西藏白菜型油菜油体特征与营养品质及油体提取物的SDS-PAGE分析
2018-10-24侯维海王建林
侯维海,王建林,旦 巴,胡 单
(西藏农牧学院 高原作物分子育种实验室,植物科学学院,西藏 林芝 860000)
植物种子内积累的三酰甘油酯(TAG)是种子重要的储藏物质之一,为种子萌发、幼苗自养提供营养和能量来源。三酰甘油酯一般由离散的球形液滴细胞器,即油体或脂肪体所包裹,在种子成熟阶段,其以小而聚集的形式广泛分布于胚乳、子叶或胚轴等组织[1-2]。前人研究表明,TAG合成过程中,油体在内质网特化位点合成并吸附在内质网上,在复活酶的作用下填充中性脂质并逐渐膨大至最佳大小,然后释放进入细胞质[3],最终小的油体彼此融合或合并形成更大的油体[4]。油体是植物细胞中最小的细胞器,不同植物中油体平均粒径为0.2~2.5 μm,其大小、数量等可能受油体C代谢、脂肪酸合成、TAG合成等多个因素的调控[4-6]。油体由单层磷脂膜包被的液态TAG基质和嵌入表面的油体结合蛋白组成[2,7-10],这些油体结合蛋白对维持油体的稳定、参与油体的形态建成和生物学功能的发挥、油脂代谢和利用等具有重要影响[1,11-12]。例如,油体在干燥种子细胞内或体外经离心分离的上浮液中均不发生融合或聚合,甚至经过长时间的贮存也能够保持稳定[11]。此外,不同植物种子其油体的大小、数量等表型均存在差异,这可能与油体C代谢、脂肪酸合成、TAG合成及细胞分裂等有关,也可能受营养和环境等多个因素的调控[5-7,13]。
植物种子油体的分离纯化液中常混有0.6%~3.0%的蛋白质[3],这些蛋白质已从多种植物种子油体中得到鉴定,被认为是油体结合蛋白并将其分为3类[14]。其中第1类是油体蛋白,其含量占油体结合蛋白的75%~80%,几乎覆盖在整个油体表面,为主要油体结合蛋白,由分子质量为15~26 ku的小分子量碱性蛋白组成[15-16]。目前,在多种植物中发现油体蛋白存在2个或2个以上的异构体,通常将其简单地分为高或低分子量类型[8-9,13,17-18]。第2类是分子质量约为27 ku的油体钙蛋白,包含1个Ca2+结合EF手型的N-末端亲水结构域和1个C-末端亲水结构域[19]。第3类是分子质量约为37 ku的油体固醇蛋白,包含1个N-末端脂质锚定位点和固醇结合脱氢酶结构域[20],涉及油体的形成和降解[21]。另外,还有脂质液滴相关蛋白(LDAP)[22]、主要脂质液滴蛋白(MLDP)[23]和分子质量为39 ku的油体结合蛋白也相继被鉴定。因此,油体结合蛋白种类较多。此外,油体提取液中易混有贮藏蛋白。
