多主棒孢SdhB-H278R突变位点AS-real-time PCR定量检测体系的建立
2018-12-27孙炳学石延霞朱发娣谢学文柴阿丽李宝聚
孙炳学,石延霞,朱发娣,谢学文,柴阿丽,李宝聚
多主棒孢SdhB-H278R突变位点AS-real-time PCR定量检测体系的建立
孙炳学,石延霞,朱发娣,谢学文,柴阿丽,李宝聚
(中国农业科学院蔬菜花卉研究所,北京 100081)
【目的】建立一种快速、高效、定量检测黄瓜多主棒孢()琥珀酸脱氢酶B亚基()H278R突变的实时荧光定量PCR(AS-real-time PCR)检测方法,并进行效果验证。【方法】从北京市大兴区采集、分离纯化病原菌,获得24株多主棒孢的单孢菌株,采用菌丝生长速率法测定其对啶酰菌胺的EC50,随机选取4株敏感(S)和8株抗性(R)菌株测其菌丝生长速率、产孢量和致病性。采用引物对Cc-SdhB-F/R测定多主棒孢基因序列,检测的碱基变化。基于多主棒孢的相同核酸序列和SNP位点设计内参引物B-H278R-TY-F/R和特异性引物B-H278R-2F/2R14,建立并优化AS-real-time PCR定量检测体系,并对引物的特异性、熔解曲线和灵敏度进行评价。利用该体系分别检测含有H278R不同突变比例的DNA和孢子悬浮液。【结果】测定的24个菌株中,敏感菌株占66.7%,EC50值为0.057—0.563 μg·mL-1;抗性菌株占33.3%,EC50值为5.395—11.710 μg·mL-1。抗性和敏感菌株仅在菌丝生长速率方面存在显著性差异,且菌丝生长速率与EC50之间存在显著负相关。在产孢量和致病性方面不存在显著性差异。测序分析发现抗性菌株均携带SdhB-H278R突变。本研究设计的引物B-H278R-2F和B-H278R-2F2R14特异性强,仅对多主棒孢SdhB-H278R突变菌株的DNA有扩增条带,对其余供试菌株均无条带扩增。普通AS-PCR检测的灵敏度为91 pg·μL-1, 而AS-real-time PCR的灵敏度可达9.1 pg·μL-1,灵敏度为普通AS-PCR的10倍。以基因组DNA为标准品,构建的AS-real-time PCR标准曲线ΔCT值与相对模板浓度的对数有良好的线性关系,相关系数为0.9857, 扩增效率为92.59%。熔解曲线吸收峰单一,内参引物B-H278R-TY-F/R和特异性引物B-H278R-2F/2R14分别在87.81℃和91.62℃处出现单一特异性峰。用DNA和孢子悬浮液对标准曲线进行验证,结果表明随着相对模板浓度逐渐降低,该检测体系的准确性逐渐升高且检测下限为5%,同时预期值与试验值呈线性相关(2=0.9998和2=0.9922)。【结论】建立了一种高效、定量、快速的AS-real-time PCR检测体系用于SdhB-H278R突变位点的检测,可为杀菌剂的抗性治理提供理论依据。
黄瓜;多主棒孢;AS-real-time PCR;抗药性;琥珀酸脱氢酶B亚基;啶酰菌胺
0 引言
【研究意义】多主棒孢()隶属于格孢菌目(Pleosporales)棒孢科(Corynesporascaceae)棒孢属(),由其引起的黄瓜棒孢叶斑病发生普遍,该病害在温室、大棚和露地中均不断加重,已成为黄瓜栽培中主要的病害之一[1]。目前大多采用化学药剂防治黄瓜棒孢叶斑病,由于多主棒孢繁殖速度快、遗传变异大,加之多年连续使用单一药剂防治,导致其对苯并咪唑类、二羧酰亚胺类、-苯氨基甲酸酯类和QoI类的大部分杀菌剂产生了抗药性[2-3]。啶酰菌胺是德国巴斯夫公司1996年开发的新型烟酰胺类内吸性杀菌剂,该药剂能够阻断电子在复合体II琥珀酸脱氢酶[3Fe-4S]结构域与泛醌之间的传递,从而影响病原真菌的电子传递,阻碍病原菌的能量代谢[3-4]。因该药剂作用位点单一,FRAC已将其归为中度抗性风险杀菌剂。自2008年啶酰菌胺在中国登记防治灰霉病并在黄瓜栽培中大量使用,间接导致多主棒孢对啶酰菌胺产生抗药性,出现大量的抗性突变株,因此,建立一种快速、准确、定量的多主棒孢SdhB-H278R突变检测方法,对监测啶酰菌胺田间的抗性频率和病害防治具有重要意义。【前人研究进展】随着啶酰菌胺的广泛使用,在不同蔬菜和作物上出现了对啶酰菌胺有抗性的病原菌。2007年,首次在开心果上发现了对啶酰菌胺产生抗性的链格孢()[5],之后在灰葡萄孢()、小双胞腔菌()和核盘菌()上也发现了对啶酰菌胺产生抗性的菌株[6-8]。2009年,Miyamoto等[9]在黄瓜上发现了对啶酰菌胺产生抗性的多主棒孢并在2010[10]年发现了SdhB-H278R/Y、SdhC-C73P、SdhD-S89P和SdhD-G109V等多种抗性突变类型,且SdhB-H278R基因型在田间的频率为0.64%。传统的抗药性检测方法包括离体测定和活体测定法,其中离体测定法主要有菌落生长速率法和孢子萌发法,均是通过测定药剂对植物病原菌的EC50值以区分敏感和抗性菌株,比较费时费力。