F1产生2n种配子的细胞学基础
2019-05-25邢德智西安市临潼区代王初级中学陕西西安710611
邢德智 (西安市临潼区代王初级中学 陕西西安 710611)
遗传定律教学的主要任务是学习孟德尔应用“假设-演绎法”对豌豆的杂交实验,以及果蝇的遗传现象作出解释。 3 个遗传定律的发现过程,都是现代科研方法“假设-演绎法”的典型应用范例。而在遗传定律的教学中,演绎推理的关键环节之一是F1产生配子的种类和比例;应用数学知识探讨F1在减数分裂中,通过基因的自由重组和交换重组分别产生2n种次级性母细胞进而形成2n种配子,不但能帮助学生理解遗传定律的本质,而且使遗传定律在理论上数学化、严谨化和科学化,补充并丰富了连锁和交换定律理论。
1 遗传定律中F1 为什么可产生2n 种配子
1.1 孟德尔定律中F1产生2n种配子的细胞学基础 在减数分裂Ⅰ中期, 每对同源染色体排列在初级性母细胞赤道板的两侧, 多对同源染色体在赤道板两侧的排列方式, 决定了非同源染色体上非等位基因之间可能出现的组合趋势; 初级性母细胞分裂为次级性母细胞时, 因次级性母细胞里显性基因的种类和个数不同, 形成了若干种次级性母细胞; 又因为1 种次级性母细胞形成1 种配子,所以,次级性母细胞的种类总数=配子种类总数。若1 种次级性母细胞里没有显性基因(只有隐性基因)记作显性基因的个数=0;如果次级性母细胞里含有0 个、1 个、2 个、3 个显性基因……则每一种对应的类型种类数为的计算结果(n 表示等位基因对的总个数=1 个次级性母细胞里非等位基因的总个数=所研究的显性基因的总个数)。次级性母细胞的类型、各种类型数目和配子数目与等位基因对个数之间的关系见表1。
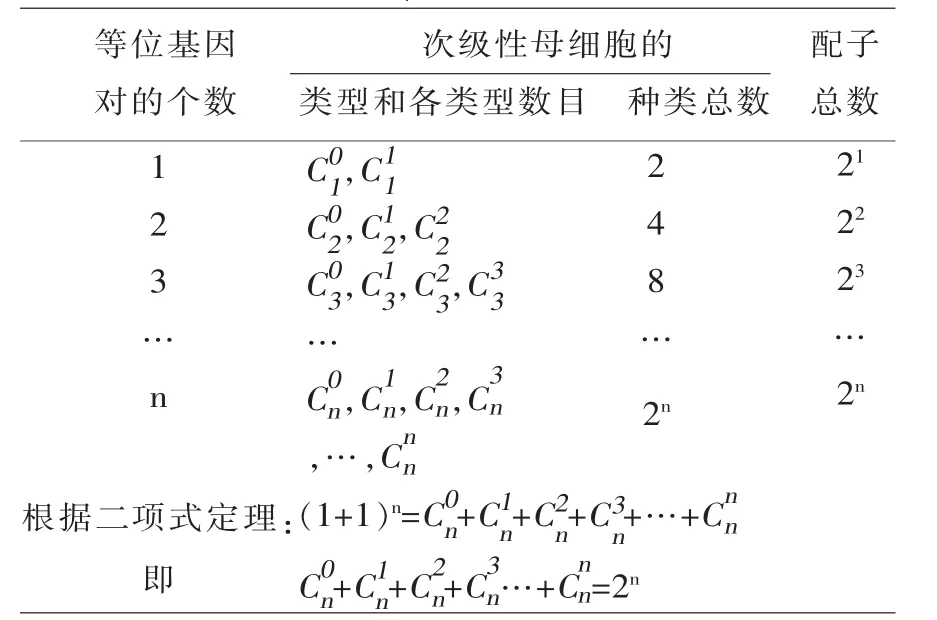
表1 孟德尔定律中F1 产生2n 种配子的细胞学基础
例如n=3 时,设F1的基因型为AaBbCc,则在减数分裂过程中形成的次级性母细胞里显性基因的类型和数目的详细情况如下:
1)含有0 个显性基因类型的数目。 含有0 个显性基因(也就是全部为隐性基因)的次级性母细胞基因组成只有abc 1 种。
2)含有1 个显性基因类型的数目。 含有1 个显性基因的次极性母细胞基因组成分别为Abc、aBc 和abC,共3 种。
3)含有2 个显性基因类型的数目。 含有2 个显性基因的次级性母细胞基因组成分别为ABc、AbC 和aBC,共3 种。
4)含有3 个显性基因类型的数目。 含有3 个显性基因(也就是全部为显性基因)的次级性母细胞基因组成只有ABC 1 种。
所以n=3 时,F1在减数分裂过程中形成次级性母细胞的类型数为4(1+1+1+1),种类数为8(1+3+3+1);种类数恰好与这4 类对应,种类数是其对应数学表示式的组合数;即一般规律中的(n+1)类2n种次级性母细胞进而形成2n 种配子。