当前,关于种子含油量与油体、油体结合蛋白之间的关系研究一直受到人们的关注。研究表明,油体结合蛋白直接涉及油体磷脂膜的形成、油体融合、防止种子失水引起的油体聚集、保持油体边界清晰和单个小油体的稳定等[8,17],并具有调控油体大小和功能的作用[4,10,23-24]。与低油品种相比,高油品种种子油体有较高的截面积[9]、更大的油体结构和更高的TAG/油体蛋白比值[10]及高油体蛋白表达水平[13]。有研究表明,甘蓝型油菜种子油体的总面积与含油量呈正相关,油体大小与油体蛋白呈负相关[13,25-26]。有研究从分子水平上对油体数量、TAG含量和油体蛋白的关系进行了解析,如Liu等[27]研究表明,在水稻中过表达大豆油体蛋白基因,可导致油体数量增加,体积减小。在拟南芥中过表达BnGRF2a油体蛋白,种子含油量显著增加[28]。抑制18 ku油体蛋白基因导致种子形成了更大的油体,而TAG含量相应下降[2,15]。在水稻中分别敲除胚乳16和18 ku油体蛋白基因,种子TAG含量分别下降约60%和80%[29]。抑制油体蛋白不仅会引起种子胚细胞的畸变,也会导致非正常的大油体形成[2]。利用RNAi抑制大豆油体蛋白,会导致油体/内质网复合体-微油体的产生,表明油体蛋白具有表面活性剂的功能。此外,油体蛋白与脂肪酸也有一定的关系。利用RNAi可以抑制拟南芥18 ku油体蛋白基因的表达,改变脂类和蛋白质的积累,其中TAGs的C20∶1增加而C18∶1有所减少[2]。 以上结果表明,油体和油体蛋白与种子含油量和脂肪酸含量紧密相关。
目前,国内外对于油体的研究常采用透射电镜技术和激光共聚焦技术等,其不足之处在于试验设备要求高且工作效率低[28,30]。西藏高原独特的气候和环境,孕育出了丰富且独具高原特征的油菜种质资源[25,31],表现为遗传变异丰富和含油量高等特点,这些优异特性与油体和油体结合蛋白表型是否存在联系目前尚无系统研究和明确的结论。鉴于此,本研究采用光学生物显微镜对西藏白菜型油菜种子油体、油体蛋白表型进行拍照,利用Imagepro-plus精确提取油体和油体蛋白表型数据,并结合含油量和总蛋白、脂肪酸含量进行分析,同时利用SDS-PAGE电泳检测油体提取物中蛋白亚基的组成,以期明确西藏高原地理生态条件下白菜型油菜种子油体表型、含油量、蛋白质和脂肪酸含量间的响应关系,并分析材料间的亲缘关系,为西藏白菜型油菜高油新品种的选育奠定基础。
1 材料与方法
1.1 材 料
供试材料西藏白菜型油菜农家种、野生近缘种共计36份(编号为1~36),由西藏农牧学院农学教研室于2015年从西藏全境内72个县的自然村搜集获得,于2016年3月中旬种植于西藏农牧学院试验田,小区面积2 m×2 m,每份材料3次重复,常规田间管理,于材料成熟后采集角果,单独脱粒进行后续试验。
1.2 方 法
1.2.1 测定项目及方法 种子含油量按照GB/T 14488.1-2008的方法测定;种子蛋白质含量按照GB/T 14489.2-2008的方法测定;种子脂肪酸(芥酸 C22∶1、棕榈酸C16∶0、硬脂肪酸C18∶0、油酸C18∶1、亚油酸C18∶2、亚麻酸C18∶3、C20烯酸)含量按照GB/T 22223-2008的方法测定。
