丙型副伤寒沙门菌lpfC基因缺失株的构建及其生物学特性分析
2018-11-23杨占峰张晶晶吴敏娜何群力
杨占峰,张晶晶,吴敏娜,何群力,
(1.郑州工业应用技术学院医学院,郑州 451100;2.新乡医学院基础医学院,新乡 453000)
沙门菌(Salmonella)是引起沙门菌病的病原体,属于肠杆菌科、沙门菌属,目前已有2600个以上的血清型被确定,主要寄居于人类和动物的肠道,可以通过侵袭肠道上皮细胞进入血液循环系统,引起严重的系统感染,出现败血症和菌血症,并可引起机体腹泻、发热等症状,严重时可导致死亡[1-3]。丙型副伤寒沙门菌(Salmonella paratyphi C)是高度适应于人类的专嗜性沙门菌,可引起严重的系统感染,导致副伤寒[4-5]。有数据显示,近些年我国副伤寒沙门菌的流行有所增加,其中数次疫情与丙型副伤寒沙门菌相关[6-9]。
lpfC基因属于沙门菌中长极性菌毛(long polar fimbriae,LPF)操纵子家族成员之一,与LPF菌毛外膜蛋白的编码相关[10]。有研究显示,在肠炎沙门菌感染巨噬细胞的过程中,lpfC基因在胞内的转录水平显著升高[11]。本研究拟通过同源重组的方法构建丙型副伤寒沙门菌BF02的lpfC基因缺失株,并构建其互补株,对其基本生物学特性进行测定。
1 材料与方法
1.1 菌株、质粒、细胞及实验动物丙型副伤寒沙门菌BF02、BF02/pkD46(Ampr)和pKD3(Cmr)、pCP20(Ampr,Cmr)、互补质粒pSTV28-lpfC(Cmr)等均由本实验室保存;人结肠癌细胞系Caco-2由本实验室保存;8周龄雌性BALB/c小鼠购自北京维通利华实验动物技术有限公司。
1.2 主要试剂2×PCP Mix、Agarose Gel DNA Extraction Kit和DNA Marker购自天根生化科技(北京)有限公司;细菌微量生化发酵管购自杭州天和微生物试剂有限公司;DMEM培养基、胎牛血清购自Gibco公司。
1.3 引物设计根据GenBank中公布的丙型副伤寒沙门菌基因组序列,利用软件Primer Premier 5.0 设计lpfC基因缺失引物、缺失鉴定引物及互补引物(表1),由南京金斯瑞生物科技有限公司合成。
