦草高于高羊茅[9]。可见,多年生黑麦草和高羊茅抗旱性的研究多集中在草坪成坪中植物生长旺盛期或后期,而且不同时期2种草本植物的抗旱性不同。除了作为草坪草,这2种植物还被用于废弃矿区和污染土壤的生态修复。张诗吟等研究了新疆煤矿废弃地7种牧草的叶绿素荧光特征与抗旱性的关系,发现高羊茅较多年生黑麦草具有更强的抗旱性[10]。孙伟等在对湖南省铅锌尾矿污染区适生植物进行筛选时发现,黑麦草对铅和锌有较强的迁移能力和富集能力[11]。谷超等研究了红枫湖底泥中重金属污染的植物修复特征,发现高羊茅对锌的吸收能力较强,黑麦草对铜和铅的吸收能力较强[12]。
种子萌发作为植物生活史的关键阶段(包括幼苗),决定着植物对环境的适应性和生存能力[13]。目前多年生黑麦草和高羊茅作为水土保持草种在江西稀土矿植被恢复区有所种植,但是它们对稀土尾砂生境适应性的研究尚且缺乏。本研究采用不同浓度(0~30%)聚乙二醇(PEG-6000)模拟稀土尾砂干旱对多年生黑麦草和高羊茅种子萌发与幼苗生长的影响,比较2种植物的抗旱性,旨在为稀土尾砂矿区混播草种的选择提供依据。
1 材料与方法
1.1 试验材料
供试草种均采自寻乌废弃稀土矿区周边,采集时间为试验前1年,室内常温条件保存。
供试稀土尾砂基质取自江西省寻乌县某废弃稀土矿尾砂区,采集表层0~20 cm范围内的尾砂,对其理化性状指标进行测定。根据国际制土壤质地分类,该尾砂土壤粒径在 2.00~0.02 mm范围的含量约占65%,属于沙质壤土,田间持水量15.02%,容重2.25 g/cm3;pH值为5.08,属于强酸性,电导率高达79.12 dS/m,是原红壤(6.9 dS/m)的10倍多,尾砂中土壤全磷含量、全氮含量、有机碳含量、有机质含量分别为1.21 g/kg、0.20 mg/kg、6.29、10.85 g/kg,速效磷含量、速效钾的含量分别为13.78、225.97 mg/kg。由此可见,尾砂土壤保水能力差,养分含量低。
1.2 试验设计与测定方法
1.2.1 试验设计及测定 2017年4月中旬在南昌工程学院水土保持实验室的培养箱中进行种子萌发试验。选择饱满、种皮完好的种子,用70%乙醇消毒5 min,蒸馏水冲洗4~5次。設定PEG-6000浓度为5%、10%、15%、20%、25%、30%共6个处理,同时以蒸馏水(CK1)、稀土尾砂浸提液(CK2)(浸提比例为1 ∶ 1,过滤后试验备用)为对照,其中5%、10%为轻度胁迫,15%、20%为中度胁迫,25%、30%为重度胁迫。