近年来,随着分子生物学技术的发展,PCR-RFLP(restriction fragment length polymorphism PCR)、AS-PCR(allele specific PCR)、AS-real-time PCR、HRM(high resolution melting)和多重等位基因PCR等分子鉴定手段广泛应用于植物病原菌对药剂的抗药性检测。Furuya等[11]和Aoki等[12]在霜霉病菌()上分别建立了对QoI杀菌剂抗性Cytb-G143A和对CAA杀菌剂PvCesA3-G1105S突变位点PCR-RFLP的检测体系。基于等位基因特异性核苷酸引物,AS-PCR和AS-real-time PCR抗药性检测技术已在核盘菌、茄链格孢()、大麦云纹病菌()和灰葡萄孢等病原菌上应用[13-16]。最近,新兴的HRM抗药性检测技术因具有高通量、快速的特点并可用于SNP位点的筛选,已开始在核盘菌对甲基硫菌灵和灰葡萄孢对环酰菌胺以及啶酰菌胺的抗药性检测上应用[17-18],准确性高达100%,但该方法只能定性检测。【本研究切入点】目前为止尚未建立一种检测多主棒孢抗性突变位点的检测体系,基于笔者课题组前期发现的一种抗啶酰菌胺的黄瓜多主棒孢SdhB-H278R抗性突变位点,使用荧光定量PCR仪,建立一种定量、高效、快速检测该突变位点的AS-real-time PCR体系,并对此体系的特异性、可重复性和灵敏度进行评价。【拟解决的关键问题】建立一种定量、高效、快速检测黄瓜多主棒孢SdhB-H278R突变位点的AS-real-time PCR体系,以期为该病原菌对啶酰菌胺的田间抗性检测和病害防控提供更好的技术支持。
1 材料与方法
试验于2017年3月至2018年4月在中国农业科学院蔬菜花卉研究所完成。
1.1 试验材料
1.1.1 菌株 2017年3月从北京市大兴区采集黄瓜多主棒孢叶斑病病样,在室内进行分离培养,单孢纯化后得到24株多主棒孢菌株,其他5株多主棒孢菌株采自河北和辽宁。所有菌株由中国农业科学院蔬菜花卉研究所蔬菜病害综合防治课题组提供。
1.1.2 试剂和培养基 96%啶酰菌胺原药(陕西美邦农药有限公司)、植物组织DNA提取试剂盒、SuperReal PreMix Plus(天根生化科技(北京)有限公司),2×Taq MasterMix (北京博迈德生物技术有限公司)、PDA培养基、YBA培养基。
1.1.3 主要仪器设备 NanoDrop2000超微量分光光度计(Thermo Fisher Scientific Inc.,USA)、3-18K型离心机(Sigma,德国)、7500 Real time PCR system(ABI,美国)、S1000 Thermal Cycle PCR仪(BIO- RAD,美国)。
1.1.4 引物合成测序 荧光定量PCR和普通PCR所用引物及基因序列由北京博迈德基因技术有限公司合成并测定。
1.2 方法
1.2.1 多主棒孢对啶酰菌胺敏感性的测定 参照Miyamoto等[9]方法,采用菌丝生长速率法测定北京大兴24株及河北、辽宁的5株多主棒孢对啶酰菌胺的敏感性。抗药性测定设置7个浓度梯度(0、0.01、0.1、1、5、10、30 μg·mL-1),以不加药剂为空白对照;在新鲜的菌落边缘打取直径5 mm的菌饼,接种于含药培养基的平板上,在26℃黑暗培养5 d,测量菌落直径,每个处理重复3次。比较试验组和对照组的菌落直径,并计算EC50值、毒力回归方程和相关系数。根据Miyamoto等[9]的敏感基线(0.04—0.59 μg·mL-1)判定多主棒孢对啶酰菌胺的敏感性。
1.2.2 抗性和敏感菌株的生物学特性测定 菌丝生长速率的测定:从24株多主棒孢菌中随机选取4株敏感菌株和8株抗性菌株作为测定生物学特性的菌株。在多主棒孢菌生长5 d的PDA平板上制备5 mm的菌饼,采用灭菌牙签将菌碟菌丝面朝下贴于PDA平板中央,密封后置于26℃培养。分别于第2—11天,采用十字交叉法测量菌落生长直径。每个菌株3次重复,试验重复2次。
产孢能力的测定:待多主棒孢菌生长至第11天,用直径5 mm的打孔器在菌落边缘打取5个菌饼置于1 mL的无菌水中,加入5个直径2 mm的研磨珠用涡旋振荡机振荡2 min洗脱分生孢子,用血球计数板计数计算分生孢子的浓度。每个菌株3次重复,试验重复2次。
致病性的测定:采集离体组织测定法,采集叶龄一致,形状、大小相似的健康黄瓜叶片,用灭菌的接种针刺破形成微伤口。将制备好直径5 mm的菌饼分别接种于黄瓜叶片,接种后放置于保鲜盒中,置于相对湿度85%,自然光照的26℃温室中,培养5 d后采用十字交叉法测量病斑直径。每个菌株测6片叶片,试验重复2次。
1.2.3 DNA提取 将多主棒孢接种到PDA平板培养5 d,刮取菌丝作为DNA提取样本。采取植物组织DNA提取试剂盒,根据试剂盒中的说明书提取DNA,提取后用NanoDrop 2000c测定DNA浓度,用ddH2O稀释至40 ng·μL-1,4℃保存备用。
