胆汁酸盐对细粒棘球绦虫分化和发育的调控※
2018-12-14张文宝
郭 刚,张文宝
(中亚高发病成因与防治国家重点实验室,临床医学研究院,新疆医科大学第一附属医院,新疆 乌鲁木齐 830054)
胆汁酸盐可以决定细粒棘球绦虫原头蚴的双向发育,即发育为无性繁殖的包囊或发育为有性繁殖的成虫,使得细粒棘球绦虫成为研究胆汁酸生物学功能及胆汁酸调控寄生虫分化、发育的生物学模型[1,2]。本文从胆汁酸盐与细粒棘球绦虫双向发育的关系,胆汁酸信号通路方面阐述细粒棘球绦虫可以作为研究胆汁酸调控分化和发育模型生物的可能性,为今后深入研究细粒棘球绦虫的分化和发育,筛选成虫发育基因提供思路。同时,也为研究胆汁酸新的生物学功能以及分析胆盐相关的生物基因或蛋白提供了一个很好的模式平台。现对胆汁酸盐对细粒棘球绦虫分化和发育的调控作用在此作一综述。
1 胆汁酸盐
胆汁酸的分子结构有两性,一端为烷基,具有亲油性,可以和油脂类结合;另一端为羟基和羧基,具有亲水性。胆汁酸常以钠盐或钾盐形式存在,形成胆汁酸盐。牛磺胆酸盐(TC,taurocholate)和甘氨胆酸盐(GC,glycocholate)占胆盐成分的80%。另外还有甘氨酸鹅脱氧胆盐(GCDC,glycochenodeoxycholate)、甘氨脱氧胆酸盐(GDC,glycodeoxycholate)、牛磺酸鹅脱氧胆盐(TCDC,taurochenodeoxycho- late,)和牛磺酸脱氧胆酸(TDC,taurodeoxycholate,)。
细粒棘球绦虫终末宿主的种类不同可能导致分泌胆汁组分的不同。例如人胆汁组成有与牛磺酸结合的胆酸、鹅脱氧胆酸、脱氧胆酸,与甘氨酸结合的石胆酸、熊脱氧胆酸,犬胆汁组成则是与牛磺酸结合的胆酸、鹅脱氧胆酸和脱氧胆酸,而狐狸胆汁只有与牛磺酸结合的胆酸与脱氧胆酸[3]。
2 胆汁酸盐与细粒棘球绦虫的双向发育
2.1 宿主体内胆汁酸盐对细粒棘球绦虫的作用
细粒棘球绦虫在幼虫阶段具有双向发育的特点,即可以发育为成虫,也可以发育为包囊幼虫[2]。犬摄入原头蚴可以在肠道里发育为成虫。另一方面,如果一个原头蚴在中间宿主比如人的器官中破裂,释放出的每一个原头蚴能够发育为一个新的包囊幼虫。
细粒棘球绦虫发育,无论是在终末宿主肠道内原头蚴头节的外翻,成虫的生长,还是在中间宿主肠道内虫卵的孵化和六钩蚴的激活都需要有胆酸盐的参与。但没有资料表明寄生虫可以合成胆酸,像许多寄生蠕虫一样,棘球绦虫无论是寄生在终末宿主小肠内的成虫,还是寄生在中间宿主肝脏的幼虫,都生活在含有胆汁环境的脏器内,提示宿主体内的胆酸提供了寄生虫生理发育所需要的条件。
原头蚴经胃液和十二指肠液短暂的作用后,定居在犬的小肠内进入成虫发育阶段。成虫自身发育分化的因素包括原头蚴的成熟状态和原头蚴的外翻[3]。原头蚴被犬吞服后进入胃,首先要经过一个酸性环境支持下的胃蛋白酶的消化和激活作用,未发育成熟或老化的及死亡的原头蚴被胃蛋白酶消化,成熟和健康的原头蚴因为具有很强的耐胃酸和抗胃蛋白酶及胰蛋白酶的作用而得以生存、生长。原头蚴外翻后,其外形像一个幼小的成虫,具有头节和颈节的结构,并且顶突、吸盘和头钩发育基本完全[4]。
2.2 体外胆汁酸盐的双向调节作用
人们对成虫早期,即前两周发育观察的大部分数据主要来源于体外培养。体外试验证明,在培养系统中加入犬胆汁或牛磺胆酸钠,原头蚴便向成虫发育,没有胆汁或牛磺胆酸钠,原头蚴向包虫囊发展。胆盐为成虫发育的必须因素,无胆盐则发育至包囊。在模拟成虫在犬体内发育条件的基础上,Smyth在上世纪60~80年代对细粒棘球绦虫的体外和体内发育做了系统的研究[2],得出培养成虫需具备两个条件的结论:(1)原头蚴需要胃肠液的处理,即先经过胃蛋白酶消化,再用胰蛋白酶和胆汁溶液处理。研究结果说明两个生理学现象:首先,原头蚴的成虫发育需要一个激活和外翻的过程;其次是原头蚴具有很强的抗胃蛋白酶和胰蛋白酶的功能;(2)成虫的培养一定要在双相培养基系统中进行,即下层有用新生牛血清制成的血清凝胶层,上层为含有胆汁的普通细胞培养基。该结果说明,成虫的生长需要类宿主组织的固体支持物,胆汁是成虫发育和分化必须的物质。近期我们的研究显示前期对原头蚴进行适当刺激促其外翻的基础上,在含胆汁培养基中培养,不需要双相培养系统,也可以促进其向成虫发育。