1.2.2 油体的提取 参考Tzen等[7]的油体提取方法并作适当改动。取0.5 g干种子置于预冷的研钵内,加入2.5 mL研磨液(pH 7.5的10 mmol/L磷酸钠缓冲液,0.6 mol/L蔗糖),迅速充分研磨成匀浆,过3层纱布,收集滤液到15 mL离心管,加入2.5 mL漂浮液Ⅰ(含pH 7.5的10 mmol/L磷酸钠缓冲液,0.4 mol/L蔗糖),10 000×g离心20 min;挑取油体至15 mL新离心管,加入5 mL去污清洗液(含0.2 mol/L蔗糖,体积分数0.1% Tween-20,pH 7.5的5 mmol/L磷酸钠缓冲液)悬浮油体,再加入5 mL 10 mmol/L磷酸钠缓冲液(pH 7.5),10 000×g离心20 min;然后挑取油体至15 mL新离心管,加入5 mL离子洗脱液(含0.6 mol/L蔗糖,2 mol/L氯化钠,pH 7.5的10 mmol/L磷酸钠缓冲液)悬浮油体,再加入5 mL漂浮液Ⅱ(含0.25 mol/L蔗糖,2 mol/L氯化钠,pH 7.5的10 mmol/L磷酸钠缓冲液)10 000×g离心20 min;挑取油体至1.5 mL离心管,离心后于4 ℃保存备用。
1.2.3 油体观察 挑取适量油体均匀涂抹于载玻片上,滴60 μL体积分数20%的甘油,封片,用酒精灯烤至微干,在Leica DM5000生物显微镜1 000倍视野下观察并拍照,每个样品3个视野,3次重复。
1.2.4 油体大小的统计 利用Imagepro plus软件对油体显微照片进行处理,并随机选取300个油体和25个油体蛋白进行粒径测量,3次重复。
1.2.5 油体蛋白的SDS-PAGE电泳 挑取0.01 g油体放入1.5 mL试管中,加入50 μL研磨液(含pH 7.5的10 mmol/L磷酸钠缓冲液,0.6 mol/L蔗糖)稀释混匀,吸取40 μL混合液加入12 μL 5×SDS-PAGE loading buffer,涡旋振荡混匀,100 ℃温浴5 min,3 000×g离心20 s,放入4 ℃冰箱预冷。取上清液25 μL进行SDS-PAGE电泳,浓缩胶5%,分离胶10%,恒流15 mA,待指示剂泳动至胶底停止电泳。采用考马斯亮蓝R250染色后,酒精和乙酸溶液脱色,拍照。
1.3 数据统计与分析
利用Imagepro plus软件对1 000×光学显微照片进行细胞观察,并统计其粒径数据,通过SPSS20软件进行数据统计与分析。利用Quantity-one软件自动读取蛋白谱带,人工辅助修正,按照同一位置,有蛋白电泳条带赋值为1,无蛋白条带赋值为0,生成“0,1”矩阵。通过NTSYS-pc 2.1软件对已生成的“0,1”矩阵进行非加权类平均法聚类,计算遗传距离和遗传相似系数,通过Microsoft Excel计算出现频率。
2 结果与分析
2.1 白菜型油菜种子含油量及蛋白质、主要脂肪酸含量分析