1.2.4 AS-real-time PCR定量检测体系的建立序列测定:根据GenBank多主棒孢序列(AB548739.1), 应用Primer Premier 5.0设计引物Cc-SdhB-F和Cc-SdhB-R(表1),以表2中所有多主棒孢的DNA为模板,用Cc-SdhB-F和Cc-SdhB-R引物扩增基因片段,PCR产物进行测序。50 μL PCR体系:2×Taq PCR MasterMix 25 μL,Cc-SdhB-F和Cc-SdhB-R引物2 μL,模板DNA 2 μL,ddH2O补足50 μL。PCR反应程序:94℃ 5 min,94℃ 45 s,58℃ 45 s,72℃ 90 s,步骤2—4进行30个循环,72℃ 10 min。
引物设计:根据的测序信息,针对SNP位点和相同的序列信息,用Primer Premier 5.0设计特异性引物B-H278R-2F/2R14和内参引物B-H278R- TY-F/R(表1),引物设计策略和引物信息如图1和表1所示。为增加引物的特异性,引物3′端与突变碱基G配对,在靠近3′端的第2、3位引入错配碱基[13-16,19]。
通过对全球自动驾驶技术领域专利市场进行研究,发现当前自动驾驶技术专利申请数量逐年上升,研发热情逐年高涨。但从技术角度出发,目前从L3有条件自动驾驶阶段迈向未来L5完全无人驾驶阶段仍需要很长的过程和时间。本文从专利时间分布、专利空间分布、专利内容分布这三大角度出发,全面揭示了全球自动驾驶技术的发展趋势、技术公开国家、技术来源国家、技术研发机构、技术研究方向、研究热点等情况,最终总结出以下6点结论,期望为自动驾驶技术的发展提供决策参考。

图1 引物设计策略

表1 引物信息
单下划线Single underline:突变位点Mutation site;双下划线double underline:人为错配碱基additional mismatched base
引物特异性检测:以含有H278R突变、其他点突变菌株以及常见的8种植物病原菌DNA为模板,以ddH2O为阴性对照,用引物B-H278R-2F和B-H278R-2R14进行PCR扩增,检测引物特异性。20 μL PCR反应体系:2×Taq PCR MasterMix 10 μL,引物B-H278R-2F和B-H278R-2R14 0.4 μL,模板DNA 1.6 μL,ddH2O补足10 μL。PCR反应程序:94℃ 5 min,94℃ 45 s,65℃ 45 s,72℃ 30 s,步骤2—4进行35个循环,72℃ 10 min。PCR产物经1%琼脂糖凝胶电泳检测后,凝胶成像系统分析结果。
标准曲线的建立:为了定量检测多主棒孢SdhB- H278R突变株在田间的比例,以含有H278R突变菌株(R278)和不含有H278R突变菌株(H278)的混合DNA作为模板,以R278 DNA摩尔质量分别占总量DNA的6.25%、12.5%、25%、50%、75%和100%作为不同的处理,3次重复,分别用特异性引物B-H278R-2F/2R14和内参引物B-H278R-TY-F/R在7500 real-time PCR system仪中进行AS-real-time PCR,计算ΔCT(ΔCT=CT特异性引物-CT内参引物),以ΔCT值为纵坐标,以R278 DNA相对摩尔质量的对数为横坐标,制作标准曲线。
20 μL反应体系:2×SuperRealPreMix Plus 10 μL,50×ROX Reference Dye 0.4 μL,上游引物和下游引物各1 μL,模板DNA 40 pg,RNase-free ddH2O补足20 μL,AS-real-time PCR反应程序:95℃ 15 min;95℃ 10 s;60℃ 32 s,40个循环;95℃ 15 s;60℃ 1 min;60℃开始,每个循环增加0.35℃,100个循环,60℃ 15 s。
普通PCR和AS-real-time PCR灵敏度检测:将定量的R278基因组DNA按浓度进行10倍梯度稀释,用引物B-H278R-2F和B-H278R-2R14进行普通PCR和AS-real-time PCR扩增,检测引物的灵敏度。
1.2.5 AS-real-time PCR定量检测体系的验证 DNA验证:以R278和H278混合DNA作为模板,以R278 DNA摩尔质量分别占总量DNA的80%、40%、20%、10%和5%作为不同的处理,3次重复。按照已建立的AS-real-time PCR体系验证标准曲线的可行性。用Excel计算标准差以及与预期值和检测值之间的线性相关性。