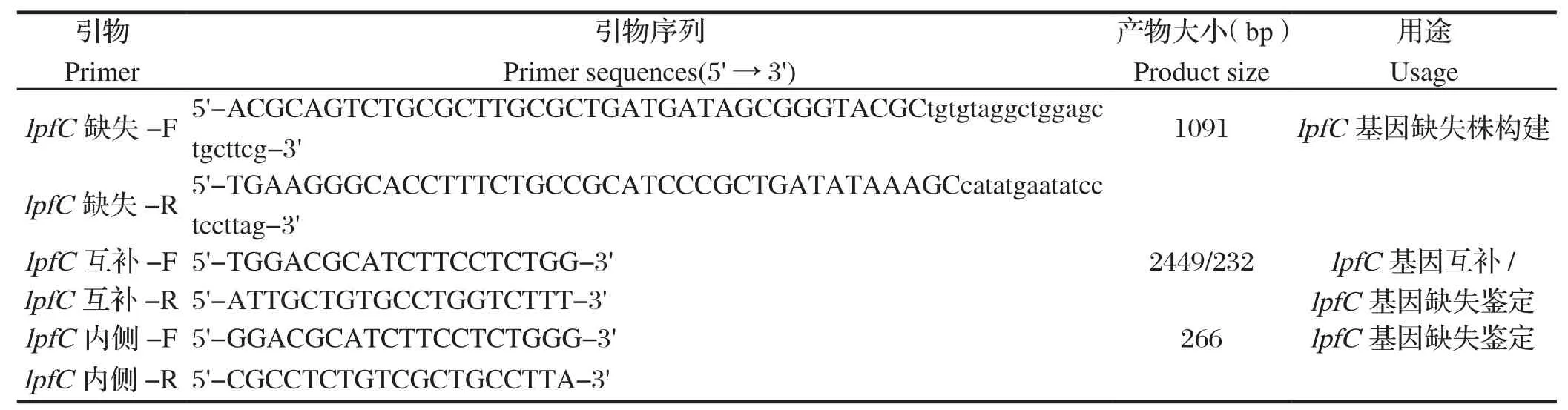
表1 本研究中所用引物序列Table 1 Primers used in this study
1.4 缺失株及互补株构建以pKD3质粒为模板,用引物lpfC缺失-F/lpfC缺失-R扩增lpfC基因上下游同源臂的氯霉素抗性片段。根据文献[12]中的方法,利用Red同源重组技术,将含有lpfC基因上下游同源臂的氯霉素抗性片段电转化入表达Red同源重组酶的BF02感受态细胞中,涂布氯霉素抗性平板。挑取单克隆,采用PCR及测序鉴定lpfC基因缺失株。此外,将本实验室保存的互补质粒pSTV28-lpfC电转化入缺失株BF02ΔlpfC,构建互补株BF02CΔlpfC。
1.5 缺失株生长特性测定分别挑取丙型副伤寒沙门菌野生株BF02、缺失株BF02ΔlpfC和互补株BF02CΔlpfC接种于LB液体培养基中,培养过夜,次日分别转接入10 mL的LB 液体培养基中(调节OD600=0.05)。37℃、220 r/min 摇振培养,在培养后的0.5、1、1.5、2、2.5、3、3.5、4 h测定培养物的OD600值,并绘制细菌的生长曲线。
1.6 缺失株生化特性测定分别挑野生株BF02、缺失株BF02ΔlpfC和互补株BF02CΔlpfC,过夜培养后挑取新鲜的单菌落转接于葡萄糖、乳糖、蔗糖、麦芽糖、甘露醇、尿素、赖氨酸脱羧酶、鸟氨酸脱羧酶和山梨醇等生化鉴定管,37℃过夜,观察生化鉴定结果。
1.7 细胞黏附与侵袭试验将新鲜培养菌液用DMEM洗涤后,按MOI=100感染人结肠癌细胞系Caco-2,在37℃、5%CO2细胞培养箱中感染2 h,同时设置DMEM阴性对照组。然后以PBS洗涤3次,用0.1%的Triton-X 100裂解细胞并倍比稀释,涂平板,进行菌落计数,统计黏附试验结果,每个菌株的黏附情况用相对于野生株的变化倍数表示。细胞侵袭试验与细胞黏附试验的感染方式相同,感染2 h后以无菌PBS洗涤3次,用含有庆大霉素的DMEM作用1 h杀死胞外细菌。然后用0.1%的Triton-X 100裂解细胞并倍比稀释,涂平板,进行菌落计数,统计侵袭试验结果,每个菌株的侵袭情况用相对于野生株的变化倍数表示。