将准备好的种子置于垫有2层滤纸的培养皿中,分别加入配置好的处理液,每个培养皿放50粒种子,重复3次,置于人工气候箱中,设定温度为白天25 ℃,夜晚20 ℃,每天光照时间为08:00—18:00,共10 h,光照度为2 000 lx。每天定时统计种子发芽数(以种子露白为准),2 d计算发芽势,9 d计算发芽率、发芽指数[7]。每2 d更换1次滤纸。
发芽率=(9 d内种子发芽数/供试种子数)×100%;
发芽势=(2 d内种子发芽数/供试种子数)×100%;
发芽指数=∑(Gt/Dt)。
式中:Gt为t d的发芽数;Dt为发芽时间,d。
1.2.2 幼苗渗透调节物质测定 种子萌发试验的同时,在南昌工程学院生物技术基地进行幼苗生长试验。将消毒过的种子播种于装有稀土尾砂的育苗盆(直径15 cm×高10 cm)内,待种子在自然条件下生长,试验时期为南昌雨季,无须添加水分。试验50 d后,每盆留生长一致的幼苗15株,3个重复。将育苗盆移入遮雨棚下,每天09:00用浓度为10%、20%、30%的PEG-6000溶液对多年生黑麦草和高羊茅幼苗进行轻度、中度、重度干旱处理,处理液体积为20 mL,蒸馏水处理为对照CK。处理7 d后采集幼苗地上部分,迅速用冰盒带回实验室低温-80 ℃保存,用于测定渗透调节物质含量。其中叶绿素含量的测定采用分光光度计法;丙二醛含量采用硫代巴比妥酸法测定;可溶性蛋白质含量采用考马斯亮蓝G-250染色法测定;游离脯氨酸含量采用磺基水杨酸法提取测定;植物组织中游离氨基酸含量采用茚三酮法测定[14]。
1.3 数据处理
采用SPSS 17.0对数据进行单因素方差分析和多重比较;用隶属函数值法进行植物抗旱性综合评价。
耐旱性综合评价采用隶属函数值法[15]。计算公式如下:
(1)
式中:u(Xij)为i物种j指标的隶属函数值;Xij为i物种j指标的测定值;Xjmax和Xjmin分别为j指标的最大值和最小值。
如果某一指标与耐旱性呈负相关,则用反隶属函数计算:
(2)
运用公式(1)或公式(2)求出不同植物多项抗旱指标的平均隶属函数值,平均隶属函数值越大,表示该种植物抗旱性越强。
2 结果与分析
2.1 模拟干旱对2种植物种子萌发的影响
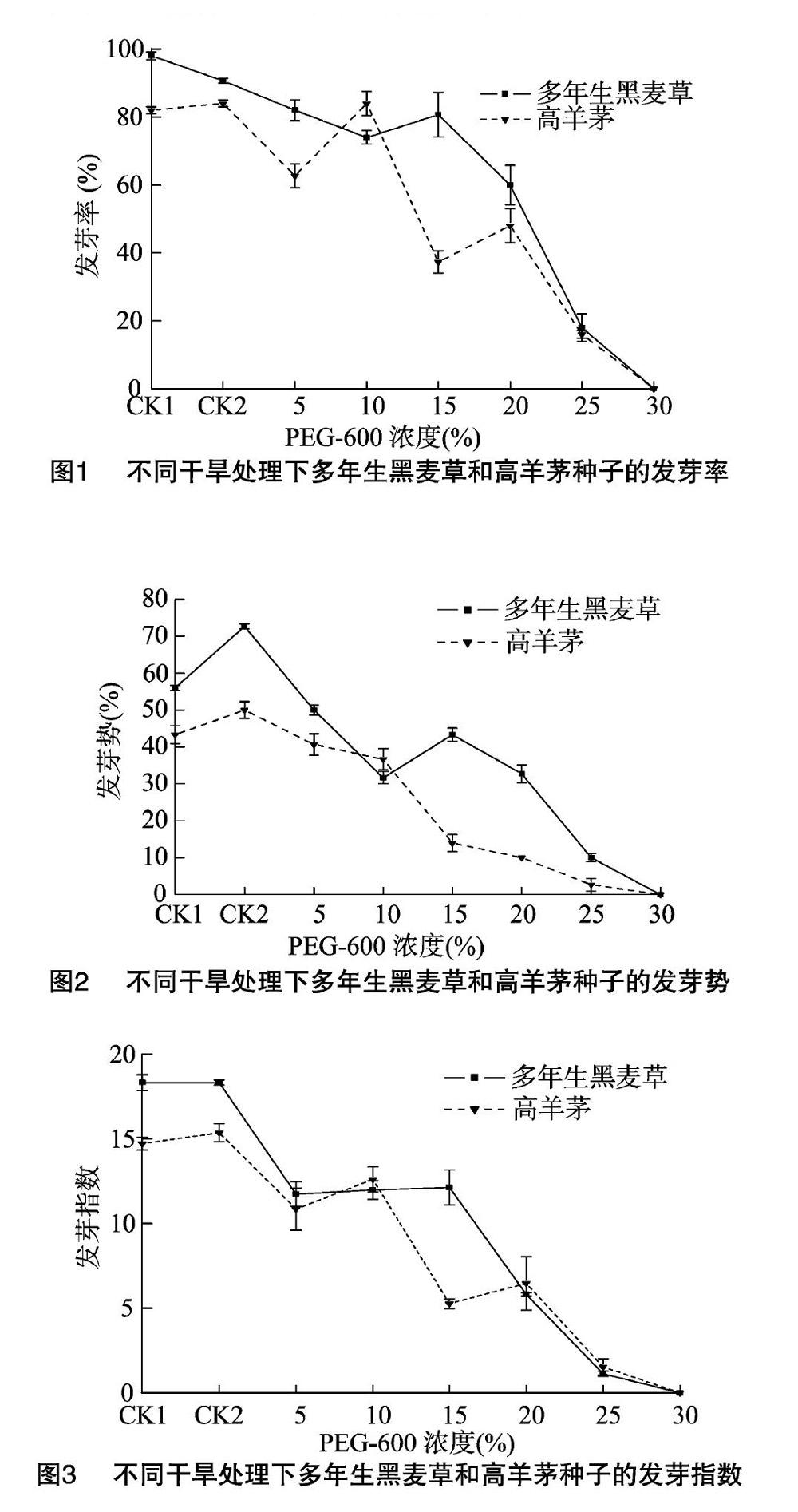