孢子悬浮液验证:将R278和H278菌株26℃下培养11 d,用0.1%的吐温20刷下孢子悬浮液并用三层纱布过滤得到孢子悬浮液。用血球计数板调节孢子浓度为1×105个/mL,配置混合孢子悬浮液,其中以H278孢子数量分别占混合孢子数的80%、40%、20%、10%和5%作为不同的处理,12 000 r/min离心,收集沉淀,用植物组织DNA提取试剂盒提取不同处理的DNA。按照已建立的AS-real-time PCR体系验证标准曲线的可行性。用Excel计算标准差以及与预期值和检测值之间的线性相关性。
2 结果
2.1 多主棒孢对啶酰菌胺的敏感性测定
采用菌丝生长速率法测定采集的多主棒孢对啶酰菌胺的敏感性,结果表明,采自北京的24株多主棒孢分为敏感和抗性2个群体,其中敏感菌株16株,占66.7%,其EC50值为0.057—0.563 μg·mL-1,平均值为0.368 μg·mL-1。抗性菌株8株,占33.3%,其EC50值为5.395—11.710 μg·mL-1,平均值为7.586 μg·mL-1(图2、表2)。
2.2 抗性和敏感菌株的生物学特性
随机选取的12株多主棒孢之间的平均菌丝生长速率存在差异,敏感菌株菌丝生长速率为0.49—0.61 cm·d-1,抗性菌株菌丝生长速率为0.30—0.56 cm·d-1(表3)。与敏感菌株相比,抗性菌株除第9天的菌丝生长速率相等外,其他时间段均比敏感菌株生长慢(图3),且抗性菌株的平均生长速率与敏感菌株差异极显著(<0.01)。菌丝生长速率与EC50之间呈显著负相关(表4)。
由表3可见,敏感菌株的产孢量在0.33×105—7.00×105个/mL,抗性菌株的产孢量在1.33×105—7.46×105个/mL,抗性菌株和敏感菌株在产孢量上不存在显著差异(0.949)。产孢量与生长速率和致病性存在低度相关,但不显著(表4)。
敏感菌株的病斑直径在0.38—0.84 cm,抗性菌株的病斑直径在0.33—0.91 cm。抗性和敏感菌株在致病性上不存在显著差异(表3)。致病性与产孢量和生长速率之间存在低度相关性,但不显著(表4)。

图2 24株多主棒孢菌的敏感性分布

图3 抗性和敏感菌株菌丝生长速率比较
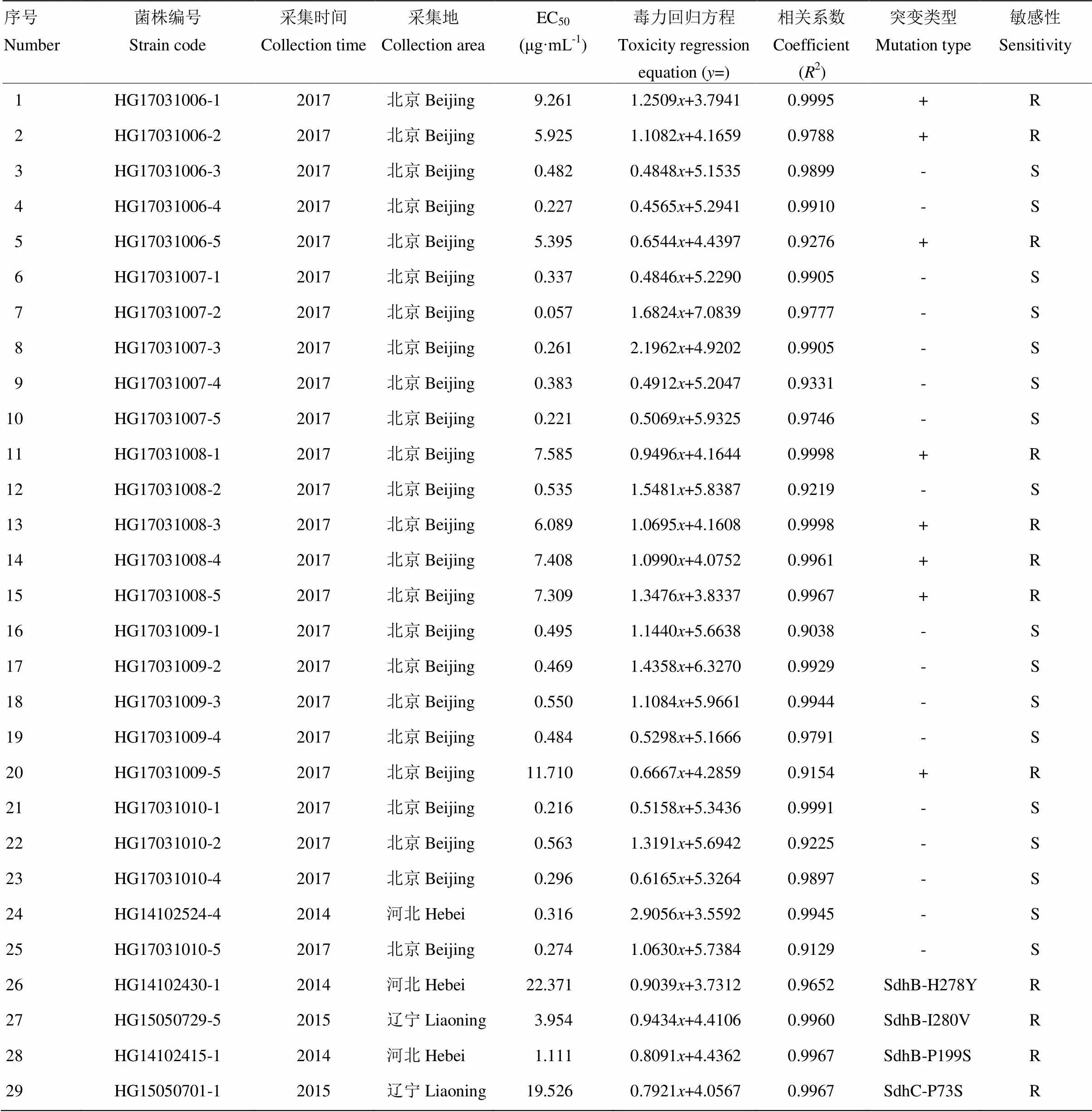
表2 供试菌株敏感性测定
+:有SdhB-H278R突变Carried SdhB-H278R mutation;-:无SdhB-H278R突变No SdhB-H278R mutation;R:抗性菌株resistant strain;S:敏感菌株sensitive strain
2.3 AS-real-time PCR定量检测体系的建立
应用引物Cc-SdhB-F和Cc-SdhB-R测定的序列,结果如图4 所示,抗性菌株(R)在第956位的A突变到G,导致在密码子的第278组氨酸(H)突变成精氨酸(R),进一步发现含有H278R突变的菌株对啶酰菌胺具有抗性。
以携带不同点突变的多主棒孢和8株常见的病原菌基因组DNA为模板,采用引物B-H278R-2F和B-H278R-2R14进行特异性验证(图5),结果表明仅有携带SdhB-H278R突变的多主棒孢基因组DNA有扩增条带,大小为244 bp,携带其他点突变的多主棒孢以及8种病原菌DNA均未检测到扩增条带,可用于不同菌种间多主棒孢H278R基因型的检测。
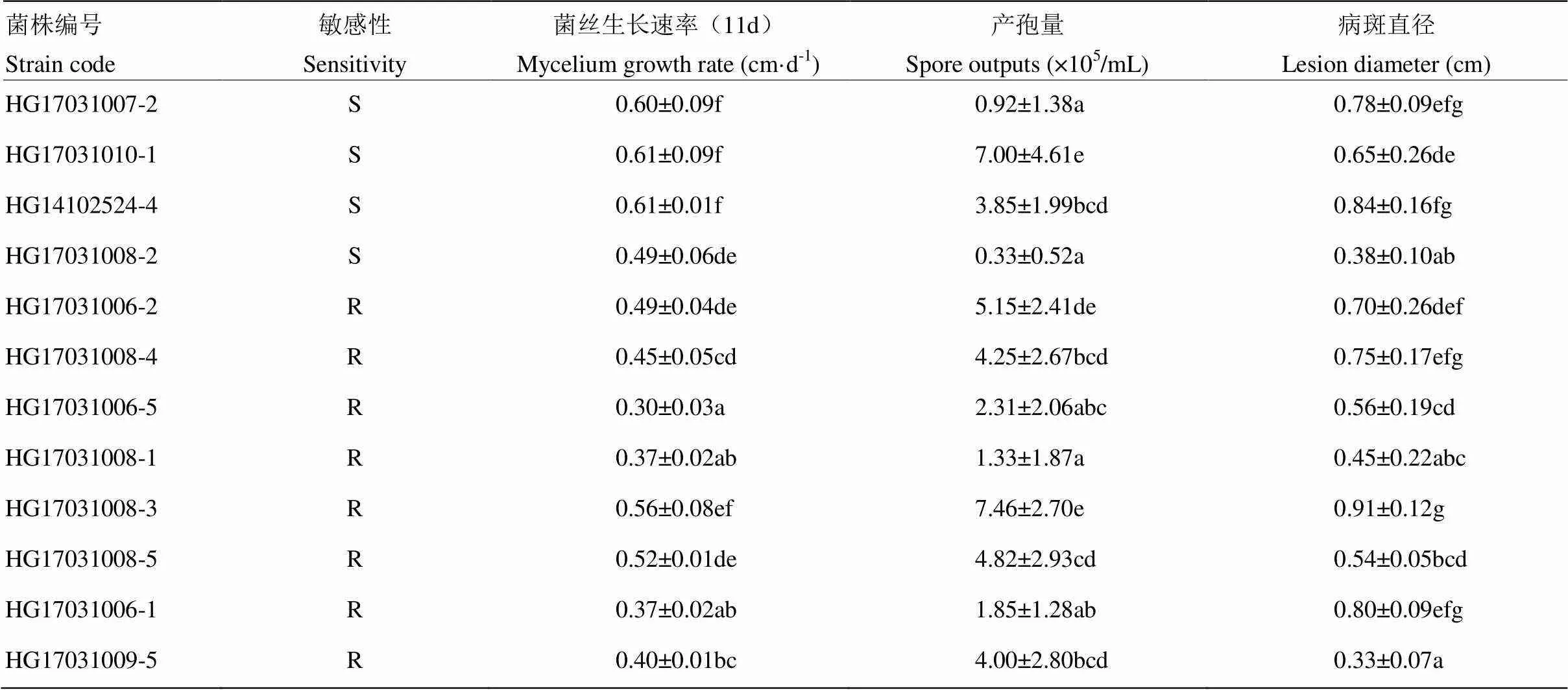
表3 抗性和敏感菌株的生物学特性
表中数据为平均值±标准差。表5同Data aremean±standard deviation. The same as Table 5根据Duncan’s最小显著性差异测定,同列数值后不同字母表示差异显著(<0.05)According to a Duncan’s least-significant difference, different lowercases after data in the same column indicate significant difference (<0.05)

图4 在SdhB上抗性和敏感菌株的核酸序列和氨基酸序列信息

M:分子量标准BM5000 Molecular weight marker BM5000;1:SdhB-H278R突变SdhB-H278R mutation;2:无突变no mutation;3:SdhB-H278Y突变SdhB-H278Y mutation;4:SdhB-I280V突变SdhB-I280V mutation;5:SdhB-P199S突变SdhB-P199S mutation;6:SdhC-S73P突变SdhC-S73P mutation;7:黄瓜叶片cucumber leaf;8:灰葡萄孢B. cinerea;9:瓜枝孢Cladosporium cucnerinu;10:立枯丝核菌Rhizoctonia solani;11:茄链格孢A. solani;12:茄葡柄霉Stemphylium solani;13:尖镰孢Fusarium oxysporum;14:核盘菌S. sclerotiorum;15:古巴假霜霉菌Pseudoperonospora cubensis;16:ddH2O