3 胆汁酸信号通路
胆酸盐对棘球绦虫的成虫发育起到至关重要的作用,但其机制还不清楚。从一些生理现象,比如原头蚴和成虫体内含有大量的脂肪,可以推断棘球绦虫脂肪的吸收和代谢与小肠的胆汁环境有关。基因组测序发现棘球绦虫有与多种胆酸结合和转移的蛋白基因[5],说明胆汁酸参与的虫体脂肪代谢是存在的。胆盐的浓度和组成对原头蚴的外翻、成虫的定位和发育是必要的因素,但如果胆盐过多,无论对虫体或宿主都会产生毒性副作用,并有可能产生裂解作用。
蜕皮激素或其他类固醇激素可以在寄生线虫通过核激素受体调节蜕皮[6,7],同样来源于宿主的胆汁酸也可能会以一个类似的方式在细粒棘球绦虫发育中起重要作用[8,9]。目前已知有两大类胆汁酸信号受体,分别为G蛋白偶联受体(GPCR,G Protein-Coupled Receptor,也称TGR5)和核激素受体[包括法尼酯X受体(FXR,Farnesoid X Receptor)]。细粒棘球蚴可以表达胆汁酸受体和转运胆汁酸的转运蛋白,并刺激相关信号通路(图1)。
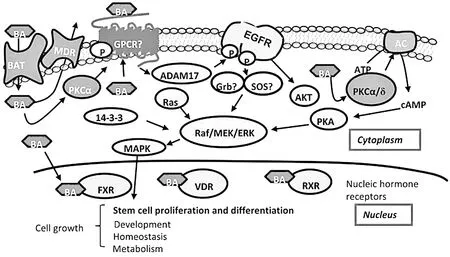
胆汁酸是由胆汁酸转运蛋白(BAT)转运到细胞内.当胆汁酸结合于G蛋白-偶联受体(GPCR),如G-蛋白偶联型胆汁酸受体(GPBAR).胆汁酸的结合引起GPBAR构象的变化,这种变化使之与鸟嘌呤核苷酸交换因子(GEF)活化,然后GPCR可以通过结合来源于GTP交换的GDP激活G-蛋白,并诱导产生细胞内cAMP和激活MAP激酶(MAPK)信号通路.胆汁酸也可以刺激G蛋白活化的腺苷酸环化酶(AC),以催化ATP转化为cAMP和焦磷酸,导致加快生产cAMP.胆汁酸也可以直接通过结合到细胞外区域,和刺激信号传导途径,激活基质金属蛋白酶ADAM17和表皮生长因子受体(EGFR),然后激活不同的激酶,包括MAP激酶和蛋白激酶AKT.
图1胆汁酸信号通路概述图
在细粒棘球蚴基因组或转录组中没有发现TGR5样受体,但确定了4个基因作为胆汁酸信号的核受体候选基因(EG_00119、EG_00780、EG_04405和EG_08428),它们编码的蛋白与FXR和维生素D受体(VDR)有超过20%的氨基酸同源,并包含一个DNA结合结构域和一个配体结合结构域[5]。基因组测序还发现了5个胆酸钠共同转运基因和7个多药耐药蛋白[5,10],以及其他一些与胆酸代谢相关的基因如固醇调节元件结合蛋白1和胆酸的β-葡萄糖苷酶相关基因等[5]。宿主的外源性胆酸有可能通过胆酸钠共同转运蛋白转运到细胞内,并与核激素受体结合,在细粒棘球蚴的发育中发挥作用。
目前并不排除在细粒棘球蚴中存在G蛋白偶联受体(GPCR)及未知胆酸受体的可能性。FXR类核受体在物种间比较保守,在细粒棘球蚴胆酸信号传输过程中可能发挥作用[5]。同时,研究发现核受体FXR/VDR相对于膜受体TGR5(EC50 300~600nM)而言,它们的生理配体(EC50 ~10μM)值较低,表现为与胆汁酸结合不敏感,需要高浓度的胆汁酸,细粒棘球蚴胆汁酸信号通路中很有可能存在类似机制。这可能解释了犬肠道中胆酸浓度高,FXR/VDR可以结合胆汁酸,激活信号通路,使原头蚴向成虫发育,而在较低的胆汁酸环境中,原头蚴向包囊方向发育。
4 结语
宿主胆汁酸可以对细粒棘球绦虫的分化、发育发挥调控作用,然而对其发挥调控作用的组分和机理尚不十分明确,今后需借助高通量转录组学、蛋白质组学等技术在此领域深入开展研究。