含油量分析结果(表1)表明,36份供试材料含油量差异较大。根据含油量高低,将36 份材料分为高油组(含油量≥138 mg/g,HO)、中油组(110 mg/g≤含油量<138 mg/g,MO)、 低油组(含油量<110 mg/g,LO),其平均含油量分别为144.26,132.77和88.29 mg/g,含油量最高和最低的材料分别为14号和23号,含油量分别为151.37和57.20 mg/g。HO、MO和LO组平均蛋白质含量分别为44.70, 51.09和66.91 mg/g,其中23号材料的蛋白质含量最高,30号材料最低,其蛋白质含量分别为73.87和38.68 mg/g。分析含油量和蛋白质含量的关系可以发现,随着含油量的逐渐增加,蛋白质含量呈下降趋势,两者存在负相关关系。
脂肪酸含量分析结果(表1)显示,芥酸、油酸、亚油酸和烯酸是脂肪酸的主要组成部分,而棕榈酸和硬脂肪酸含量相对较低。根据芥酸(C22∶1)含量的大小,可将供试材料分为高芥酸组(芥酸含量≥110 mg/g)、中芥酸组(55 mg/g≤芥酸含量<110 mg/g)、低芥酸组(芥酸含量<55 mg/g)3种类型,其中芥酸最高值为132.33 mg/g,最低值为27.39 mg/g。油酸(C18∶1)、亚油酸(18∶2)含量表现出与芥酸相反的结果。高芥酸组材料的油酸含量为3.22~42.60 mg/g,亚油酸含量为31.22~47.00 mg/g;中芥酸组材料的油酸含量为21.44~101.51 mg/g,亚油酸含量为36.40~49.32 mg/g;低芥酸组材料的油酸含量为119.19~143.22 mg/g,亚油酸含量为41.96~53.62 mg/g。上述结果表明,随着芥酸含量的降低,油酸和亚油酸含量逐渐增加。由表1还可看出,饱和脂肪酸和烯酸在不同芥酸组材料间差异较大,无一定的规律性。
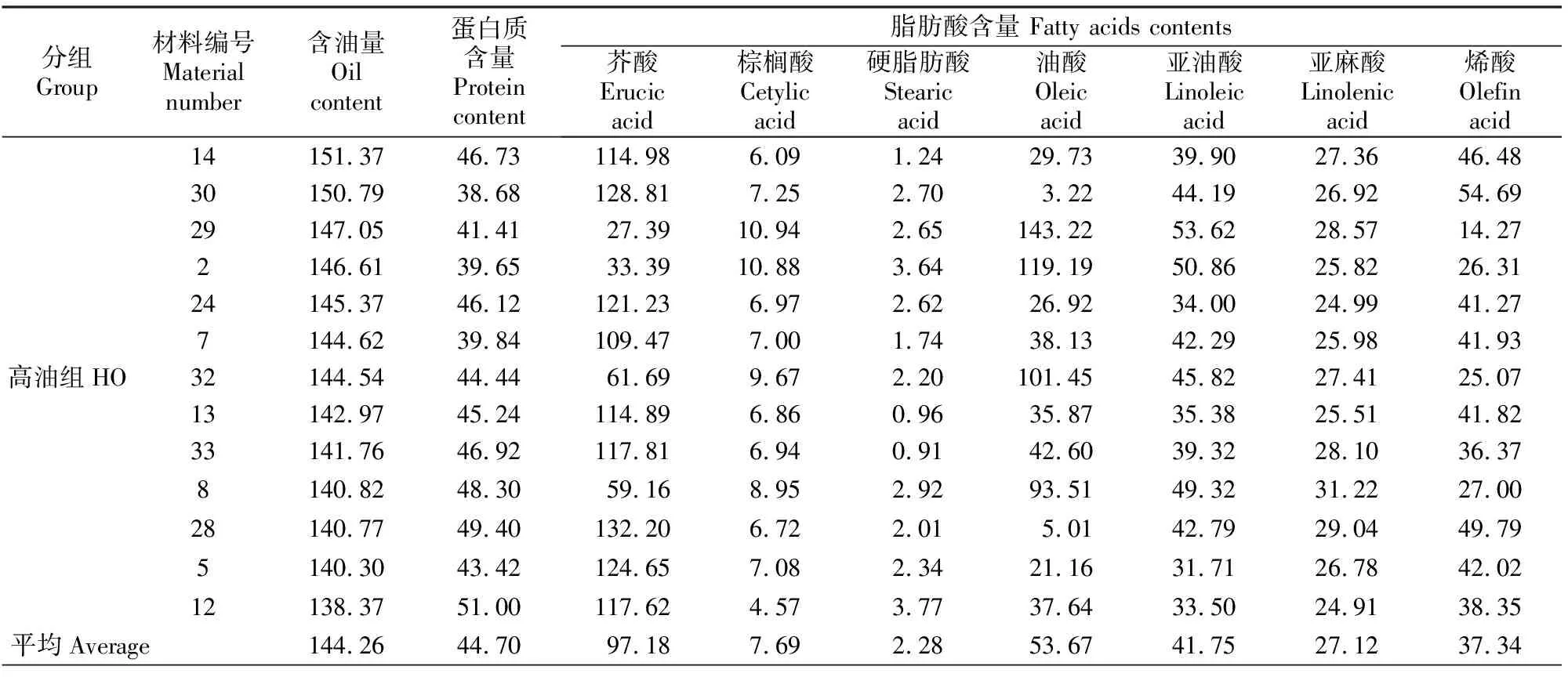
表1 不同白菜型油菜含油量及蛋白质、主要脂肪酸含量
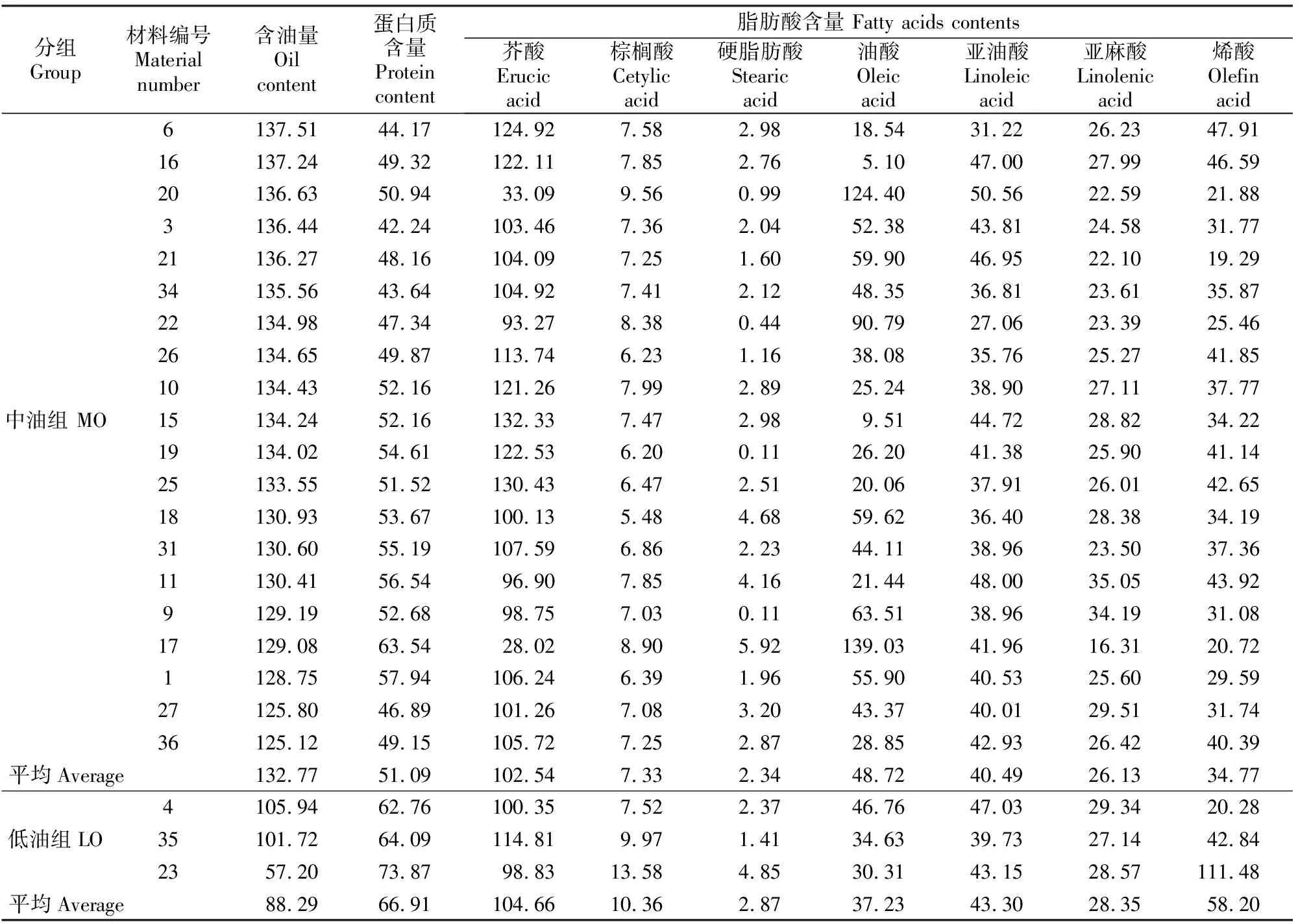
表1(续) Continued table 1
2.2 白菜型油菜油体和油体蛋白特征
2.2.1 油体和油体蛋白形态特征 HO、MO、LO组油体和油体蛋白形态特征如图1所示。

Pb.油体蛋白;Ol.大油体;Os.小油体Pb.Protein body;Ol.Large oil body;Os.Small oil body图1 白菜型油菜种子的油体形态(1 000×)Fig.1 Morphological structure of oil bodies isolated from Brassica campestris L.seeds (1 000×)
由图1可以看出,在成熟的白菜型油菜种子中,油体以近圆形为主,少量呈扁圆形和不规则形状;油体数量较多,充满了整个视野且趋向聚集状态,二维面积和直径有很大差异,其中小油体数量显著高于大油体数量。油体蛋白也多为近圆形,二维面积和粒径较大,但数量较少。
