南淝河表层沉积物细菌群落结构特征及驱动因素
2018-09-25杨长明吴亚琼王育来郭韵昊
杨长明,吴亚琼,王育来,郭韵昊
南淝河表层沉积物细菌群落结构特征及驱动因素
杨长明1*,吴亚琼1,王育来2,郭韵昊1
(1.同济大学教育部长江水环境重点实验室,上海 200092;2.安徽工业大学能源与环境,安徽 马鞍山 234000)
为了探讨南淝河不同河段沉积物细菌群落结构特征差异及其主要驱动因素,在对南淝河不同土地利用区域河段表层沉积物的主要理化指标(pH、CEC、TOC、DOC、氮分级形态)进行分析基础上,重点通过MiSeq高通量测序技术对沉积物中细菌群落结构特征进行了分析,并利用冗余分析(RDA)方法探讨了导致群落结构差异的主要驱动因素.结果表明:沉积物中门水平下优势细菌种群为变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)和拟杆菌门(Bacteroidetes);纲水平下除未分类外主要是厌氧绳菌纲(Anaerolineae)、γ-变形菌纲(Gammaproteobacteria)、梭状芽胞杆菌纲(Clostridia)、δ-变形菌纲(Deltaproteobacteria),此外ε-变形菌纲(Epsilonproteobacteria)主要在南淝河下游采样点被检测到,说明其在南淝河沉积物生态系统中并不是普遍存在的;属水平下大多为未分类序列,长绳菌属()和芽孢杆菌属()有优势地位.南淝河不同河段所在流域土地利用方式不同会导致污染物类型、污染源强和沉积物理化指标的不同,进而影响沉积物细菌群落结构,中游城区和工业区段的细菌多样性整体上高于上、下游;RDA分析结果表明,表层沉积物的CEC、TOC、DOC、TP、WAEF-N和IEF-N是导致细菌群落结构特征差异的主要影响因子,其中变形菌门、拟杆菌门和厚壁菌门受TP影响较大,绿弯菌门则受弱酸浸取态氮(WAEF-N)影响较大.
南淝河;表层沉积物;细菌群落结构;MiSeq高通量测序;冗余分析(RDA)
城市河流沉积物是陆源污染物(营养物、重金属、有机污染物等)的主要储存场所[1].沉积物是城市重污染河流内源污染的重要来源,具有产生二次污染的潜在危险,其特殊的生境条件使其具有丰富的微生物资源,而微生物是沉积物和上覆水进行物质循环的重要驱动力[2].微生物在城市内河沉积物物质循环及污染物释放过程中发挥重要作用,其群落结构特征也能够作为评估城市河道污染负荷与河流生态系统整体健康状态的主要根据[3].
传统的环境微生物研究方法主要依赖分离和培养,不能很好地揭示微生物的群落结构,因此分子生物学研究方法逐渐被大量采用.已有学者运用PCR-DGGE[4]、T-RFLP[5-6]等技术研究了沉积物微生物群落结构分布,然而近些年高通量测序技术被广泛应用,该技术不仅数据通量大,而且准确度相对较高.部分学者运用高通量测序技术对程海[7]、太湖[8]等水体的沉积物细菌群落结构进行了分析,但应用该技术对跨越不同土地利用类型,并具有复杂污染源强的城市型河流沉积物细菌群落结构及其主要驱动因子的研究还鲜有报道.
南淝河为巢湖的一条重要入湖河流,其上游植被覆盖以林地为主,中游主要为老城区和工业园区,下游主要是郊区、农田和湿地,土地利用方式呈现多样化格局.南淝河水体污染状况严峻,水质为劣V类,前期政府部门实施了一些治理工程,包括城市污水管网建设和改善工程、污水厂扩建和提标工程,这在一定程度上较好地管理和控制了外源污染输入,然而南淝河的水体水质并没有很大的提高,这和沉积物内源污染物的不断释放密切相关[9].已有学者研究了南淝河沉积物有机污染物分布和生态风险[10]、水环境质量[11]、不同形态磷含量分布[12]以及不同季节异养细菌的总量[13],但对细菌群落结构特征及其与理化指标关系的相关报道较少.
本研究在对南淝河表层沉积物基本理化指标进行分析的基础上,利用MiSeq高通量测序技术对南淝河沉积物细菌群落结构特征和多样性进行分析,并将细菌门群落结构、多样性指标和沉积物基本理化指标进行冗余分析,进而全面阐明南淝河沉积物微生物群落结构特征及其主要的驱动因素,为今后利用分子生物学手段评价和指示沉积物污染特征和河流健康水平提供一定的依据.
1 材料与方法
1.1 样品采集与处理
南淝河是流经合肥市的巢湖一级支流,主河道全长70km,上游为森林公园,中游流经合肥主城区,下游流域主要为农田和湿地, 沿岸分布有较多码头.研究区域位于整个南淝河(北纬31.69°~31.88°,东经117.22°~117.42°),考虑到研究区域内土地利用类型以及排口类型的不同所导致的入河污染负荷和组成可能存在差异,本研究分别选择城市雨水排口、垃圾填埋场、雨污合流排口、污水厂尾水排口、泵站、船舶加油站以及支流汇入口(农业排水)等共17个采样点,各采样点位置及所在流域基本信息如图1和表1所示.