利用10倍梯度稀释的基因组DNA对引物B-H278R-2F和B-H278R-2R14进行普通PCR和AS-real-time PCR灵敏度检测,结果表明普通PCR检测的灵敏度为91 pg·μL-1(图6-A),AS-real-time PCR检测的灵敏度为9.1 pg·μL-1(图6-B)。AS-real-time PCR的灵敏度为普通PCR的10倍。

表4 EC50与生物学特性的相关性分析
表中数据为Pearson相关系数Data in the table are Pearson correlation coefficient values;*:<0.05

A:普通PCR灵敏度检测sensitivity detection with ordinary PCR;B:AS-real-time PCR灵敏度检测sensitivity detection with AS-real-time PCR;M:分子量标准BM5000 Molecular weight marker BM5000;1—8:DNA稀释为9.1、9.1×10-1、9.1×10-2、9.1×10-3、9.1×10-4、9.1×10-5、9.1×10-6、9.1×10-7 ng·μL-1 DNA diluted to 9.1, 9.1×10-1, 9.1×10-2, 9.1×10-3, 9.1×10-4, 9.1×10-5, 9.1×10-6, 9.1×10-7 ng·μL-1;9:ddH2O
利用优化的条件进行AS-real-time PCR,特异性引物B-H278R-2F/2R14扩增产物在熔解温度为91.62℃处出现单一的特异性峰(图7-A),内参引物B-H278R- TY-F/R扩增产物在熔解温度为87.81℃处出现单一的特异性峰(图7-B)。
以R278 DNA摩尔质量分别占总量DNA的6.25%、12.5%、25%、50%、75%和100%作为不同的处理,用引物B-H278R-2F/2R14、B-H278R-TY/R进行AS- real-time PCR检测,建立了标准曲线=-3.513+9.807,斜率为-3.513,截距为9.807,相关系数2=0.9857,扩增效率为92.59%。R278 DNA占总DNA的6.25%—100%的比例中,呈现出良好的线性关系(图8)。
2.4 AS-real-time PCR定量检测体系的验证
以R278菌株DNA摩尔质量和孢子数量分别占总量的80%、40%、20%、10%和5%作为不同的处理,进行AS-real-time PCR检测并验证标准曲线的可行性。结果显示预期值为80%、40%、20%、10%和5%时,相应的DNA检测值依次为85.95%、44.64%、22.24%、11.86%和6.40%,孢子检测值依次为116.45%、48.39%、26.38%、11.86%和6.83%(表5)。随着预期值逐渐降低检测值的标准差也依次下降,且预期值在5%时具有最小的标准差。同时预期值和检测值具有很高的相关性(2=0.9998和2=0.9922),说明此检测体系具有可行性,在检测SdhB-H278R突变比例为5%时具有更高的准确性。

图8 AS-real-time PCR建立检测SdhB-H278R突变的标准曲线
3 讨论
多主棒孢是一种重要的植物病原菌,是棒孢属内发现最早、寄主范围最广的种,可侵染葫芦科、茄科和豆科等大部分蔬菜,园艺花卉,橡胶等经济作物以及大豆等粮食作物,约380属530多种植物[20]。黄瓜棒孢叶斑病在我国北京、山东、甘肃、河北、广东和黑龙江等多个省份[21-23]以及在日本和韩国[9,10,24]均大范围发生,一般田间发病率为10%—25%,严重时可达60%—70%,甚至100%。因此由多主棒孢引起的黄瓜棒孢叶斑病在黄瓜上已成为一种重要的病害。啶酰菌胺是德国巴斯夫公司开发的一种SDHIs类杀菌剂,自2006年开始在日本登记防治黄瓜棒孢叶斑病,随后很快出现了对啶酰菌胺的抗药性[9-10]。但在国内该药剂并未登记在防治黄瓜棒孢叶斑病上,只登记用于防治黄瓜灰霉病,随着啶酰菌胺在防治黄瓜灰霉病中的广泛使用,灰葡萄孢对啶酰菌胺产生了抗药性[25],间接地多主棒孢对啶酰菌胺也相应产生了抗药性,导致黄瓜棒孢叶斑病大量发生。目前多主棒孢上的抗性突变包括SdhB-P199S、H278Y/R、I280V、SdhC-S73P、SdhD-S89P、D95E、H108R、G109V[10],而H278R为首次在国内报道,因此建立一种AS-real-time PCR检测方法非常必要,可用于指导后续的抗性治理。若H278R抗性频率升高,应使该药剂减少甚至停止使用,并采用不同作用机制的杀菌剂轮换使用或具有负交互抗药性药剂复配使用,可延长啶酰菌胺的使用寿命,减少黄瓜棒孢叶斑病的发生。

表5 AS-real-time PCR验证