2.1.1 发芽率 不同处理下,随着干旱程度的加强,多年生黑麦草和高羊茅的发芽率呈下降趋势;PEG-6000浓度为5%~30%时,其发芽率低于蒸馏水(CK1)、浸提液(CK2);与蒸馏水CK1相比,浸提液CK2中多年生黑麦草的发芽率明显降低,两者处理的高羊茅则不差异明显;多年生黑麦草和高羊茅的发芽率分别在20%和15%胁迫时明显降低,说明中度胁迫大幅抑制了2种植物的种子萌发;25%浓度胁迫时,二者发芽率为17%左右,30%时种子停止发芽(图1)。
2.1.2 发芽势和发芽指数 多年生黑麦草和高羊茅的发芽势、发芽指数均随着干旱程度增强呈降低趋势。多年生黑麦草的发芽势在CK2条件下明显高于CK1,发芽指数在这2个条件下无明显差异;高羊茅的发芽势、发芽指数在CK1和CK2之间均无明显差异,且多年生黑麦草的发芽势、发芽指数明显高于高羊茅。在PEG-6000的浓度为5%~30%条件下,2种植物的发芽势和发芽指数均明显低于CK1、CK2;相较于CK1,高羊茅的发芽势、发芽指数在15%的胁迫下分别明显降低了 67.69%、64.20%,多年生黑麦草则在20%胁迫下明显降低了41.61%、68.30%。另外,轻度(5%、10%)胁迫和重度(25%、30%)胁迫条件下的发芽势和发芽指数在2种植物之间无明显差异(图2、图3)。
2.2 模擬干旱对2种植物幼苗生理特性的影响
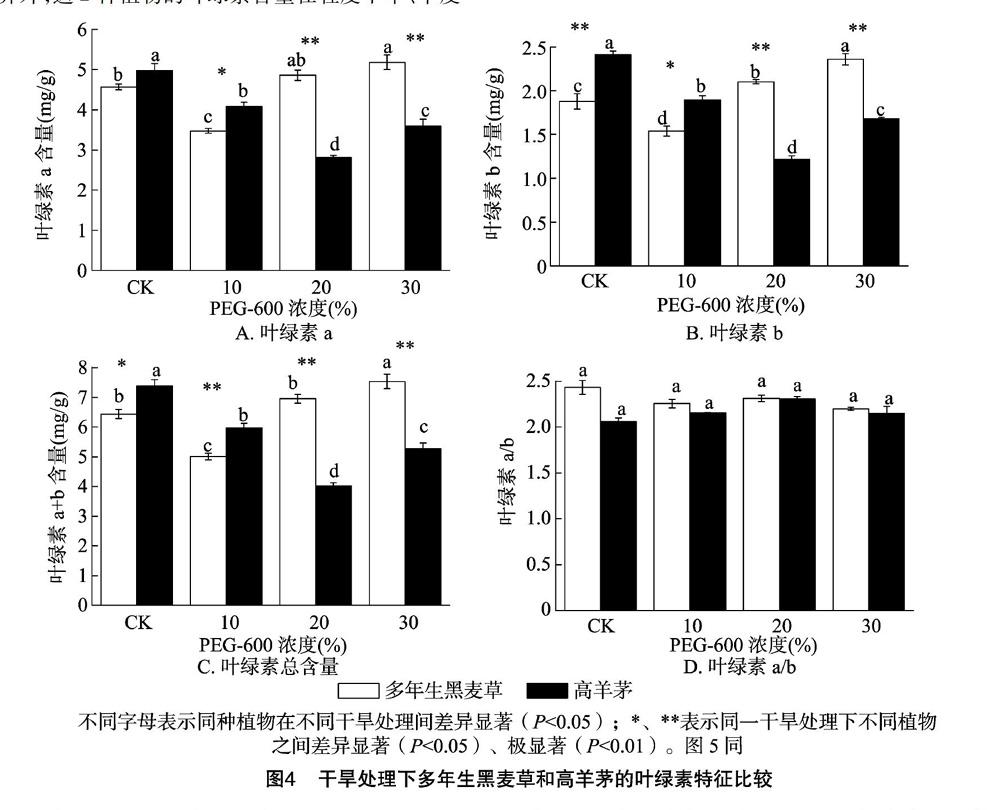
2.2.1 叶绿素含量 多年生黑麦草和高羊茅的叶绿素a、叶绿素b和叶绿素a+b含量变化趋势随干旱胁迫(10%~30% PEG-6000浓度)的增强基本一致,多年生黑麦草的叶绿素a、叶绿素b、叶绿素a+b含量随着胁迫程度增强而升高,高羊茅则呈先下降后升高的趋势(图4-A、图4-B、图4-C);除了多年生黑麦草的叶绿素a含量在中度干旱和重度干旱之间无显著差异外,这2种植物的叶绿素含量在轻度干旱、中度干旱和重度干旱之间差异显著。对相同处理来说,对照和轻度胁迫下高羊茅的叶绿素含量高于多年生黑麦草,而中度和重度胁迫下的多年生黑麦草的叶绿素含量极显著高于高羊茅。对叶绿素a/b来说,不管是不同处理间还是不同植物之间均无显著差异(图4-D)。