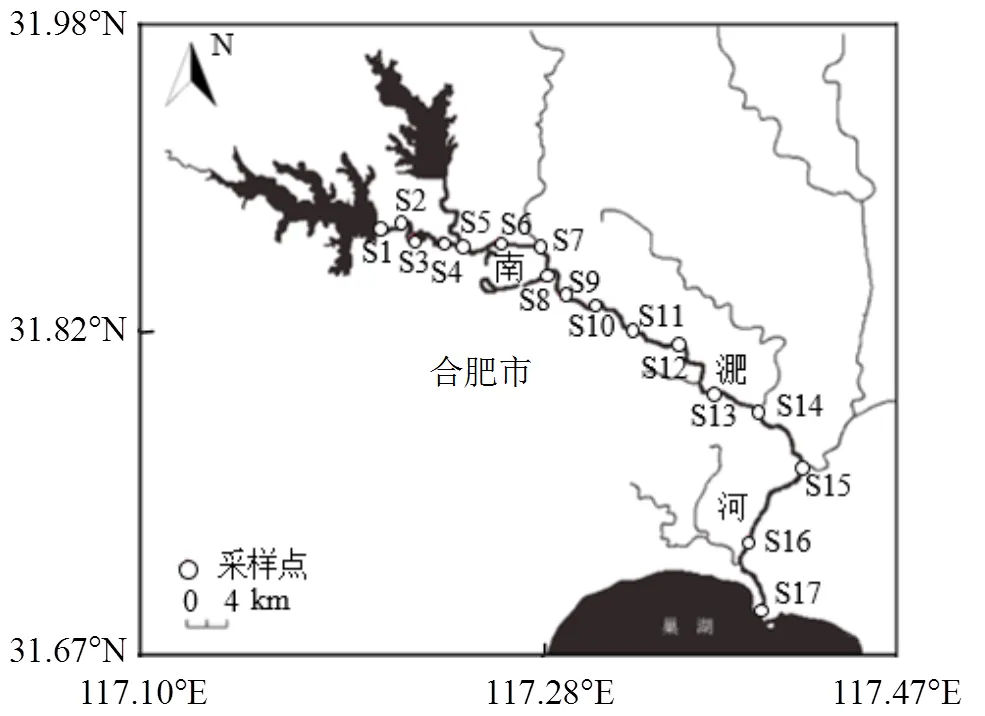
图1 南淝河沉积物采样点空间分布

表1 不同沉积物采样点基本概况
于2016年3月分别采集17个样点的表层沉积物样品和上覆水样,表层沉积物采用抓斗采样器(采样深度0~10cm),每个采样点采集3个沉积物样并混合均匀,新鲜样采集后装于聚乙烯封口袋中并编号,放入冷藏箱带回实验室,一部分样品冷冻干燥研磨后过100目筛,置于聚乙烯封口袋中并在避光状态下低温保存,用于理化指标分析;一部分样品冻存(-20℃)以备后续微生物群落结构分析;同时收集上覆水,采用便携式参数水质分析仪(哈希SensIon156,美国)现场测定pH值和溶解氧(DO),其余指标在实验室内测定.
1.2 样品分析
1.2.1 样品理化指标的测定 上覆水的总磷(TP)使用钼酸铵分光光度法(GB 11893-89),总氮(TN)使用碱性过硫酸钾消解紫外分光光度法(HJ 636- 2012),氨氮(NH4+-N)使用纳氏试剂分光光度法(HJ 535-2009),叶绿素a使用丙酮法.沉积物的pH值用酸度计(pHS-3C型)测定,阳离子交换量(CEC)采用氯化钡-硫酸强迫交换法测定,总有机碳(TOC)和溶解性有机碳(DOC)用总有机碳测定仪(岛津TOC- VCPH系列)测定,TP采用碱熔-钼锑抗分光光度法(HJ 632-2011)测定. 沉积物中的4种可转化态氮(TF-N)分为离子交换态氮(IEF-N)、弱酸浸取态氮(WAEF-N)、强碱浸取态氮(SAEF-N)和强氧化剂浸取态氮(SOEF-N),采用王圣瑞等[14]的连续分级提取的方法测定.
1.2.2 沉积物细菌群落结构分析 沉积物样品DNA的提取: 本研究沉积物基因组DNA的提取采用OMEGA土壤提取试剂盒进行提取,利用琼脂糖凝胶电泳检测DNA完整性,并采用Qubit2.0DNA检测试剂盒准确测定基因组DNA的含量,以确定PCR反应应该加入的DNA量.