1.8 毒力试验将野生株BF02、缺失株BF02ΔlpfC和互补株BF02CΔlpfC单菌落分别接种于液体LB培养基中,37℃、180 r/min培养10~12 h,以PBS洗涤细菌3次并重悬,调整细菌浓度。攻毒前先给小鼠断水断食6 h,然后将3种菌液分别以2×108、4× 107、8×106、1.6×106、3.2×105CFU 的剂量腹腔注射小鼠,每个剂量组5只,对照组注射PBS。连续观察2周并统计小鼠死亡情况,计算3个菌株的半数致死量(lethal dose,LD50),评价其对BALB/c 小鼠的毒力。
2 结果
2.1 基因缺失株、互补株的构建与鉴定挑取疑似基因缺失株进行PCR鉴定,结果显示缺失株BF02ΔlpfC内侧鉴定引物lpfC内侧-F/lpfC内侧-R无法扩增出lpfC基因(图1泳道4);外侧鉴定引物lpfC互补-F/lpfC互补-R可以扩增出替换lpfC基因,大小为232 bp的氯霉素抗性片段(图1)。PCR产物测序结果表明缺失株BF02ΔlpfC构建成功。构建互补株时,内侧鉴定引物lpfC内侧-F/lpfC内侧-R和外侧鉴定引物lpfC互补-F/lpfC互补-R分别扩增出了大小为266 bp和232 bp的条带(图1),与预期结果一致,表明互补株BF02CΔlpfC构建成功。
2.2 生长曲线的测定生长曲线结果显示,野生株BF02、缺失株BF02ΔlpfC和互补株BF02CΔlpfC的生长速度无显著差异,表明lpfC基因不影响丙型副伤寒沙门菌BF02的生长速度(图2)。
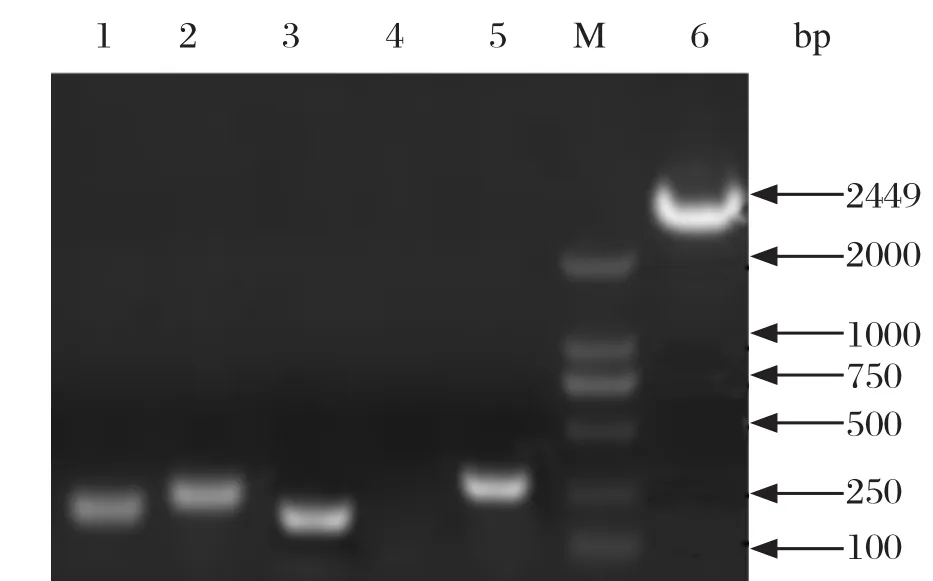
图1 缺失株BF02ΔlpfC及互补株BF02CΔlpfC的鉴定Fig.1 Identi fication of BF02ΔlpfC and BF02CΔlpfC with primers
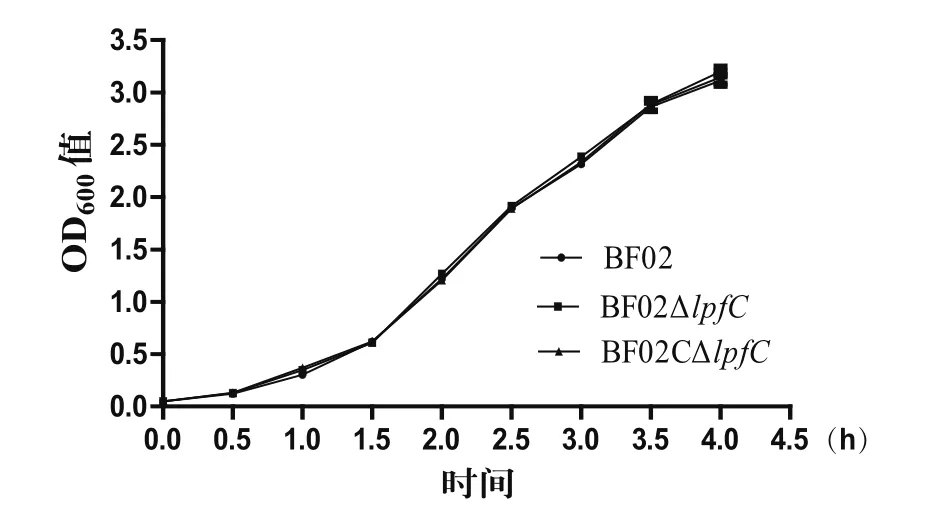