2.2.2 油体和油体蛋白粒径 对分离的油体和油体蛋白粒径进行统计与分析,并进行显微观察,所得结果见表2和图2。由表2和图2可以看出,各材料油体粒径为0.49~8.66 μm,随着供试材料含油量的增加,油体平均粒径表现出增加趋势。HO组各材料油体数量较多,粒径较大,其中最大油体粒径为3.49~8.66 μm,平均油体粒径为1.89~2.78 μm,组内平均粒径为2.12 μm;MO组各材料最大油体粒径为3.17~7.99 μm,平均油体粒径为1.62~2.03 μm,组内平均粒径为1.82 μm;LO组各材料最大油体粒径为1.07~3.12 μm,平均油体粒径为1.39~1.48 μm,组内平均粒径为1.44 μm。
表2和图2显示,HO组各材料油体蛋白粒径相对较大,但数量较少,其中最大油体蛋白粒径为5.45~21.02 μm,平均油体蛋白粒径为2.90~12.15 μm,组内平均粒径为5.24 μm;MO组各材料最大油体蛋白粒径为5.67~18.58 μm,平均油体蛋白粒径为3.36~8.60 μm,组内平均粒径为4.58 μm;LO组各材料油体蛋白粒径相对较小,但数量较多,其中最大油体蛋白粒径为5.01~8.11 μm,平均油体蛋白粒径为2.54~5.78 μm,组内平均粒径为3.81 μm。

表2 不同白菜型油菜的油体和油体蛋白粒径Table 2 Analysis of diameter of oil body and oleosin from Brassica campestris L materials μm
2.3 白菜型油菜种子营养成分与油体、油体蛋白的相关性分析
由表3可以看出,在成熟的白菜型油菜种子中,含油量与蛋白质、棕榈酸、烯酸含量之间呈极显著负相关,其相关系数分别为-0.835,-0.477和-0.568;含油量与油体平均粒径呈极显著正相关,相关系数为0.634,而与油体蛋白平均粒径相关性不显著;蛋白质含量与油体平均粒径呈极显著负相关,相关系数为-0.561;油体平均粒径与油体蛋白平均粒径呈显著正相关,相关系数为0.354;油体平均粒径和油体蛋白平均粒径与脂肪酸含量的关系均无明显规律性。
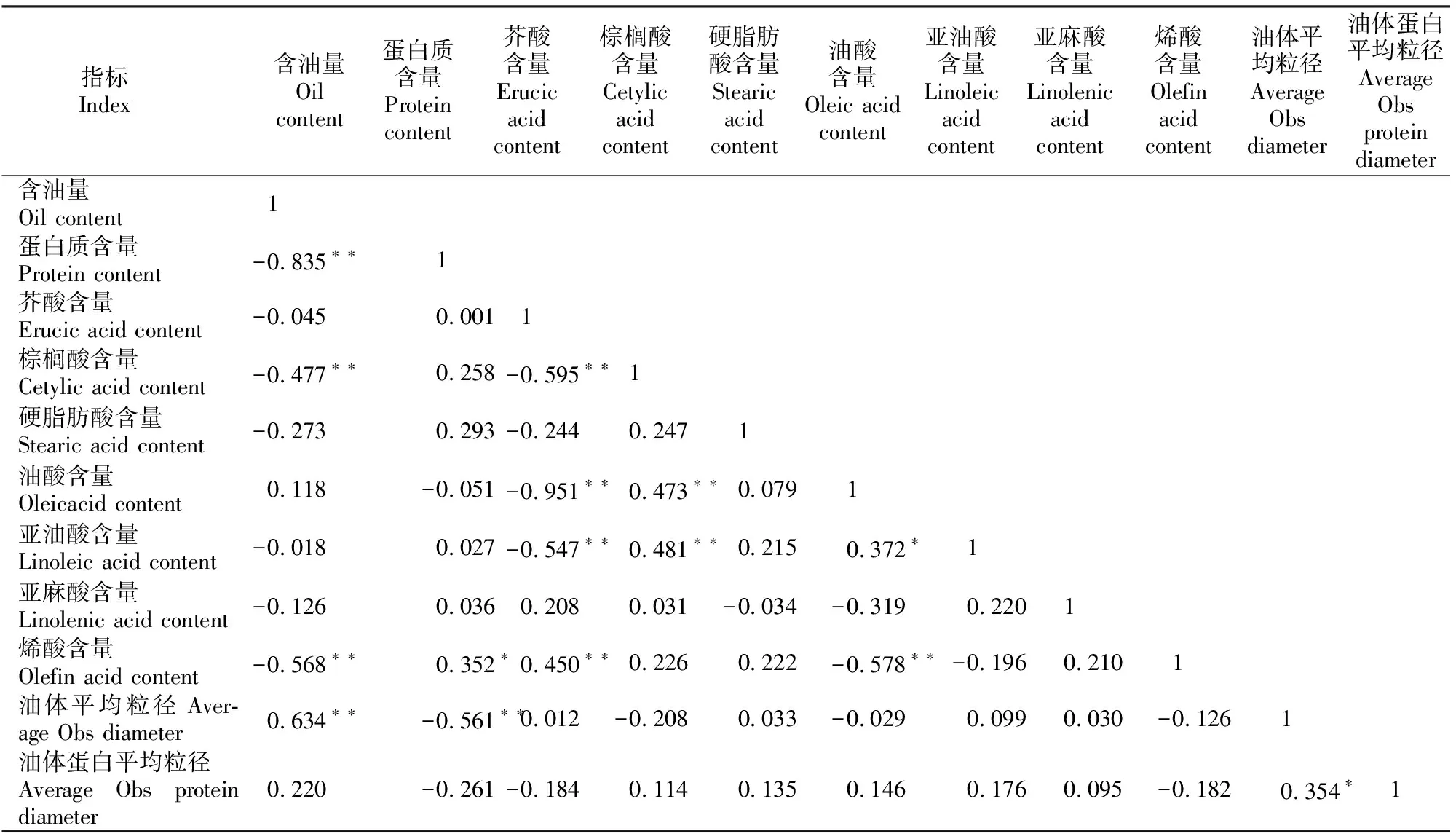
表3 不同白菜型油菜种子含油量及蛋白质、脂肪酸含量与油体和油体蛋白粒径的相关性Table 3 Correlation analysis on contents of seed oil,protein and main fatty acids and average diameter of oil body and oil body protein form different Brassica campestris L.materials
注:**表示在0.01水平上相关性显著(双侧检验),*表示在0.05水平上相关性显著(双侧检验)。
Note:**.Correlation is significant at the 0.01 level (2-tailed),*.Correlation is significant at the 0.05 level (2-tailed).