2.2.2 渗透调节物质 随干旱胁迫程度(10%~30%PEG-6000)的增强,多年生黑麦草幼苗的丙二醛含量逐渐降低,重度干旱时最低,比对照降低了28.31百分点(图5-A),可溶性蛋白质和游离氨基酸含量呈相似的变化趋势,在轻度胁迫、重度胁迫条件下较高,与对照和中度胁迫时差异显著(图5-B、图5-C);中度干旱、重度干旱脯氨酸含量显著高于对照和轻度干旱(图5-D),说明不同程度的干旱胁迫可诱使多年生黑麦草通过不同的渗透物质来维持生长。随干旱胁迫的增强高羊茅的丙二醛含量呈现增高趋势、可溶性蛋白含量、游离氨基酸含量先降低后升高,重度胁迫下游离等基酸含量显著高于低中度胁迫(图5-A、图5-B、图5-C)。对这2种植物来说,不管是在哪种胁迫程度下,多年生黑麦草的丙二醛含量、可溶性蛋白含量、游离氨基酸的含量均显著高于高羊茅;而且干旱胁迫下多年生黑麦草各渗透调节物质的变化幅度高于高羊茅,说明多年生黑麦草在应对干旱胁迫时具有更大潜力(图5)。
2.3 多年生黑麦草和高羊茅各指标的相关性
多年生黑麦草和高羊茅幼苗生长中各指标相关性分析(表1)显示,叶绿素a含量、叶绿素b含量和叶绿素a+b之间呈均极显著正相关,这3个指标与叶绿素a/b之间呈负相关,但相关性不显著。渗透调节物质中,丙二醛含量、可溶性蛋白质含量与游离氨基酸含量两两之间存在极显著的正相关关系,相关系数分别为0.657、0.586、0.817。
2.4 多年生黑麦草和高羊茅对稀土尾砂干旱适应性的综合评价
通过11个指标对干旱处理下多年生黑麦草、高羊茅种子萌发和幼苗生长的适应性进行综合评价,从表2可以看出,多年生黑麦草、高羊茅各指标的平均隶属函数值为0.639、0.329,说明多年生黑麦草对稀土尾砂干旱胁迫的适应性高于高羊茅。
3 讨论与结论
稀土尾砂废弃地土壤结构严重沙化,保水能力差,植物易遭受水分胁迫。种子为植物最重要的繁殖器官,其种子萌发和幼苗阶段的生长对植物的成活起着决定性的作用,较其他阶段更容易受到水分等环境因子的影响。采用PEG-6000模拟水分胁迫,是通过抑制种子吸水过程而抑制其萌发,随着干旱胁迫的增强,多数植物种子的发芽率、发芽势、发芽指数均呈下降趋势[16-18],也有低浓度PEG-6000处理促进植物种子萌发的报道[19]。在本试验中,多年生黑麦草和高羊茅的发芽情况随胁迫程度增强呈下降趋势,说明干旱抑制了这2种植物的种子萌发,轻度胁迫也未起到促进作用,这与史薇等的研究结果[16,20]一致。当PEG-6000浓度分别为20%、15%时显著降低,说明多年生黑麦草较高羊茅在中度胁迫下具有较强的发芽能力;当浓度达到25%时,2种植物的发芽率、发芽势、发芽指数极低,30%浓度下,种子失去萌发能力。