图2 野生株BF02、缺失株BF02ΔlpfC和互补株BF02CΔlpfC的生长曲线Fig.2 Growth curves of wild-type strain BF02, mutant strain BF02ΔlpfC and complementary strain BF02CΔlpfC
2.3 生化特性的测定生化鉴定结果显示,丙型副伤寒沙门菌野生株BF02、缺失株BF02ΔlpfC和互补株BF02CΔlpfC的生化特性一致,均能发酵葡萄糖、麦芽糖、甘露醇、赖氨酸脱羧酶、鸟氨酸脱羧酶和山梨醇,而不能发酵乳糖、蔗糖和尿素等,表明lpfC基因的缺失不影响丙型副伤寒沙门菌的生化特性。
2.4 黏附侵袭试验黏附侵袭结果显示,缺失株BF02ΔlpfC对Caco-2细胞的黏附侵袭能力均低于野生株BF02和互补株BF02CΔlpfC,差异具有显著统计学意义(P< 0.05)(图3),表明lpfC基因的缺失显著影响丙型副伤寒沙门菌对Caco-2细胞的黏附侵袭能力。
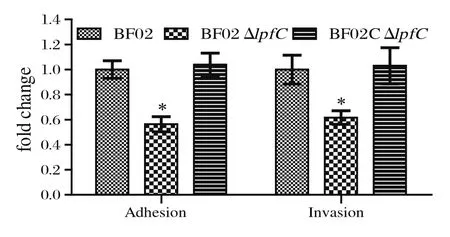
图3 野生株BF02、缺失株BF02ΔlpfC和互补株BF02CΔlpfC对Caco-2细胞的黏附侵袭能力Fig.3 Adhesion and invasion capacity of wild-type strain BF02, mutant strain BF02ΔlpfC and complementary strain BF02CΔlpfC to Caco-2 cells
2.5 毒力试验根据bliss法计算,丙型副伤寒沙门菌BF02的LD50为3.50×106CFU,缺失株BF02ΔlpfC的LD50为1.75×107CFU,互补株BF02CΔlpfC的LD50为5.27×106CFU,缺失株BF02ΔlpfC的LD50是其亲本野生株BF02的5倍,互补株BF02CΔlpfC的毒力部分恢复到野生株的水平。
3 讨论