如果用m 表示次级性母细胞里显性基因的个数,n 表示非等位基因的总个数,则次级性母细胞类型及类型种类数表示为:也可记作为;m=0,1,2,3,…,n),由于m 的个数等于(n+1),而所以,次级性母细胞共有(n+1)类2n种。 初级性母细胞分裂时,由于等位基因均等分配,非等位基因以同等的机会自由组合, 从而形成了种类不同但数目相等的次级性母细胞, 进而形成种类不同但数目相等的配子。1.2 连锁和交换定律中F1产生2n种配子的细胞学基础 2002年笔者通过数学建模、演算、归纳和推导得出:在不完全连锁遗传中F1可产生2n种配子,并证明了此结论[1]。近几年研究还发现在不完全连锁遗传中, 性母细胞类型及类型种类数与连锁基因个数和交换次数的关系。在连锁遗传中,如果n 表示等位基因对的个数(或连锁基因个数),k 表示交换次数, 则k 交换性母细胞的种类数为(n∈N*;n≥2;k =0,1,2,3,…,n-1)的组合数[2]。 如果等位基因对的个数依次取2,3,4,…,n;k 依次取0,1,2,3,…,n-1 时,得到和表1 类似的表2。

表2 不完全连锁遗传中F1 产生2n 种配子的细胞学基础
例如n=4 时, 设F1的 基 因型为++++/abcd(abcd 表示4 个连锁的隐性基因,+号表示对应的显性基因,a 为末端基因距着丝粒最远),则在减数分裂过程中因交换片断的种类和交换次数不同,最多可形成的初级性母细胞类型和数目的详细情况如下:
1)0 交换型的数目。 0 交换就是没有发生交换, 基因组成与F1的基因型完全相同, 即++++/abcd,0 交换型初级性母细胞只有1 种,之后只形成2 种“亲本型”配子。
2)单交换型的数目。包括单交换Ⅰ型(A-a 交换)1 种、单交换Ⅱ型(AB-ab 交换)1 种和单交换Ⅲ型(ABC-abc 交换)1 种,发生交换后的染色单体 的 基因 组 成 分 别 为:a+++/+bcd,ab++/++cd,abc+/+++d,单交换型初级性母细胞共3 种,之后共形成6 种“单交换重组型”配子。
3)双交换型的数目。包括双交换Ⅰ型(A-a 交换后再交换aB-Ab)1 种、双交换Ⅱ型(A-a 交换后再交换aBC-Abc)1 种和双交换Ⅲ型(AB-ab 交换后再交换abC-ABc)1 种,连锁基因发生2 次不同的单交换后的染色单体的基因组成分别为:+b++/a+cd,+bc+/a++d,++c+/ab+d,双交换型初级性母细胞共3 种,之后共形成6 种“双交换重组型”配子。
4)三交换型的数目。 三交换就是在任意1 种双交换+b++/a+cd,+bc+/a++d 和++c+/ab+d 的基础上再发生1 次单交换, 且交换结果是一种新的类型,这种结果只有1 种,即a+c+/+b+d。 三交换型初级性母细胞只有1 种,之后只形成2 种“三交换重组型”配子。
所以n=4 时,F1在减数分裂过程中形成初级性母细胞的类型为4(1+1+1+1)类,种类数为8(1+3+3+1)种;种类数恰好与这4类对应,种类数是其对应数学表示式的组合数;即一般规律中的n 类2n-1种初级性母细胞最终也可形成2n种配子。 虽然数学形式与自由组合定律相似,但是生物学意义却不同。
在不完全连锁遗传中, 初级性母细胞类型及类型种类数表示为也可记作为;k=0,1,2,3,…,n-1),由于k=0,1,2,3,…,n-1,初级性母细胞类型总数最多等于n,而所以,最多共有n 类2n-1种初级性母细胞。
1.3 孟德尔定律与连锁和交换定律中F1产生2n种配子细胞学基础的异同 连锁和交换定律中F1产生2n种配子的机理虽然与自由组合定律不同,但却有异曲同工之妙,其相同或相似之处为:
1)在孟德尔定律中,由于次级性母细胞中显性基因的种类和数目不同,共形成(n+1)类次级性母细胞,(n+1)类次级性母细胞种类数恰好为n次二项式展开式的系数,其和为2n,所以2n种次级性母细胞进而形成2n种配子。 在连锁和交换定律中, 因初级性母细胞中染色单体交换片断的种类及交换次数不同,共形成n 类初级性母细胞,且n 类初级性母细胞种类数恰好为(n-1)次二项式展开式的系数, 其和为2n-1,2n-1种初级性母细胞形成2n种次级性母细胞进而形成2n种配子。
3)在孟德尔定律及连锁和交换定律中,次级性母细胞类型和初级性母细胞类型分别可用和(n∈N*;n≥2;k=0,1,2,3,…,n-1)展开式得出。
不同之处:
1)自由组合定律中,等位基因一定要分离,非等位基因一定要自由组合, 所以基因重组的概率几乎为100%,或者说“重组”是必然的,因此,理论上F1一定产生2n种配子;又因为非等位基因之间的组合是互不干扰且机会是均等的,所以,形成各种次级性母细胞亦即各种配子的几率相同,因此,配子种类不同但数目相等。在连锁和交换定律中,由于各种交换具有很大的偶然性,并且即使发生了交换,也不一定形成重组型配子;例如三点测验中(n=3),多数情况下F1可形成23,即8 种配子,少数情况下只形成6 种配子, 这是因为发生了干涉,且“干涉” 是完全的,1 个双交换型性母细胞也没有;不完全连锁形成各种“重组型”配子具有很大的偶然性,交叉互换是基因重组的必要条件而非充分条件,因此,F1产生2n种配子只是一种理论上的可能。又因为交换片断的种类及交换次数的频率均不相同,所以,形成配子的种类不同,数目也不相等。 交换重组后形成配子的种类和比例,决定于交换性母细胞的种类和其对应的交换率。
2)孟德尔定律中次级性母细胞类型数目的计算公式, 与连锁和交换定律中初级性母细胞类型数目的计算公式,虽然数学形式相似,但其实际意义不同。
孟德尔定律中,初级性母细胞分裂时,由于等位基因分离,非等位基因自由组合,从而形成的次级性母细胞中含有n 个不同的非等位基因;…分别表示次级性母细胞里含有0 个、1 个、2 个、3 个显性基因……次级性母细胞里含n个不同的非等位基因, 其中显性基因是0 个的种类明显只有1 种,1 个是显性基因的有n 种…这里所表示的意义与数学上基本一致; 各类次级性母细胞种类的数值就是其在数学上分别计算的结果。
在连锁和交换定律中,初级性母细胞类型数目计算公式的实际意义是:假设,初级性母细胞里染色单体上的连锁基因均在着丝粒的一端,且始终为二线交换,同时不考虑染色体干涉;则不论交换与否或交换多少次,每条染色单体上的连锁基因总是n 个,最多可有(n-1)种交换片断进行交换。
双交换和多交换不但有二线交换还有三线交换及四线交换, 但是三线交换和四线交换形成的配子种类包含在0,1,2,3,…k 次二线交换所形成的配子种类中; 所以每1 种二线交换的初级性母细胞净增加2 种配子;高次交换除形成“高次交换型”配子外,还形成“低次交换型”配子,包括“亲本型”配子。
1.4 基因自由重组与交换重组的本质 F1产生2n种配子的推导计算过程, 可从数学上理解什么是基因的自由组合,什么是基因的交换重组,以及基因自由重组和交换重组的本质。