通过测定在北京市大兴区采集的黄瓜多主棒孢对啶酰菌胺的敏感性并参考Miyamoto等[9]的研究结果,将多主棒孢菌株分为敏感和抗性菌株两个群体,通过对黄瓜多主棒孢序列的测定,抗性菌株均为SdhB-H278R突变。Miyamoto等[9]检测到SdhB-H278R共4株且划分为高等抗性菌株,EC50范围为8.9—10.7 μg·mL-1,而本文检测到SdhB-H278R突变共8株且EC50范围更广(5.395—11.710 μg·mL-1),推测可能与检测到H278R基因型的数量有关。通过对抗性和敏感菌株的生物学特性分析,发现抗性和敏感菌株除在菌丝生长速率方面存在显著差异外,在产孢量和致病性方面差异不显著,说明H278R突变株与敏感菌株之间的田间适合度差异不大,在药剂选择压力下易发展成优势群体导致黄瓜叶斑病的大量发生。-H272Y/R/L、H277Y/R/Y和H278R/Y几种点突变类型已经在小双胞腔菌、链格孢和灰葡萄孢等多种病原菌中被报道[7,14,16,26-28]。这几种点突变均发生在泛醌结合口袋及其附近[29],且氨基酸序列具有高度的保守性,可以将这几种突变归为一类。LALÈVE等[30]研究发现,灰葡萄孢H272R/L/Y突变在以葡萄糖作为碳源时,前期菌丝生长速率较慢,后期一致;另外H272Y突变不影响SDH和呼吸活性。推测这类点突变在田间的适应性较强,极易导致田间病害的发生。结合AS-real-time PCR检测体系,预测含有H278R突变位点菌株的发展趋势,可指导药剂的合理使用及制定合理的田间管理策略。
本研究建立的AS-real-time PCR检测方法可以快速定量检测田间多主棒孢SdhB-H278R突变位点的频率,经验证AS-real-time PCR检测方法预期值和试验值的线性相关性为2=0.9998和2=0.9922。此方法可以检测到5%的SdhB-H278R抗性突变,灵敏度是普通PCR的10倍。相比高苇等[31-32]建立的检测黄瓜多主棒孢实时荧光定量PCR检测技术和普通PCR检测技术,此检测体系可以定量检测SdhB-H278R抗性突变位点以及抗性突变在田间频率的变化,克服了PCR-RFLP检测体系中变异系数大的难题[33-35]。与常规PCR检测方法相比较,该检测体系具有快速、无污染和灵敏度高的特点,并可根据检测结果指导科学用药以及降低用药成本和减少环境污染。
4 结论
建立了快速检测黄瓜多主棒孢菌SdhB-H278R突变位点的AS-real-time PCR技术体系,该体系自动化程度高,准确性强、灵敏度高,整个过程可在一个封闭的体系中进行,无污染、操作简单,可用于检测该突变位点在田间的频率。
[1] 李宝聚, 高苇, 石延霞, 谢学文. 多主棒孢和棒孢叶斑病的研究进展. 植物保护学报, 2012, 39(2): 171-176.LI B J, GAO W, SHI Y X, XIE X W. Progress in researches onleaf spot., 2012, 39(2): 171-176. (in chinese)
[2] 于淑晶, 王满意, 田芳, 赵卫光, 边强, 李宝聚. 黄瓜棒孢叶斑病的防治及抗药性研究进展. 农药, 2014, 53(1): 7-11.YU S J, WANG M Y, TIAN F, ZHAO W G, BIAN Q, LI B J. Progress in research on control of cucumberleaf spot and fungicide resistance., 2014, 53(1): 7-11. (in chinese)
[3] 李良孔, 袁善奎, 潘洪玉, 王岩. 琥珀酸脱氢酶抑制剂类(SDHIs)杀菌剂及其抗性研究进展. 农药, 2011, 50(3): 165-169.LI L K, YUAN S K, PAN H Y, WANG Y. Progress in research on SDHIs fungicides and its resistance., 2011, 50(3): 165-169. (in chinese)
[4] 颜范勇, 刘冬青, 司马利锋, 石恒, 胡欣. 新型烟酰胺类杀菌剂——啶酰菌胺. 农药, 2008, 47(2): 132-135.YAN F Y, LIU D Q, SIMA L F, SHI H, HU X. Boscalid, a novel carboxamide aka anilide class of fungicides., 2008, 47(2): 132-135. (in chinese)
[5] AVENOT H, Michailides T J. Resistance to boscalid fungicide inisolates from pistachio in California., 2007, 91(10): 1345-1350.