沙门菌菌毛结构复杂,参与沙门菌对宿主细胞的粘附,与沙门菌的致病性密切相关。lpfABCDE是长极性菌毛操纵子,有研究显示,LpfC蛋白参与LpfA菌毛亚单位的输出和组装,并且与沙门菌侵入派尔氏结及在其内定植相关[10]。目前,关于丙型副伤寒沙门菌致病机理及其防控的研究还很少。
本研究采用Red同源重组系统构建丙型副伤寒沙门菌BF02的lpfC基因缺失株,可获得无耐药性基因缺失株,不会引起生物安全隐患。Red同源重组系统也不会对缺失基因的下游基因产生极性效应,因此,该技术被广泛用于大肠杆菌、沙门菌等细菌的基因缺失株构建[12]。作为与沙门菌LPF菌毛合成相关的基因,生物学特性分析结果表明,lpfC基因的缺失不影响丙型副伤寒沙门菌的生长特性和生化特性,说明其对丙型副伤寒沙门菌的生长代谢没有影响;在肠炎沙门菌中lpfC基因缺失后,这些特性与我们的研究结果一致[13]。黏附侵袭试验结果显示,lpfC基因的缺失显著降低了丙型副伤寒沙门菌对Caco-2细胞黏附侵袭能力,表明lpfC基因在丙型副伤寒沙门菌黏附及侵袭细胞过程中发挥重要作用。有研究显示,lpfC基因的缺失显著降低肠炎沙门菌对IPEC-J2的黏附能力,但对IPEC-J2的侵袭能力无明显影响[14],这与我们的研究结果不完全一致,具体原因有待于进一步深入研究。毒力试验结果显示,lpfC基因缺失株的LD50是其亲本野生株的5倍,说明lpfC基因的缺失会使丙型副伤寒沙门菌的毒力降低。肠炎沙门菌和鼠伤寒沙门菌在缺失了lpfC基因后,缺失株的LD50分别是野生株的2倍和3倍[13,15],说明lpfC基因的缺失同样会引起肠炎沙门菌和鼠伤寒沙门菌的毒力降低,这与我们的研究结果一致,也说明lpfC基因与沙门菌毒力相关。
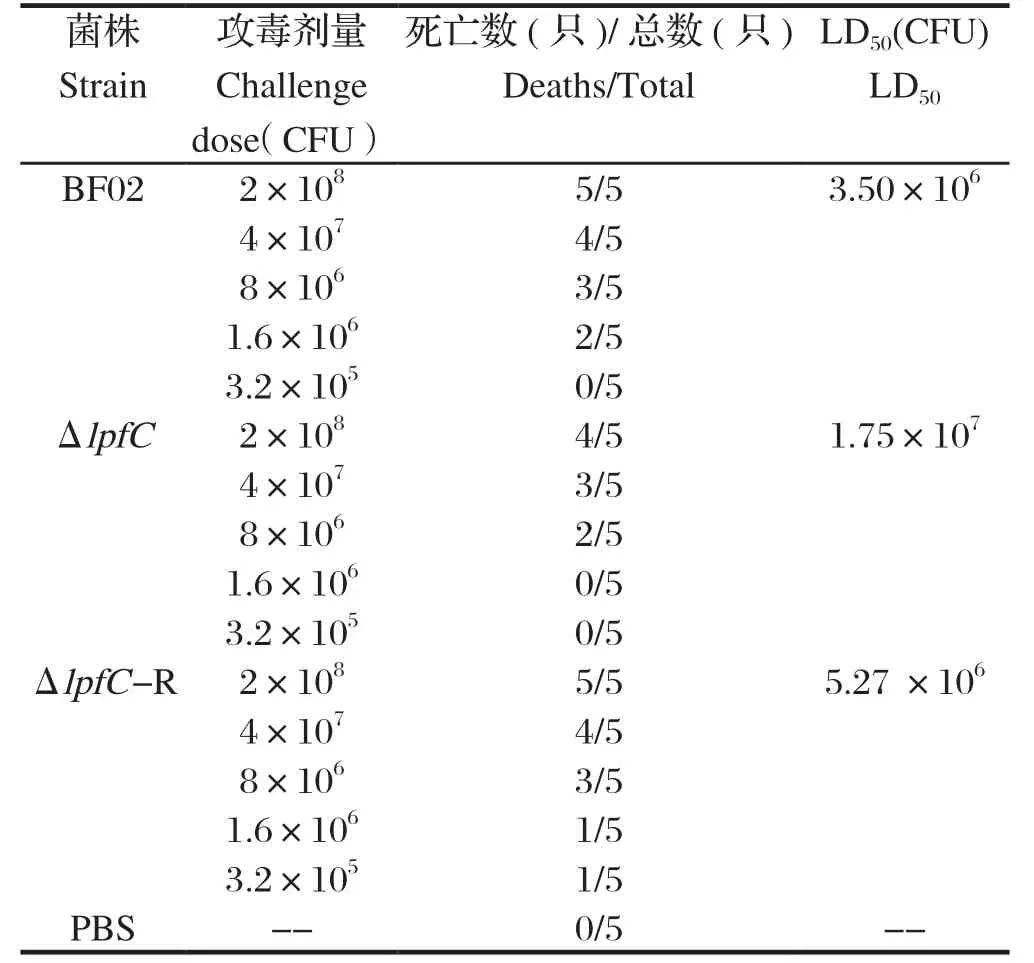
表2 野生株BF02、缺失株BF02ΔlpfC和互补株BF02CΔlpfC的LD50测定Table 2 LD50 of BF02, BF02ΔlpfC and BF02CΔlpfC
本研究成功构建了丙型副伤寒沙门菌的lpfC基因缺失株,并分析了lpfC基因对丙型副伤寒沙门菌生物学特性和致病性的影响,为进一步研究lpfC基因的功能及其在丙型副伤寒沙门菌致病过程中所起的作用奠定了基础。