2.4 白菜型油菜油体粗提液SDS-PAGE电泳分析
由表4和表5可以看出,36份供试材料共分离出蛋白谱带740条,各供试材料蛋白谱带条数为12~26条,平均每份材料的谱带数为20.5条,其中29和13号材料蛋白谱带数最多,均为26条,3号材料最少,为12条。按照迁移率大小,共分离出迁移率不同的谱带32 条,各谱带在供试材料中出现的频率为5.6%~100.0%,其中13,19,20,21和24号谱带在供试材料中均有出现,出现频率为100%,是共有带;11,15,22和31号谱带在供试材料中出现频率较低,均低于14%。

表4 不同白菜型油菜油体粗提液中的蛋白亚基数量Table 4 Protein subunit number of oil body crude extracts form different Brassica campestris L.materials
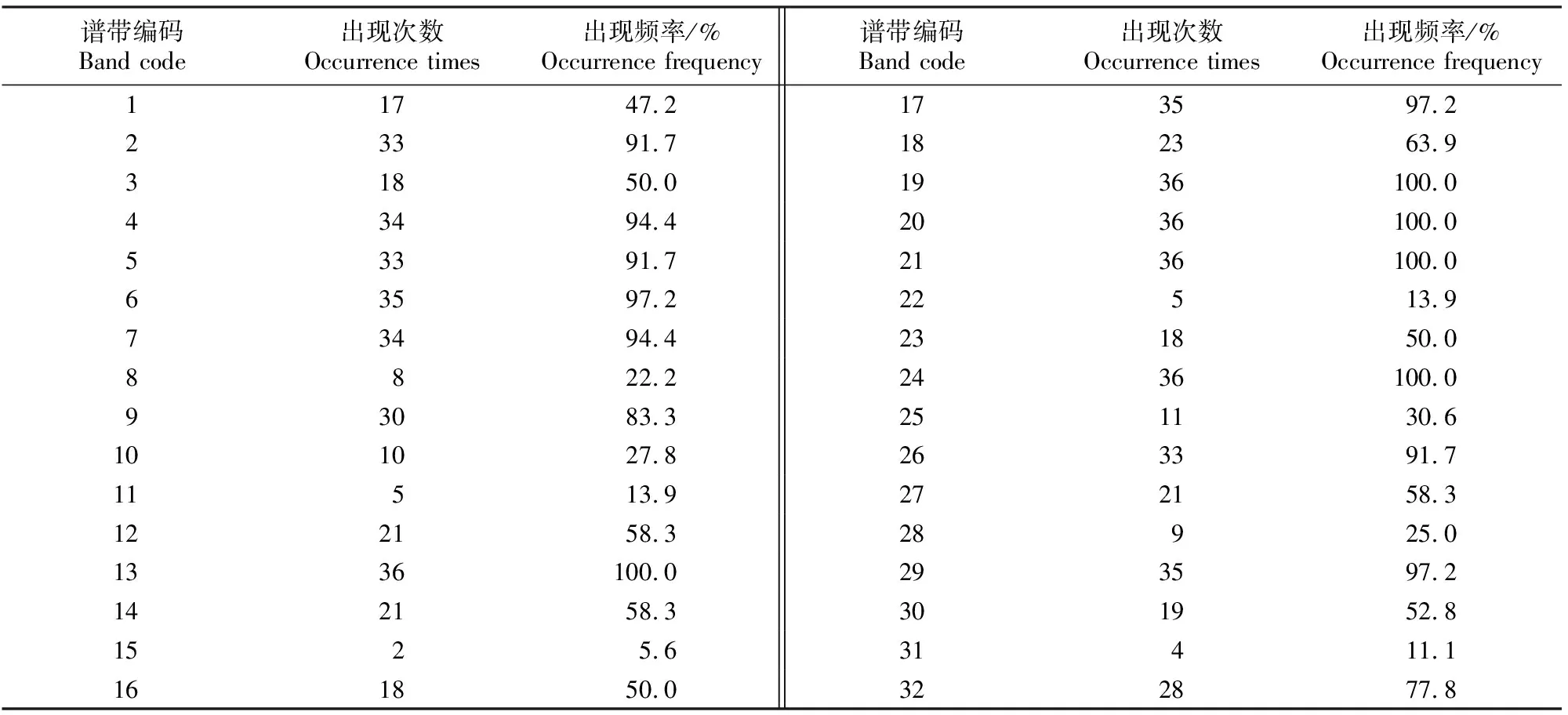
表5 不同白菜型油菜油体粗提液中蛋白亚基的出现频率Table 5 Frequency of different subunits of protein in crude extracts of oil bodies form different Brassica campestris L.