PCR扩增:选用V3~V4区的引物对提取的DNA进行PCR扩增,引物序列(5’-3’)如下:341F (CCCTACA-CGACGCTCTTCCGATCTG),805R(GACTGGAGTTCCTTGGCACCCGAGAATTCCA),并利用barcode区别样品序列.PCR反应条件为:95℃预变性30s;95℃ 15s,55℃ 15s,72℃30s,5个循环;72℃5min.PCR产物进行琼脂糖凝胶电泳检测,并用PCR产物纯化试剂盒纯化.
MiSeq高通量测序高通量测序:扩增后的细菌DNA进行宏基因组建库、测序,然后对所得的数据进行生物信息学分析.扩增区域为 V3~V4 (引物为341F, 805R);基于MiSeq 2x300测序平台的高通量测序由生工生物工程(上海)股份有限公司测定完成,该测序平台单次运行可完成15Gb的数据测序,最大读长2´300bp,片段数单端25M,双端50M.对各样品序列做质量控制,去除短片段和低复杂度序列,使用Mothur软件去除非靶区域序列和嵌合体.
1.3 数据处理与统计分析
试验数据采用Excel 2010和SPSS 19.0统计软件进行处理,所有试验数据通过ANOVA变异分析程序进行统计分析,采用邓肯新复检验法对采样点上覆水、沉积物理化指标差异进行统计估计,在<0.05的概率水平视为显著.
在相同测序深度下采用Uclust软件进行操作分类单元(OUT)聚类;使用Mothur软件分析多样性,计算Shannon指数、ACE指数、Chao指数、Coverage指数和Simpson指数;运用加权重的Unifrac算法进行主坐标分析(PCoA);通过RDP classifier把序列进行物种分类;运用R语言绘制热图,并对细菌门和理化指标进行冗余分析(RDA).
2 结果与讨论
2.1 不同采样点上覆水水质特征

表2 不同采样点上覆水主要水质指标

续表2
由表2显示,南淝河不同采样点上覆水pH值变化范围为7.18~9.02,偏弱碱性;溶解氧(DO)变化范围为0.15~7.54mg/L,其中共有9个样本溶解氧含量远小于2mg/L,且多集中在中游城区段;叶绿素(chl-a)变化范围为0.01~0.06mg/L.总氮(TN)含量为32.20~ 51.84mg/L;氨氮(NH3-N)为0.69~12.69mg/L;总磷(TP)含量普遍较高,其变化范围分别为和0.25~ 2.54mg/L.结合国家地表水环境质量标准(GB3838- 2002),南淝河各采样点上覆水水质指标均未达到地表水V类标准,流经合肥老城区的中游河段水质较差,特别是NH3-N含量显著高于上游和下游河段.
2.2 不同采样点沉积物理化特征
采用SPSS软件对上、中、下游的理化指标进行显著性差异分析表明(表3),南淝河上、中、下游表层沉积物CEC、TOC、IEF-N和WAEF-N差异显著(<0.05).pH值变化范围为7.24~8.04,偏弱碱性,而通常微生物通过代谢活动对有机质进行分解,释放出的CO2和有机酸理论上会使pH值偏弱酸性,但所有样本点pH值均偏弱碱性,这可能是因为采样点区域支流众多,彼此之间有较强的水力联系,有机质降解所产生的酸会受到稀释,且人类活动频繁,pH值则会更多地受周围工业废水和生活污水排入的影响.CEC表征了沉积物胶体的物理化学吸附性能,对生源要素在沉积物和上覆水界面的转化过程具有重要影响,南淝河各采样点CEC变化范围为30.98~155.07cmol/kg,在一定程度上表征了南淝河沉积物具有较好的缓冲性能,其中S12的CEC值最高,可能是因为该点为农业集中区,众多农业活动可能会产生影响.
沉积物TOC可以较全面地反映水体受有机物污染的程度,其来源可能是水体自养生物的合成和陆源性污染物质的输入[15].南淝河不同采样点TOC含量变化范围为12.67~71.34g/kg,其中S9的TOC含量最高,可能是因为该点为污水厂尾水排放口,外源输入的有机质含量较高;且中游TOC的含量明显高于上、下游,表明南淝河市中心附近的有机污染情况加重;位于橡胶坝后的S11~S17样点,TOC含量相对较低,可能是由于水流较快,沉积物泥沙含量较高.DOC含量变化范围为0.253~1.124g/kg,其中南淝河中游采样点表层沉积物DOC最高,上游次之,而下游普遍较低.特别是位于中游的王小郢污水厂尾水排放口S9的DOC高达1.124g/kg,位于望塘污水处理厂尾水排口S4的DOC含量也比较高,可能是因为污水厂尾水排放对沉积物DOC具有一定的积累效应.
磷是水体富营养化的主要限制性营养元素,沉积物是水体中磷的主要贮存场所.南淝河不同采样点表层沉积物TP变化范围为785.58~3384.4mg/kg,平均为2107.4mg/kg,远高于与之相邻的十五里河沉积物[16],说明该区域沉积物中磷对水体有较高的污染风险. S16采样点沉积物TP最高,可能是因为该点位于下游农业集中区,受农业面源污染影响较大,加上该采样点位于店埠河入南淝河河口,而该支流上游有一家大型磷矿企业,其生产排水可能会造成该采样点磷的累积.