叶绿素含量是植物对干旱胁迫反应敏感性的生理指标,大量的研究结果表明,干旱胁迫通过抑制叶绿素合成并加速其分解使叶绿素含量直线下降[21]。本研究显示,随着干旱胁迫加剧,多年生黑麦草和高羊茅的叶绿素含量先降低后升高,分别在轻度(10%)和中度(20%)干旱胁迫下最低,且中度和重度干旱胁迫下,多年生黑麦草的叶绿素含量显著高于高羊茅,说明在中度、重度干旱条件下,多年生黑麦草受到影响较小,具有更强的光合潜力。
渗透调节作用是植物适应水分胁迫的重要机制,其中丙二醛是膜脂过氧化的产物[22],通过丙二醛的变化可了解干旱胁迫对植物的破坏程度;游离氨基酸、脯氨酸和可溶性蛋白是植物体内的主要渗透调节物质,三者均能通过降低细胞原生质的渗透势,促使细胞从外界吸收水分,使植物保持一定的含水量和膨压[23],减轻胁迫对植物体的伤害[24-25]。本试验中,不管是在轻度、中度干旱胁迫下还是在重度干旱胁迫下,多年生黑麦草的丙二醛、可溶性蛋白质和游离氨基酸的含量均显著高于高羊茅,且变化幅度较大,说明多年生黑麦草受到干旱胁迫时,其体内的渗透调节物质大量积累,减少干旱对细胞膜造成破坏,使植物具有更强的渗透调节能力。相关性分析也表明,丙二醛含量与游离氨基酸和可溶性蛋白的含量极显著相关(r=0.586、r=0.657),这与王晓龙等的研究结果[26]一致。
也有研究指出,大多渗透调节物质均随干旱胁迫的增加呈递增或者先升高后降低的趋势[7,16],但是不同植物在不同环境条件下各不相同。本研究中多年生黑麦草和高羊茅叶片中脯氨酸含量随胁迫加剧而上升,这与以往研究结果[21,23]一致;而可溶性蛋白质和游离氨基酸含量则呈现先上升后降低然后又上升的趋势,一方面与稀土尾砂复杂的土壤环境胁迫有关,另一方面轻度胁迫(10%)下植物的渗透调节能力占主导地位,叶片中可溶性蛋白质和游离氨基酸含量很高,中度胁迫(20%)时可能激发抗氧化酶系统活性,大量氨基酸和可溶性蛋白合成保护酶类物质含量显著降低[21,24],而在重度干旱胁迫下,酶活性受到抑制,其合成减弱,进而使游离氨基酸和可溶性蛋白质含量大幅升高[17]。不同渗透调节物质的相关性分析表明,游离氨基酸含量和可溶性蛋白质含量呈极显著正相关,与脯氨酸呈负相关,主要是由于游离氨基酸是合成植物生长所需蛋白质和酶类的基本物质[27],而游离氨基酸的种类有19种,不同氨基酸的调节能力和响应时间存在差异。
