罗格列酮保护氧糖剥夺复氧PC12细胞通过减少HMGB1释放和上调DUSP8
2019-07-09张美杰李文静毛森林刘美玲蔡灵钰俞春江
王 莉,张美杰,李文静,毛森林,刘美玲,黄 山,蔡灵钰,俞春江
研究证实PPARγ受体激动剂在脑缺血再灌注损伤、帕金森病和肌萎缩侧索硬化等模型中减少神经元凋亡[1~5],但PPARγ在缺血性脑损伤的具体机制尚不明确。炎性细胞分泌HMGB1,在脑缺血再灌注损伤中具有启动与放大免疫反应的作用,PPARγ受体激动剂可减少HMGB1释放[6]。近年报道PPARγ抗凋亡可能与双特异性蛋白磷酸酶有关[7]。研究表明MAPK信号通路参与脑缺血再灌注后细胞凋亡,双特异性蛋白磷酸酶使MAPK通路重要成员去磷酸化,发挥抗凋亡作用[7]。因此,我们推测PPARγ受体激动剂可能减少HMGB1分泌,并上调双特异性蛋白磷酸酶8,发挥神经保护作用。本研究采用氧葡萄糖剥夺/复氧PC12细胞模型,观察罗格列酮干预后的PPARγ、RAGE、HMGB1、DUSP8、Bcl-xl等指标变化,探讨罗格列酮对缺血再灌注损伤神经细胞的保护作用及机制。
1 材料和方法
1.1 细胞培养及分组 PC12细胞接种在含10%胎牛血清的1640培养基中,置于5% CO237 ℃温箱培养。培养液每2 d更换一次,待细胞覆盖培养瓶壁70%~80%,按照1∶4的比例传代。分组:(1)正常对照组:正常培养的PC12细胞;(2)溶剂对照组(vehicle 组): 含0.1%DMSO的培养液处理的PC12细胞;(3)OGD10 h/R24 h组:PC12细胞氧葡萄糖剥夺10 h/复氧24 h;(4)OGD10 h/R24 h+RGZ组:OGD10 h后复氧时给予RGZ作用于PC12细胞24 h;(5)OGD10 h/R24 h+RGZ+GW9662组:OGD10 h后复氧时给予RGZ和GW9662作用于PC12细胞24 h。每组设6个复孔。
1.2 主要试剂和仪器 大鼠HMGB1 ELISA试剂盒(上海西唐生物科技有限公司);多克隆兔抗鼠PPARγ 抗体(Santa Cruz公司),多克隆兔源抗DUSP8抗体(ORIGENE公司,美国);辣根酶标记山羊抗兔二抗(北京中杉金桥生物技术有限公司);PPARγ 激动剂罗格列酮(rosiglitazone,RGZ);PPARγ 拮抗剂 GW9662 均购自 Cayman Chemical 公司;全自动酶标仪(BIO-RAD 680,美国);倒置相差显微镜(Nikon TS100,日本)。
1.3 OGD/R模型建立 将PC12细胞按照2×104/ml密度接种于96孔板,每孔加入培养液约100 μl,置5% CO2培养箱24 h。待细胞贴壁后,将对照组培养基更换为含糖Earle’s平衡盐溶液继续培养;用无糖Earle’s平衡盐溶液替换实验组的完全培养基,将实验组放入37 ℃,含95% N2、5% CO2混合气体的培养箱10 h后,将对照组和实验组更换为完全培养基,再置于37 ℃、5% CO2培养箱中分别培养6 h、12 h、24 h以及48 h模拟不同的再灌注时间。
1.4 细胞存活率检测 将PC12细胞接种在96孔板中,并通过四基甲偶氮唑蓝(MTT)测定细胞活力。PC12细胞OGD10 h后分别复氧6 h、12 h、24 h、48 h后,在常规培养基中于37 ℃与MTT(100 μg/ml)一起温育。4 h后洗涤细胞,然后加入100 μl二甲亚砜(DMSO)溶解。使用酶联免疫吸附测定仪,在570 nm监测产物形成。