有研究表明,沉积物中不同形态氮的环境地球化学行为存在差异[17].南淝河沉积物中可转化态氮含量的关系为SOEF-N>WAEF-N>SAEF-N>IEF-N.有研究表明,SOEF-N含量和有机质及沉积物的氧化还原环境相关性较大,沉积物生境的还原性越强,对有机质的保存越有利,矿化作用越弱,SOEF-N的含量越大[18];WAEF-N的产生和分布主要受碳酸盐和pH的影响[19];SAEF-N的形成及分布受沉积物氧化还原环境的影响较大,当水-沉积物界面为还原状态时容易释放并参加氮循环系统[20].除了位于雨水泵站排口的S6和S8两个采样点外,各采样点中IEF-N是所有可转化态氮形态中含量最低的,可能是该形态氮更容易从沉积物中释放[21].

表3 南淝河采样点表层沉积物理化指标
2.3 不同采样点沉积物细菌群落结构特征

图2 不同采样点沉积物OTU数量分布
2.3.1 测序结果分析 南淝河17个样本共得到365547条有效序列,并在97%相似值下进行OTU聚类,共获得49611种OTU,如图2所示.其中S3,S4,S5,S6,S7,S9,S12,S13样本中OTU数目相对较高,S4和 S9均为尾水排口,数量基本一致且较高;S13为污水厂排口加大型码头,有明显的污水入河,所以OTU数量最多;S12为农业集中区,仅次于S13;整体而言中游河段细菌菌种数量相对于上、下游较高.S8有明显的下降,可能是因为其所在位置有雨水泵站,起到稀释作用,导致附近水体中营养物质含量较低;S10、S11处也有所下降,可能是因为S10处有橡胶坝,且该位置开始进入郊区,人口密度降低,土地利用强度下降,入河污染负荷减弱.
2.3.2 样品多样性分析 (1)α多样性分析:由表4可知,不同采样点沉积物Shannon指数在7.38~8.22之间,ACE指数在21421.72~55107.90之间,Chao1指数在13354.83~25658.03之间.