在不同的PEG浓度下,各种植物的抗旱性程度不会保持不变,植物种子萌发和幼苗阶段的抗旱性信息不同,应该对生长阶段的抗旱性分別评价[28],但是在实际生产中,种子萌发特性很好地植物可能在幼苗期由于高温或者缺水而死亡,只有在种子萌发和幼苗阶段都有较高抗旱性的植物才能很好地存活。因此,本研究中将种子萌发和幼苗阶段作为整体进行隶属函数综合评价,结果表明,多年生黑麦草具有更强的抗旱性。
稀土尾砂干旱环境下,多年生黑麦草较高羊茅具有更强发芽率、发芽势、发芽指数,幼苗生长阶段,植物通过叶绿素、丙二醛、可溶性蛋白、游离氨基酸等生理调节作用来维持生长,多年生黑麦草对稀土尾砂干旱环境有更好的适应性。在稀土矿尾砂区采用植物措施修复时,可优先考虑选择冷季草多年生黑麦草,与暖季草混合进行快速覆绿。
参考文献:
[1]Li X F,Chen Z B,Chen Z Q,et al. A human health risk assessment of rare earth elements in soil and vegetables from a mining area in Fujian Province,Southeast China[J]. Chemosphere,2013,93(6):1240-1246.
[2]Liang T,Li K X,Wang L Q. State of rare earth elements in different environmental components in mining areas of China[J]. Environmental Monitoring and Assessment,2014,186(3):1499-1513.
[3]刘文深,刘 畅,王志威,等. 离子型稀土矿尾砂地植被恢复障碍因子研究[J]. 土壤学报,2015,52(4):879-887.
[4]荣秀连,王 波,刘 刊,等. PEG-6000模拟干旱胁迫对冷季型草坪种子萌发特性影响[J]. 北方园艺,2010(8):80-82.
[5]郑轶琦,臧国长. PEG胁迫对冷季型草坪草种子萌发和幼苗生长的影响[J]. 种子,2011,30(12):34-37.
[6]初晓辉,岳信龙,任 健,等. 水分胁迫下4种冷季型草坪草抗旱性比较研究[J]. 草原与草坪,2012,32(4):15-19.
[7]侯 敏. 干旱胁迫对3种草坪草的生长及叶绿素含量的影响[J]. 山西林业科技,2015,44(3):24-25.