1.5 Western blot检测PPARγ、RAGE和DUSP 8及 Bcl-xl蛋白表达水平 吸弃培养液,预冷PBS洗涤两次,将细胞移入1 ml离心管,4 ℃下2000 rpm离心5 mim,弃上清,在离心沉淀的细胞碎片内加入150 μl细胞裂解液及1.5 μl蛋白酶抑制剂(PMSF)混匀后于冰上裂解30 min,4 ℃下12000 rpm离心30 min,取上清液移至EP管-80 ℃保存。以10%SDS-聚丙烯酰胺凝胶电泳分离,半干电转移法转移至PVDF膜,室温下封闭1 h,分别加入1∶500稀释的单克隆兔抗鼠PPARγ一抗(美国Santa Cruz,sc-73554);1∶500稀释的多克隆兔抗鼠DUSP8一抗(ORIGENE公司,美国,TA322744),4 ℃孵育过夜;分别加入1∶3000稀释的辣根酶标记山羊抗兔二抗(北京中杉金桥生物技术有限公司,ZB2301),室温孵育2 h。化学发光法显色,应用凝胶图像处理系统分析目标条带的分子量和净光密度值。
1.6 ELISA法检测HMGB1水平 取各组细胞上清液 40 μl,按照说明书步骤操作,在波长为 450 nm下测量标准孔、样本孔的OD值,根据标准曲线计算HMGB1含量。
2 结 果
2.1 建立OGD/R细胞损伤模型 PC12细胞经缺氧低糖处理10 h后,经不同复氧时间6~48 h后,测定细胞存活率。PC12细胞缺氧低糖处理10 h后,其存活率随再灌注时间延长而降低,细胞存活率依次为(82.63±2)%、(66.17±3.82)%、(25.04±3.48)%、(24.44±0.17)%,同对照组相比差异明显(P<0.01)。OGD10 h/R48 h组同OGD10 h/R24 h组相比较,无显著差异(P>0.05),故选择OGD10 h/R24 h作为适宜的细胞损伤模型(见图1A)。
2.2 罗格列酮和GW9662在OGD/R细胞模型中对细胞存活率影响
2.2.1 不同浓度罗格列酮对OGD/R处理后细胞存活率影响 PC12细胞OGD10 h后,复氧时给予不同浓度罗格列酮(0~15 μmol)作用24 h,检测细胞存活率。结果显示vehicle组与罗格列酮5 μmol组细胞生存率分别为(99.07±0.4)%、(101.07±4.55)%,同RGZ 0 μmol组相比无显著性差异(P>0.05);罗格列酮10 μmol和15 μmol组,细胞存活率分别为(148.56±6.1)%、(135.35±2.46)%,两组相比无显著差异(P>0.05);罗格列酮10 μmol和15 μmol组,同RGZ 0 μmol组相比,细胞存活率明显上升,差异均显著(P<0.01)(见图1B)。
2.2.2 不同浓度GW9662对罗格列酮作用的OGD/R处理后PC12细胞存活率的影响 PC12细胞OGD10 h后,复氧时给予10 μmol罗格列酮与不同浓度GW9662(0~15 μmol)作用24 h,测定细胞存活率。结果显示GW9662浓度为10 μmol、15 μmol时,细胞生存率下降至(42.36±1.43)% 和(31.08±1.08)%,同RGZ10 μmol/GW9662 0 μmol组相比较,差异明显(P<0.01)。RGZ 10 μmol/GW9662 15 μmol组与RGZ 10 μmol/GW9662 10 μmol组比较,无显著差异(P>0.05)(见图1C)。
2.3 罗格列酮在OGD/R细胞模型中对PPARγ受体的影响 PC12细胞OGD10 h后,复氧时给予10 μmol罗格列酮与15 μmol GW9662作用24 h,观察其对PPARγ蛋白的影响。结果显示,同对照组相比,PC12细胞经过OGD10 h/R24 h处理后,PPARγ蛋白表达水平明显下降(P<0.01);同OGD10 h/R24 h组相比,罗格列酮治疗组PPARγ蛋白表达水平上升(P<0.01);而GW9662能够下调罗格列酮引起的PPARγ蛋白表达上升,且同罗格列酮治疗组相比较,GW9662组蛋白表达明显下降(P< 0.01)(见图2A)。