中游样本点的Shannon、ACE、Chao1指数相对上、下游较高,说明中游城区段的细菌多样性较高,其中S4多样性最高,该采样点为望塘污水厂尾水排口,可能是因为该点营养盐含量较高且有利于微生物的生存,S16的Shannon指数最低,这可能与该位点处于南淝河下游且为农业和工业集中区,有毒有害物质较多;各样点沉积物细菌Simpson指数变化范围为0.0008~0.0035,总体变化不大,处于轻微的波动,表明优势种群更替不明显;除S2外,测序的覆盖率均在74%~84%之间,虽然并没有达到100%的覆盖率,但是测序结果对样本的解释仍然在可信范围内,测序深度已经反映出该沉积物中大多数的细菌类群.

表4 各采样点沉积物α多样性指数统计
(2) β多样性分析:选取对样本解释度最高的3个成分分析其样本间的差异,通过PCoA分析结果可以明显看出17个样本中分布关系相近的类群倾向于聚在一起(图3).其中,S1、S14、S15、S16、S17距离较近,S3、S6、S11、S12、S13距离较近,S4、S5、S7、S8、S9、S10相对集中,S2异质性较高,整体而言,各样本间的细菌群落结构存在差异,且中游段位点间差异较上、下游相对较小.
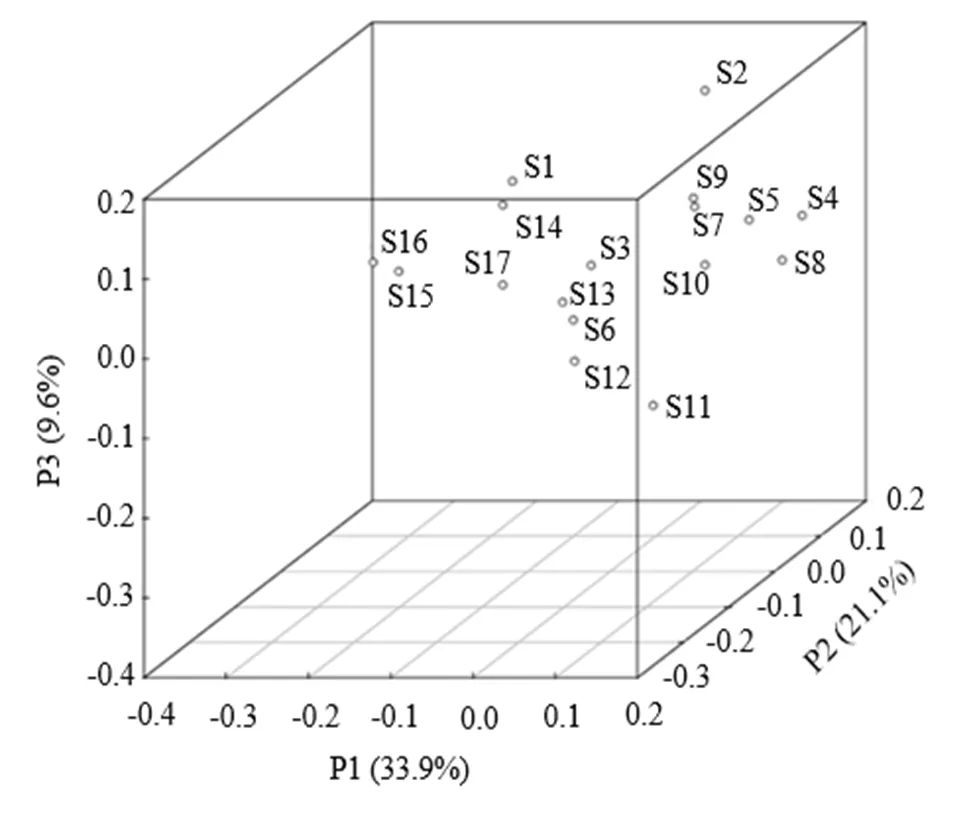
图3 不同采样点沉积物主坐标分析
2.3.3 细菌群落结构分析 (1)门水平下细菌群落结构
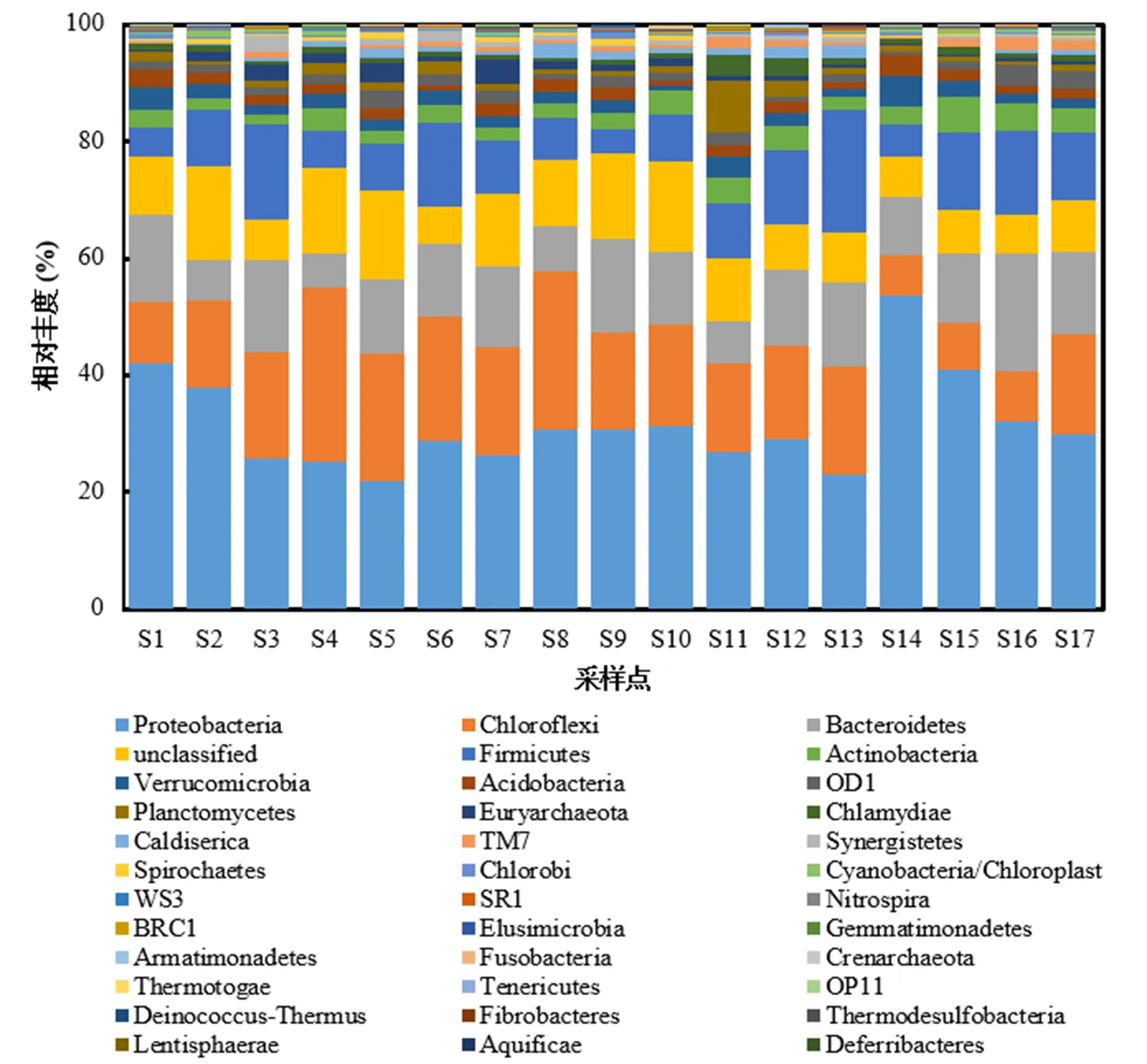
图4 沉积物样品中门水平所有样本菌群分布
图4为南淝河不同采样点表层沉积物门水平下细菌群落结构.由图4可知,沉积物中门水平主要有35个细菌门,其中最优势细菌类群为变形菌门(相对丰度为21%~54%),其次是绿弯菌门(相对丰度为6%~30%)、拟杆菌门(相对丰度为5%~20%)、厚壁菌门(相对丰度为4%~20%),这些细菌门在滇池[22]、太湖[23]、水库[24]等中也有发现;同时放线菌门、疣微菌门、酸杆菌门、浮霉菌门、衣原体、互养菌门、蓝藻门等均有发现.此外,S14样本点变形菌门含量达到了50%以上,可能与处于重度污染的二十埠河的汇入有关;S4是唯一绿弯菌门含量比变形菌门含量高的采样点,可能是因为望塘污水厂尾水水质与其他样点水质存在差异.