[8]赵志军,单贵莲,段新慧,等. 干旱胁迫对3种冷季型草坪草光谱反射率及生理特征的影响[J]. 草原与草坪,2016,36(6):23-29.
[9]高 涵,吴 伟,刘秀萍,等. 水分胁迫下几种冷季型草坪草抗旱机理研究[J]. 水土保持研究,2006,13(3):126-128.
[10]张诗吟,杨建军,莫 爱. 干旱胁迫下七种牧草叶绿素荧光参数与抗旱性关系研究[J]. 湖北农业科学,2017,56(1):107-111.
[11]孙 伟,王 琼,张 望,等. 湖南省铅锌尾矿污染区植被修复适生植物选择研究[J]. 价值工程,2016,35(17):208-211.
[12]谷 超,梁隆超,陈 卓. 4种牧草植物对红枫湖底泥中重金属污染的植物修复研究[J]. 环境工程,2015,33(7):148-151.
[13]张佳宁,刘 坤. 植物调节萌发时间和萌发地点的机制[J]. 草业学报,2014,23(1):328-338.
[14]李合生. 植物生理生化实验原理和技术[M]. 北京:高等教育出版社,2000.
[15]种培芳,苏世平,李 毅. 4个地理种群红砂的抗旱性综合评价[J]. 草业学报,2011,20(5):26-33.
[16]史 薇,徐海量,趙新风,等. 胀果甘草种子萌发对干旱胁迫的生理响应[J]. 生态学报,2010,30(8):2112-2117.
[17]Abudureheman B,Yan C,Wei Y. Fruit growth and seed germination characteristics of Nanophyton erinaceum:a dominant desert shrub[J]. International Journal of Plant Research,2012,25(1):34-40.
[18]Zeng Y J,Wang Y R,Baskin C C,et al. Testing seed germination responses to water and salinity stresses to gain insight on suitable microhabitats for restoration of cold desert shrubs[J]. Journal of Arid Environments,2014,100(1):89-92.
[19]闫兴富,周立彪,思彬彬,等. 不同温度下PEG-6000模拟干旱对柠条锦鸡儿种子萌发的胁迫效应[J]. 生态学报,2016,36(7):1989-1996.
[20]胡 红,曹 昀,王 颖. 水分胁迫对狗牙根种子萌发及幼苗生长的影响[J]. 草业科学,2013,30(1):63-68.
[21]桑子阳,马履一,陈发菊. 干旱胁迫对红花玉兰幼苗生长和生理特性的影响[J]. 西北植物学报,2011,31(1):109-115.
[22]董伊晨,刘悦秋. 土壤水分对异株荨麻(Urtica dioica)保护酶和渗透调节物质的影响及其与叶片光合和生物量的相关性[J]. 生态学报,2009,29(6):2845-2851.
[23]李 力,刘玉民,王 敏,等. 3种北美红枫对持续高温干旱胁迫的生理响应机制[J]. 生态学报,2014,34(22):6471-6480.
[24]谢志玉,张文辉,刘新成. 干旱胁迫对文冠果幼苗生长和生理生化特征的影响[J]. 西北植物学报,2010,30(5):948-954.
[25]丁玉梅,马龙海,周晓罡,等. 干旱胁迫下马铃薯叶片脯氨酸、丙二醛含量变化及与耐旱性的相关性分析[J]. 西南农业学报,2013,26(1):106-110.
[26]王晓龙,李 红,杨 曌,等. 干旱胁迫下4种禾本科牧草根系抗旱性比较[J]. 中国饲料,2016(13):22-24,32.
[27]张海娜,方向文,蒋志荣,等. 柠条平茬处理后不同组织游离氨基酸含量[J]. 生态学报,2011,31(9):2454-2460.
[28]罗 冬,王明玖,李元恒,等. 四种豆科牧草种子萌发和幼苗生长对干旱的响应及抗旱性评价[J]. 生态环境学报,2015,24(2):224-230.