2.4 罗格列酮在OGD/R细胞模型中抗晚期炎症反应的作用
2.4.1 罗格列酮在OGD/R细胞模型中对HMGB1影响 PC12细胞OGD10 h后,复氧时给予10 μmol罗格列酮与15 μmol GW9662作用24 h后,测定细胞HMGB1分泌量。结果显示,同对照组相比,PC12细胞经过OGD10 h/R24 h处理后,HMGB1分泌量明显上升(P<0.01);罗格列酮组HMGB1分泌量下降55.9 %,差异显著(P<0.01),提示罗格列酮可以通过减少HMGB1而发挥抗炎效应。而加入GW9662后,HMGB1分泌量上升达59.8%,差异显著(P<0.01)(见图2B)。罗格列酮减少HMGB1释放的效应,被GW9662所阻断,提示其作用机制经由PPARγ受体介导。
2.4.2 罗格列酮在OGD/R细胞模型中对RAGE蛋白影响 PC12细胞OGD10 h后,复氧时给予10 μmol罗格列酮与15 μmol GW9662作用24 h后,检测RAGE蛋白表达水平。结果显示,OGD10 h/R24 h后,RAGE表达显著增加,而经过罗格列酮处理后,RAGE表达明显下调,但罗格列酮这种作用可被GW9662所阻断,表现为GW9662处理组RAGE蛋白表达显著增加 (P<0.01)(见图2C),提示其作用机制经由PPARγ受体介导。
2.5 罗格列酮在OGD/R细胞模型中抗凋亡作用
2.5.1 罗格列酮在OGD/R细胞模型中对DUSP8蛋白影响 PC12细胞OGD10 h后,复氧时给予10 μmol罗格列酮与15 μmol GW9662作用24 h后,检测DUSP8表达情况。结果显示,同对照组相比,OGD10 h/R24 h处理后,DUSP8表达水平明显下降(P<0.01);罗格列酮干预后DUSP8表达水平明显上升(P<0.01);而GW9662能够下调DUSP8表达上升,表现为GW9662处理组DUSP蛋白表达显著降低 (P<0.01)(见图3A),提示罗格列酮引起DUSP8蛋白上调是经由PPARγ受体介导的。
2.5.2 罗格列酮在OGD/R细胞模型中对Bcl-xl蛋白影响 PC12细胞OGD10 h后,复氧时给予10 μmol罗格列酮与15 μmol GW9662作用24 h后,检测细胞Bcl-xl表达情况。结果显示,同对照组比较,OGD10 h/R24 h处理后,Bcl-xl表达水平明显下降 (P<0.01);罗格列酮组Bcl-xl表达水平显著上升(P<0.01);而GW9662能够下调罗格列酮引起的Bcl-xl表达上升,因此GW9662组的Bcl-xl表达明显下降(P<0.01)(见图3B)。说明罗格列酮引起的抗凋亡蛋白Bcl-xl上调是经由PPARγ受体介导的。

A:不同再灌注时间对PC12细胞存活率影响;B:不同浓度罗格列酮和0.1%DMSO对OGD10 h/R24 h的PC12细胞存活率影响;C:不同浓度GW9662对10 μmol罗格列酮作用后OGD10 h/R24 h的PC12细胞存活率影响。与control组相比**P<0.01;与RGZ 0 μmol组相比**P<0.01;与RGZ10 μmol/GW9662 0 μmol组相比◆◆P<0.01
图1 不同再灌注时间以及不同浓度罗格列酮和GW9662对PC12细胞存活率影响

A:PPARγ蛋白表达;B:HMGB1蛋白分泌情况;C:RAGE蛋白表达。与对照组相比较##P<0.01;与OGD10 h/R24 h组相比较**P<0.01;与OGD10 h/R24 h+10 μmol RGZ组相比较■■P<0.01
图2 罗格列酮10 μmol、GW9662 15 μmol对OGD10 h/R24 h的PC12细胞的PPARγ、HMGB1以及RAGE蛋白表达的影响