(2)纲水平下细菌群落结构:图5为南淝河不同采样点表层沉积物纲水平下细菌群落结构.由图5可知,沉积物中纲水平主要有49个类群,其中厌氧绳菌纲、γ-变形菌纲、梭状芽胞杆菌纲、δ-变形菌纲为优势菌群,其次含量较高的有β-变形菌纲(Betaproteobacteria)和α-变形菌纲(Alphaproteobacteria),且只在S3、S15、S16、S17样本中发现了ε-变形菌纲,说明其在南淝河沉积物中并不是普遍存在,可能与某种特殊污染物有关.过去研究表明在受碳氢化合物尤其是多环芳烃(PAHs)污染的Apapa沉积物中,ε-变形菌纲的丰度最高[25]. γ-变形菌纲则是太湖富营养化严重时的优势菌群[26].
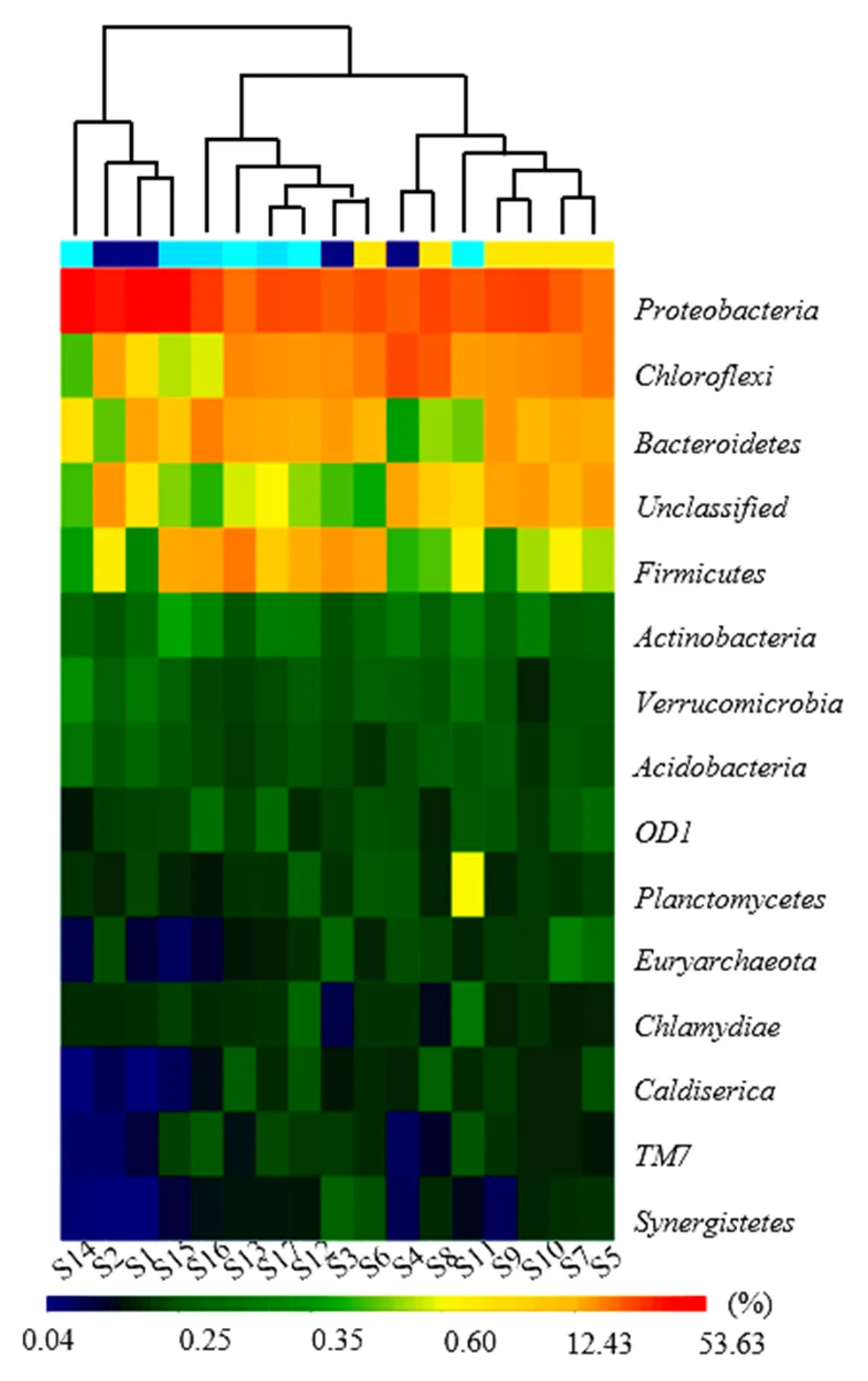
图6 沉积物样品中属水平物种丰度
(3)属水平下细菌群落结构:将样本和属水平下的菌群信息进行聚类后绘制热图(图6),如图6所示,南淝河表层沉积物中属于绿弯菌门中厌氧绳菌纲的、、,属于变形菌门中γ-变形菌纲的、δ-变形菌纲的、β-变形菌纲的,属于拟杆菌门中鞘脂杆菌纲的、黄杆菌纲的丰度相对较高. Bai等[25]利用454高通量测序技术发现在有机污染重的滇池草海中和为优势菌群.此外,S5、S7、S8、S11、S12和S2、S9、S10以及S4聚为一类,S3、S6、S13和S15、S16、S17以及S1、S14聚为一类,整体上属水平下中游河段沉积物细菌群落结构相似性较高.