[6] YIN Y N, KIM Y K, XIAO C L. Molecular characterization of boscalid resistance in field isolates offrom apple., 2011, 101(8): 986-995.
[7] AVENOT H F, THOMAS A, GITAITIS R D, JR LANGSTON D B, STEVENSON K L. Molecular characterization of boscalid- and penthiopyrad-resistant isolates ofand assessment of their sensitivity to fluopyram., 2012, 68(4): 645-651.
[8] WANG Y, DUAN Y B, WANG J X, ZHOU M G. A new point mutation in the iron-sulfur subunit of succinate dehydrogenase confers resistance to boscalid in., 2015, 16(7): 653-661.
[9] MIYAMOTO T, ISHII H, SEKO T, KOBORI S, TOMITA Y. Occurrence ofisolates resistant to boscalid on cucumber in Ibaraki Prefecture, Japan., 2009, 58(6): 1144-1151.
[10] MIYAMOTO T, ISHII H, STAMMLER G, KOCH A, OGAWARA T, TOMITA Y, FOUNTAINE J M, USHIO S, SEKO T, KOBORI S. Distribution and molecular characterization ofisolates resistant to boscalid., 2010, 59(5): 873-881.
[11] FURUYA S, SUZUKI S, KOBAYASHI H, SAITO S, TAKAYANAGI T. Rapid method for detecting resistance to a QoI fungicide inpopulations., 2010, 65(8): 840-843.
[12] AOKI Y, FURUYA S, SUZUKI S. Method for rapid detection of thegene allele conferring resistance to mandipropamid, a carboxylic acid amide fungicide, inpopulations., 2011, 67(12): 1557-1561.
[13] 李红霞, 周明国. 用等位基因特异性寡核苷酸(ASO)-PCR快速检测抗多菌灵的油菜菌核病菌. 中国农业科学, 2004, 37(9): 1396-1399.LI H X, ZHOU M G. Rapid identification of carbendazim resistant strains ofusing allele-specific oligonucleotide (ASO)-PCR., 2004, 37(9): 1396-1399. (in chinese)
[14] MALLIK I, ARABIAT S, PASCHE J S, BOLTON M D, PATEL J S, GUDMESTAD N C. Molecular characterization and detection of mutations associated with resistance to succinate dehydrogenase- inhibiting fungicides in., 2014, 104(1): 40-49.
[15] WHEELER I, KENDALL S, BUTTERS J, HOLLOMON D. Detection of benzimidazole resistance inusing allele-specific oligonucleotide probes./, 1995, 25(1/2): 113-116.
[16] DE MICCOLIS ANGELINI R M, MASIELLO M, ROTOLO C, POLLASTRO S, FARETRA F. Molecular characterization and detection of resistance to succinate dehydrogenase inhibitor fungicides in()., 2014, 70(12): 1884-1893.
[17] LEHNER M S, JÚNIOR T J P, SILVA R A, VIEIRA R F, SCHNABEL G, MIZUBUTI E S G. Fungicide sensitivity of: A thorough assessment using discriminatory dose, EC50, high-resolution melting analysis, and description of new point mutation associated with thiophanate-methyl resistance., 2015, 99(11): 1537-1543.
[18] SAMARAS A, MADESIS P, KARAOGLANIDIS G S. Detection ofB gene mutations in SDHI-resistant isolates ofusing high resolution melting (HRM) analysis., 2016, 7: Article 1815.
[19] 聂燕钗, 王斌, 赵子琴, 周怀谷. 等位基因特异性PCR技术及其法医学应用. 法医学杂志, 2014, 30(4): 282-287.NIE Y C, WANG B, ZHAO Z Q, ZHOU H G. Allele-specific PCR and its application in forensic science., 2014, 30(4): 282-287. (in chinese)
[20] DIXON L J, SCHLUB R L, PERNEZNY K, DATNOFF L E. Host specialization and phylogenetic diversity of, 2009, 99(9): 1015-1027.
[21] 杨苗.我国蔬菜棒孢叶斑病病原菌多样性研究[D]. 北京: 中国农业科学院, 2013.YANG M. Diversity of pathogen of corynespora leaf spot on vegetables in china[D]. Beijing: Chinese Academy of Agricultural Sciences, 2013. (in chinese)
[22] 高苇, 李宝聚, 石延霞, 谢学文. 河北青县黄瓜棒孢叶斑病病原菌种群分化的研究. 华北农学报, 2011, 26(5): 9-15.GAO W, LI B J, SHI Y X, XIE X W. Population differentiation ofin qing county, Hebei province., 2011, 26(5): 9-15. (in chinese)
[23] 李长松, 张眉, 李林, 李凡, 齐军山, 徐作珽, 张博. 山东省黄瓜棒孢叶斑病(褐斑病)病原菌鉴定和防治. 中国蔬菜, 2009(18): 29-33.Li C S, ZHANG M, Li L, LI F, QI J S, XU Z T, ZHANG B. Identification of cucumber target leaf spot (brown spot) pathogen and its control., 2009(18): 29-33. (in chinese)
[24] KWON M K, KANG B R, CHO B H, KIM Y C. Occurrence of target leaf spot disease caused byon cucumber in Korea., 2010, 52(3): 424.
[25] 余玲, 刘慧平, 韩巨才, 张宝俊. 山西省灰霉菌对啶酰菌胺的敏感性测定. 山西农业大学学报(自然科学版), 2012, 32(3): 232-234.YU L, LIU H P, HAN J C, ZHANG B J. Sensitivity offrom Shanxi province to boscalid., 2012, 32(3): 232-234. (in chinese)
[26] VELOUKAS T, LEROCH M, HAHN M, KARAOGLANIDIS G S. Detection and molecular characterization of boscalid-resistantisolates from strawberry., 2011, 95(10): 1302-1307.