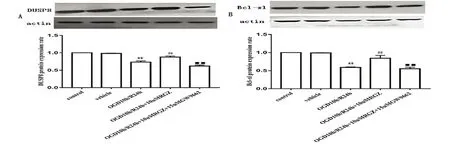
A:对DUSP蛋白表达影响;B:对Bcl-xl蛋白表达影响。与对照组相比较**P<0.01;与OGD10 h/R24 h组相比较##P<0.01;与OGD10 h/R24 h+10 μmol RGZ组相比较■■P<0.01
图3 罗格列酮10 μmol与GW9662 15 μmol对OGD10 h/R24 h处理的PC12细胞的DUSP8和Bcl-xl蛋白表达影响
4 讨 论
近年来对缺血性脑卒中研究热点集中于缺血再灌注损伤的具体机制,已证实PPARγ激动剂能够减轻缺血再灌注损伤、帕金森病及肌萎缩侧索硬化等细胞模型的损伤,这为PPARγ激动剂用于治疗脑组织缺血再灌注损伤提供了充分研究基础[8,9]。
有研究报道罗格列酮可以减轻缺血后的炎症反应,涉及IL-1、IL-6、TNF等炎性介质[10],本研究进一步证实了罗格列酮可以通过减少HMGB1的释放,而发挥减轻脑缺血再灌注后晚期炎症反应的效果[11]。HMGB1是一种重要的晚期炎症介质,可由细胞直接破坏释放,也可以由免疫细胞分泌。在细胞迁移和炎性反应时,HMGB1是RAGE的生理性配体[12]。HMGB1/RAGE可通过激活核因子κB(NF-κB)促进炎性反应,进而诱导神经元凋亡[13]。研究报道PPARγ 激活可以调节HMGB1/RAGE信号通路,在急性肺损伤中发挥保护作用[14]。PPARγ是否可在神经细胞缺血再灌注损伤模型中,通过调节HMGB1/RAGE信号通路发挥保护作用,国内外缺乏相关报道。本研究发现罗格列酮使氧糖剥夺神经细胞RAGE表达下调,HMGB1分泌显著下降,而PPARγ受体抑制剂GW9662可以逆转该作用,提示罗格列酮通过PPARγ激活调控HMGB1分泌而发挥抗炎效应,并且可能通过调节HMGB1/RAGE信号通路发挥保护作用,但其具体机制值得深入研究。
蛋白激酶是近期研究较多的靶点,应激激活蛋白激酶(SAPK)包括c-Jun N末端蛋白激酶、p38丝裂原活化蛋白激酶,在脑缺血后被激活,抑制c-Jun N末端蛋白激酶或者p38丝裂原活化蛋白激酶,可以拮抗脑缺血后导致的细胞死亡[15~17]。近年来已有研究证实防止细胞凋亡是噻唑烷二酮类对于脑缺血损伤后神经保护的主要机制之一,罗格列酮通过下调含半胱氨酸的天冬氨酸蛋白水解酶而减少细胞凋亡,其机制涉及JNK和p38激酶的磷酸化作用[7]。并且,相关研究集中探讨蛋白质酪氨酸磷酸酶(DUSPs)对于蛋白激酶磷酸化作用的调控[7]。DUSP8可以特定作用于JNK和p38,有研究报道罗格列酮对脑缺血动物模型神经细胞死亡的保护作用,主要是通过促进DUSP8上调,从而阻止JNK磷酸化激活作用实现的[7]。
在本研究中,我们证实了使用罗格列酮作用于OGD/R神经细胞模型后, DUSP8蛋白和Bcl-xl表达均增加,而PPARγ受体抑制剂GW9662可以逆转该作用。从而肯定了以罗格列酮为代表的PPARγ受体激动剂通过上调DUSP8和Bcl-xl,对缺血和再灌注模型中神经细胞的死亡起到保护作用。
综上所述,我们证实了以罗格列酮为代表的PPARγ受体激动剂在神经细胞缺血和再灌注模型中可以通过减轻炎症反应和上调DUSP8发挥抗凋亡作用,而罗格列酮保护神经元的作用可以被PPARγ受体抑制剂逆转,进一步证实其作用机制是通过PPARγ受体激活实现的。但本研究尚缺乏对于JNK信号通道的检测,若能证实DUSP8蛋白升高,直接导致JNK磷酸化程度降低,从而减少细胞凋亡,则对于PPARγ受体激动剂的神经元保护机制,将提供更有力证据支持。因此,对于PPARγ受体激动剂的神经元保护机制的具体步骤,如对JNK信号通道的检测等,尚待深入探讨。