2.4 细菌群落结构与理化指标的相关性分析
将门水平下样本中的细菌物种与沉积物的理化指标进行RDA分析,结果如图7所示.第一轴的主要影响因子是CEC、TP、WAEF-N、IEF-N和DOC,第二轴的主要影响因子是TOC和DOC, 即说明CEC、TOC、DOC、TP、WAEF-N和IEF-N对南淝河沉积物中的细菌群落结构影响较大.过去也有研究表明,有机质、TOC、TN、TP以及C/N是影响细菌群落结构的主要因子[27-30],本研究结果总体与上述相似,但与Wu等[31]对有机碳的分析结果存在差异.另外,本研究通过不同形态氮与细菌群落结构的RDA分析结果显示,WAEF-N对细菌群落结构影响最大.

图7 门水平下沉积物RDA分析
由图7还可发现,17个样本点分布相对离散,说明微生物群落结构特征存在差异,且距离原点较远,说明受理化指标影响较大;其中,S4、S8和S7的细菌群落结构特征与TOC和DOC呈正相关;S1、S9、S15和S17细菌群落结构相似,聚集在第二象限,受pH值影响较大;S14则主要受SAEF-N 影响;S2和S11主要与WAEF-N、IEF-N呈正相关;S5和S13主要与CEC呈正相关;S3、S6、S10和S12聚集于第一象限,主要与TP呈正相关,其中S6和S10的相对位置距离较近,说明相似度较高,二者均设有泵站或橡胶坝等水利设施,可能有利于有机质的积累与微生物的增长.此外,除相对丰度较高的4种细菌外,绝大部分菌种聚集分布在靠近原点的位置,说明它们之间有较强的依存关系,且理化指标对它们的影响较小.但变形菌门相对丰度与TP呈正相关,拟杆菌门和厚壁菌门的相对丰度与TP、CEC、IEF-N和WAEF-N呈正相关, 其中主要影响因子为TP.变形菌门、拟杆菌门和厚壁菌门相对丰度均与TOC和DOC呈负相关;绿弯菌门与WAEF-N、IEF-N、CEC和TP呈正相关,且主要影响因子为WAEF-N. 分析结果进一步表明,沉积物中的优势细菌群落与部分理化指标关系密切.
3 结论
3.1 南淝河上覆水质为劣Ⅴ类,其中氨氮超标严重.不同采样点表层沉积物样的CEC、TOC、IEF-N和WAEF-N差异显著,其中位于中游的老城区河段采样点TOC和DOC明显高于上游和下游采样点;可转化态氮含量关系为SOEF-N>WAEF-N>SAEF-N>IEF-N,其中SOEF-N是沉积物可转化氮素中主要形态.
3.2 南淝河中游城区段细菌的多样性和相似性均高于上、下游;不同采样点表层沉积物中门水平优势菌群为:变形菌门、绿弯菌门、拟杆菌门、厚壁菌门;纲水平优势菌群为厌氧绳菌纲、γ-变形菌纲、梭状芽胞杆菌纲、δ-变形菌纲;属水平优势菌群为长绳菌属和芽孢杆菌属.
3.3 RDA分析结果表明,沉积物的CEC、TOC、DOC、TP、WAEF-N和IEF-N为导致细菌群落结构特征差异的关键影响因子;此外,变形菌门、拟杆菌门和厚壁菌门受TP影响较大,绿弯菌门则受WAEF-N影响较大.以上结论为今后深入研究沉积物细菌种群与环境因素的关系提供一定参考.
[1] Kevin G T, Philip N O. Sediments in urban river basins: a review of sediment contaminant dynamics in an environmental system conditioned by human activities [J]. Journal of Soils Sediments, 2009,9(4):281-303.