[27] 史晓晶. 番茄早疫病菌对啶酰菌胺的抗性检测及抗性机理初探[D]. 太谷: 山西农业大学, 2015. SHI X J. Study on sensitivity ofto boscalid and resistant mechanisms[D]. Taigu: Shanxi Agricultural University, 2015. (in chinese)
[28] YANG J H, BRANNEN P M, SCHNABEL G. Resistance into SDHI fungicides causes rare disease outbreak in peach orchards., 2015, 99(1): 65-70.
[29] HORSEFIELD R, YANKOVSKAYA V, SEXTON G, WHITTINGHAM W, SHIOMI K, OMURA S, BYRNE B, CECCHINI G, IWATA S. Structural and computational analysis of the quinone-binding site of complex II (succinate-ubiquinone oxidoreductase): a mechanism of electron transfer and proton conduction during ubiquinone reduction., 2006, 281(11): 7309-7316.
[30] LALÈVE A, GAMET S, WALKER A, DEBIEU D, TOQUIN V, FILLINGER S. Site-directed mutagenesis of the P225, N230 and H272 residues of succinate dehydrogenase subunit B fromhighlights different roles in enzyme activity and inhibitor binding., 2014, 16(7): 2253-2266.
[31] 高苇, 李宝聚, 王万立, 郝永娟, 石延霞. 土壤中黄瓜棒孢叶斑病病原菌实时荧光定量PCR检测技术研究. 华北农学报, 2014, 29(2): 71-74.GAO W, LI B J, WANG W L, HAO Y J, SHI Y X. Detection ofin soil with real-time quantitative PCR., 2014, 29(2): 71-74. (in chinese)
[32] 陈璐. 黄瓜细菌性角斑病菌和多主棒孢菌PCR检测技术的建立[D]. 北京: 中国农业科学院, 2014.CHEN L. PCR-based specific detection ofpv.and[D]. Beijing: Chinese Academy of Agricultural Sciences, 2014. (in chinese)
[33] MICHALECKA M, MALINOWSKI T, BRONIAREKNIEMIEC A, BIELENIN A. Real-time PCR assay with SNP-specific primers for the detection of a G143A mutation level infield populations., 2011, 159(7/8): 569-578.
[34] LUO Y, MA Z H, MICHAILIDES T J. Quantification of allele E198A in beta-tubulin conferring benzimidazole resistance inusing real-time PCR., 2007, 63(12): 1178-1184.
[35] KIANIANMOMENI A, SCHWARZ G, FELSENSTEIN F G, WENZEL G. Validation of a real-time PCR for the quantitative estimation of a G143A mutation in the cytochrome1gene of., 2010, 63(3): 219-224.
Establishment of AS-real-time PCR for Quantitatively Detecting the H278R Allele in theAssociated within Cucumber
SUN BingXue, SHI YanXia, ZHU FaDI, XIE XueWen, CHAI ALi, LI BaoJu
(Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences, Beijing 100081)
【Objective】The objective of this study is to establish a rapid and efficient allele specific real-time PCR (AS-real-time PCR) method for quantitative detection of the H278R mutation in theassociated within cucumber, and to verify the effect. 【Method】A total of 24 single conidial strains ofwere isolated from Daxing, Beijing.The method of mycelial growth inhibition was used to determine the EC50value to boscalid. Mycelium growth rate, spore outputs and pathogenicity of 4 susceptible (S) and 8 resistant (R) strains were measured. Primer pair Cc-SdhB-F/R was used to detect the sequence ofand the base change ofBased on the sequencing results inby Cc-SdhB-F/R primers, specific primer pair B-H278R-2F/2R14 and internal primers B-H278R-TY-F/R were designed. An AS-real-time PCR reaction system was established and optimized. The specificity, sensitivity and melt curve of the system were also evaluated.DNA and spore suspension containing different proportions of H278R mutation were detected with the optimized reaction system, respectively. 【Result】The sensitive frequency ofto boscalid was 66.67% in 24 strains and the EC50values ranged from 0.057 to 0.563 μg·mL-1. The resistance frequency ofto boscalid was 33.33% and the EC50values ranged from 5.395 to 11.710 μg·mL-1. Resistant and susceptible strains only significantly differed in terms of mycelium growth rate, and there was a significant negative correlation between the rate of mycelial growth and EC50values, but no significant difference in spore outputs and pathogenicity. Sequencing analysis showed that all the resistant strains carried SdhB-H278R mutation. The primers B-H278R-2F/2R14 were of great specificity, the specific fragment was amplified from the DNA of H278R strains, but not from the DNA of other fungal strains. The sensitivity of ordinary AS-PCR was 91 pg·μL-1, while that of AS-real-time PCR was 9.1 pg·μL-1, which was 10 times higher than that of ordinary AS-PCR. The standard curve established by AS-real-time PCR showed a fine linear relationship between ΔCT value and lg of H278R frequency, the correlation coefficient of the standard curve was 0.9857 and with high amplification efficiency (92.59%). The absorption peak of melting curve was single. The internal reference primers B-H278R-TY-F/R and the specific primer B-H278R-2F/2R14 had a single specific peak at 87.81℃and 91.62℃, respectively. The result of mixtures DNA and mixtures spore suspension to verify the standard curve showed a fine linear relationship between expected percentage and detected percentage (2=0.9998 and2=0.9922). As the proportions of H278R mutation inDNA and spore suspension decreased, the accuracy of the system gradually increased, and the detection limit was 5%. 【Conclusion】An efficient, quantitative, and rapid AS-real-time PCR detection system was established for the detection of SdhB-H278R mutation site, which can provide a theoretical basis for the SDHIs resistance management.
cucumber;; AS-real-time PCR; resistance;; boscalid
2018-06-22;
2018-09-15
国家重点研发计划(2018YFD0201300)、中国农业科学院科技创新工程(CAAS-ASTIP-IVFCAAS)、农业部园艺作物生物学与种质创制重点实验室开放课题(IVF2017)
孙炳学,E-mail:sunbingxuechina@163.com。石延霞,E-mail:shiyanxia@caas.cn。孙炳学和石延霞为同等贡献作者。
李宝聚,E-mail:libaoju@caas.cn
10.3864/j.issn.0578-1752.2018.24.006
(责任编辑 岳梅)