SDS-PAGE电泳结果(图3)表明,36 份供试材料的蛋白谱带具有一定的多态性。蛋白各亚基相对分子质量为25.5~128.7 ku,其中谱带较集中区域在27.4~87.8 ku。各参试材料在68.32,51.33,41.23,32.82和27.44 ku处的谱带恒定且明显。这些数据表明,西藏白菜型油菜种质在蛋白水平上存在着一定的遗传变异,蛋白亚基数量和组合方式变化丰富,存在着广泛的等位基因变异。
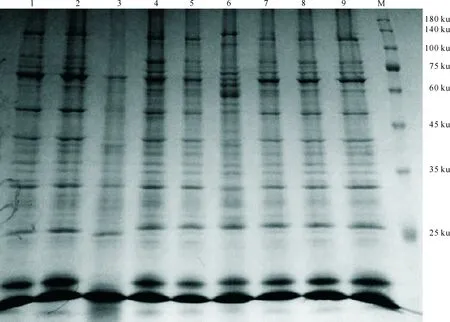
1~9.分别代表1~9号供试材料;M.蛋白质Mark1-9.Represent 1-9 test materials;M.Represents protein Mark
2.5 白菜型油菜油体蛋白的遗传相似性分析
材料间遗传相似系数越大,表明遗传距离越小,亲缘关系越近。N-J进化树分析结果(图4)显示,西藏36 份白菜型油菜油体蛋白的遗传相似系数为0.61~0.98,表明36份供试材料间遗传背景差异较大。13和14号材料、17和18号材料间的遗传相似系数最大,分别为0.98和0.975,表明这2组材料间的遗传距离较小,亲缘关系较近。1和3号材料间遗传相似系数最小,为0.61,但遗传距离最大,亲缘关系最远。在遗传相似系数为0.835时可将36份供试材料聚为9个类群,其中第Ⅰ类群有3份材料;第Ⅱ类群最大,有15份材料;第Ⅲ类群有3份材料;第Ⅳ类群包括3份材料;第Ⅴ类群有2份材料;第Ⅵ类群有2份材料;第Ⅶ类群有6份材料;第Ⅷ类群有1份材料;第Ⅸ类群有1份材料。
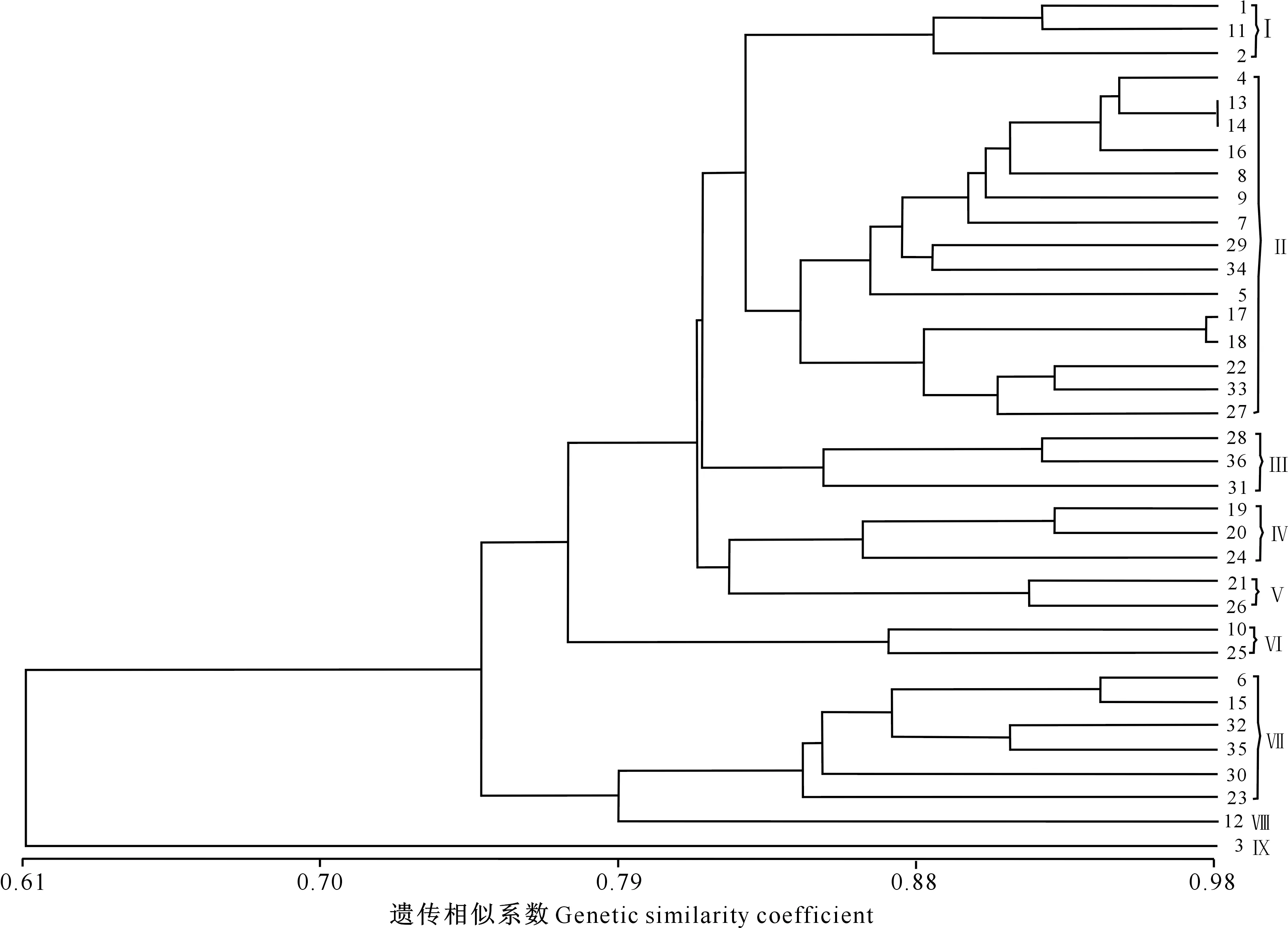
图4 36份西藏白菜型油菜油体粗提液蛋白亚基带型的N-J 进化树分析Fig.4 N-J phylogenetic tree clusters of protein band patterns from crude extracts of oil bodies form 36 Brassica campestris L.materials
3 讨 论
油体是植物贮藏脂质的最重要细胞器,其大小取决于物种、品种和外界环境因素,在一般作物种子中,油体是平均粒径为0.5~2.5 μm的小球体[1,24]。Jolivet等[32]利用光学显微镜发现,拟南芥种子油体均为球形,且大小相似,粒径为(2.81±0.72)μm。韦存虚等[33]研究认为,白菜型油菜种子油体粒径为0.1~6.0 μm,为便于统计,以粒径1.5 μm作为区分大小油体的分界线,>1.5 μm的大油体的平均粒径为2.39 μm,≤1.5 μm的小油体的平均粒径为0.57 μm,其中0.4~0.5 μm的油体数量最多,粒径>0.5 μm的油体数量随着粒径的增加而逐渐减少。