自由组合就是所有非等位基因之间的组合;通过每一对等位基因分离的独立性、等位基因对之间非等位基因组合时的选择性而实现; 即从每一对等位基因中任选1 个基因形成的组合, 共2n种;所以,自由组合的前提条件是每对同源染色体上的2 个基因要具有选择性, 即每对同源染色体上的2 个基因都是等位基因,且n≥2,非等位基因组合的结果是一定形成2n种配子。
推理计算过程表明,基因交换重组的本质是:基因连锁是基因交换的基础; 双交换是通过2 次不同的单交换实现的, 多交换是通过多次不同的单交换实现的,单交换是其他交换的基础。
自由重组是染色体间的重组, 交换重组是染色体内的重组。
2 3 个遗传定律的联系
2.1 分离定律与自由组合定律 分离定律是基础,自由组合定律是分离定律的拓展和延伸,二者是“量”上的区别,常统称为孟德尔定律。 当只研究1 对等位基因时, 形成的2 种次级性母细胞里各自只有1 个基因, 不存在非等位基因的自由组合,所以,F1只形成2 种配子;分离定律是自由组合定律在1 对等位基因时的特殊表现。 当研究2对或多对等位基因时,由于所有等位基因分离,导致了次级性母细胞里非等位基因的自由组合,所以,F1共形成2n种配子。
2.2 分离定律与连锁和交换定律 连锁和交换定律也是对分离定律的拓展和延伸, 与分离定律不但有“量”的不同,而且有“质”的区别。 设RF 表示重组率,当没有交换即RF=0 时,为完全连锁;完全连锁与分离定律相似,F1只形成2 种配子。当0<RF<50%时,表现为不完全连锁,是连锁遗传中最常见的形式;不完全连锁特征为:①F1产生的初级性母细胞一部分发生了交换、 一部分未发生交换,②F1产生的配子中“亲本型”配子多,“重组型”配子少。
2.3 自由组合定律与连锁和交换定律 自由组合定律与连锁和交换定律都是研究2 对或多对相对性状遗传的。当研究2 对相对性状时,自由组合定律中的重组率=50%; 连锁和交换定律中,Haldane 作图函数RF=1/2(1-e-2x)和Haldane 曲线的性质均反映出:当2 个基因图距很大时,重组率接近50%[3]。这时的基因交换恰似自由组合,或者说表现为自由组合, 其实将其称为完全交换(100%的性母细胞都在2 个基因之间发生了交换)更合适[4]。 所以,重组率接近或等于50%时,其遗传形式应为完全交换或自由组合[5]。
3 小结
在孟德尔定律中, 等位基因分离导致非等位基因自由组合, 从而形成2n种次级性母细胞,是形成2n种配子的细胞学基础;用排列组合的知识理解为什么形成2n种次级性母细胞,可加深对孟德尔定律本质的理解。在连锁和交换定律中,等位基因交换导致非姐妹染色单体上连锁基因的重新组合,从而最多形成2n-1种初级性母细胞,是最多形成2n种配子的细胞学基础;同理,用排列组合的知识理解双交换、 多交换都是在单交换的基础上进行的, 各类交换最多共形成2n-1种染色单体“对”,亦即2n-1种初级性母细胞,是理解连锁和交换定律本质的关键。
在连锁和交换定律中,配子种类,特别是比例的形成较为复杂,探究性极强,是中学生应用“假设-演绎法”再发现科学定律的极好素材。