本研究结果表明,西藏白菜型油菜油体多呈近圆形,并有少量扁圆形和不规则形状的油体,油体粒径为 0.49~8.66 μm,按照韦存虚等[33]的方法将油体分为大和小2种类型,其中粒径<1.5 μm的小油体数量居多,粒径>1.5 μm的大油体粒径多集中在1.5~2.5 μm,粒径大小因材料不同而表现出一定差异。此结果与Tzen等[7]报道的甘蓝型油菜、芥菜型油菜、花生、亚麻等植物种子油体粒径0.6~2.0 μm相比有一定差异,但与韦存虚等[33]报道的白菜型油菜种子油体粒径相接近。此外,供试材料均有数量较少但粒径>5 μm的巨大油体,推测为异常大油体。Hu等[9]推测认为,在甘蓝型油菜种子中,异常大油体(粒径>5 μm)与低含油量高度相关,但本研究尚不支持这个结论。本研究发现,西藏白菜型油菜种子油体粒径偏大,这可能与本研究采用光学显微镜在放大1 000倍视野下对油体粒径进行统计时,部分极小油体无法有效统计有关,也可能与西藏特殊的地理环境有关,高原缺氧及强辐射条件导致参试材料油体中形成了较多异常大油体[1,9,26]。Heneen等[34]观察燕麦胚乳发育时发现,大油体是由小油体融合形成的,且认为种子含油量与粒径>1.5 μm的油体相关。因此白菜型油菜大油体的产生机理、油体大小与含油量的关系等还有待进一步研究。
本研究发现,西藏白菜型油菜种子含油量和蛋白质含量呈极显著负相关,这与Hu等[9]和Cober等[35]等的研究结果一致。油体、油体蛋白平均粒径分析显示,平均油体粒径表现为高油组>中油组>低油组,且油体蛋白也表现出相似趋势。相关分析表明,含油量与蛋白质、棕榈酸、烯酸含量呈极显著负相关,与油体平均粒径呈显著正相关,但与油体蛋白平均粒径相关性不大;油体平均粒径与油体蛋白平均粒径呈显著正相关,但两者均与脂肪酸含量相关性不大。该结果与Hu等[9]的研究结果相反,但与Ting等[8]和韦存虚等[33]的研究结果相似。
油体蛋白在纯化制备过程中易受到储藏蛋白的污染,故难分离得到较纯的油体蛋白[9]。本研究对油体粗提液进行SDS-PAGE电泳分析,分离出了清晰可辨的蛋白谱带,其分子质量为25.5~128.7 ku,其中包括27 ku的油体钙蛋白、37 ku的油体固醇蛋白和39 ku的油体蛋白,但已报道的油体结合蛋白均为小分子质量蛋白,说明该油体粗提液富含较多的贮藏蛋白,这与Jolivet等[32]对拟南芥种子油体粗提液SDS-PAGE电泳分析的结果类似。基于种子蛋白质的SDS-PAGE电泳分析方法,已经成为一种解决植物分类和进化问题、品种和变种的鉴定、种质特征分析和相关评价信息补充的有效工具[36-37]。本研究中,油体粗提液蛋白谱带的多样性提示供试材料的遗传背景具有一定的差异。N-J进化树分析表明,在相似系数为0.835时可将36份西藏白菜型油菜种质材料分为9个类群,不同类群间和同一类群的不同材料间含油量、蛋白质含量、脂肪酸含量并未表现出相近的规律,也未表现出明显的地域分布,这可能归因于材料生境的差异[38]。