[2] 屈建航,李宝珍,袁红莉.沉积物中微生物资源的研究方法及其进展 [J]. 生态学报, 2007,6:2636-2641.
[3] 沈 烁,杨长明,成水平.PCR-DGGE分析合肥市塘西河表层沉积物细菌群落结构空间分布特征 [J]. 应用与环境生物学报, 2015, 1:80-87.
[4] 牛凤霞,吉芳英,赵 艮,等.龙景湖沉积物的细菌群落垂向分布特征.中国环境科学, 2017,37(6):2322-2331.
[5] Zhao D, Huang R, Zeng J, et al. Diversity analysis of bacterial community compositions in sediments of urban lakes by terminal restriction fragment length polymorphism (T-RFLP) [J]. World Journal of Microbiology & Biotechnology, 2012,28(11):3159-3170.
[6] 刘 睿,周孝德.水沙环境变化对季节性多沙河流沉积物菌群特征的影响 [J]. 中国环境科学, 2017,37(11):4342-4352.
[7] 丁 春,盛周君.基于主成分分析法的南淝河水质综合评价 [J]. 安徽农业科学, 2007,35(35):11583-11584.
[8] 李 慧,曹秀云,宋春雷,等.巢湖及其入湖河流(南淝河)沉积物磷形态与吸附行为的垂直变化 [J]. 长江流域资源与环境, 2012(s2):3-9.
[9] 王永霞.云南高原程海湖沉积物中的细菌多样性研究 [D]. 昆明:云南大学, 2014.
[10] 蔡林林,周巧红,王 川,等.南淝河细菌群落结构的研究 [J]. 环境科学与技术, 2012,3:1-6.
[11] 陈 楠.太湖沉积物微生物群落组成与物质循环及藻华爆发的相关性 [D]. 北京:中国农业大学, 2015.
[12] 沈 烁.南淝河不同排口表层沉积物DOM、氮形态及反硝化细菌群落特征 [D]. 上海:同济大学, 2015.
[13] 邓 琳,李宏卓,廖达伟,等.南淝河沉积物有机物污染及其生态风险评价 [J]. 环境科学与技术, 2011,8:179-184.
[14] 丁 春,盛周君.基于主成分分析法的南淝河水质综合评价 [J]. 安徽农业科学, 2007,35(35):11583-11584.
[15] 李 慧,曹秀云,宋春雷,等.巢湖及其入湖河流(南淝河)沉积物磷形态与吸附行为的垂直变化 [J]. 长江流域资源与环境, 2012,(s2): 3-9.
[16] 蔡林林,周巧红,王 川,等.南淝河细菌群落结构的研究 [J]. 环境科学与技术, 2012,3:1-6.
[17] 王圣瑞,金相灿,焦立新.不同污染程度湖泊沉积物中不同粒级可转化态氮分布 [J]. 环境科学研究, 2007,20(3):52-57.
[18] Cho Y S,Lee W C, Kim J B, et al. Establishment of environmental assessment using sediment total organic carbon and macrobenthic polychaete community in shellfish farms [J]. Journal of the Korean Society of Marine Environment and safety, 2013,19(5):430-438.
[19] 李如忠,李 峰,周爱佳,等.巢湖十五里河沉积物氮磷形态分布及生物有效性 [J]. 环境科学, 2012,33(5):1503-1510.
[20] 沈洪艳,张绵绵,倪兆奎.鄱阳湖沉积物可转化态氮分布特征及其对江湖关系变化的响应 [J]. 环境科学, 2015,36(1):87-93.
[21] 马红波,宋金明.渤海南部海域柱状沉积物中氮的形态与有机碳的分解 [J]. 海洋学报, 2002,24(5):64-70.
[22] 王 梅,刘 琰,郑丙辉,等.城市内河表层沉积物氮形态及影响因素-以许昌清潩河为例 [J]. 中国环境科学, 2014,34(3):720-726.
[23] Wang S, Jin X, Jiao L, et al. Nitrogen fractions and release in the sediments from the shallow lakes in the middle and lower reaches of the Yangtze River area, China [J]. Water, Air, and Soil Pollution, 2008,187(1-4):5-14.
[24] Wang P F, Zhao L, Wang C, et al. Nitrogen distribution and potential mobility in sediments of three typical shallow urban lakes in China [J]. Environmental Engineering Science, 2009,26(10):1511-1521.
[25] Bai Y, Shi Q, Wen D, et al. Bacterial Communities in the Sediments of Dianchi Lake, a Partitioned EutropHic Waterbody in China [J]. Plos One, 2012,7(5):e37796-e37796.
[26] Shao K, Gao G, Qin B, et al. Comparing sediment bacterial communities in the macrophyte-dominated and algae-dominated areas of eutrophic Lake Taihu, China [J]. Canadian Journal of Microbiology, 2011,57(4):263-72.
[27] Zhang H H, Huang T L, Chen S N, et al. Abundance and Diversity of Bacteria in Oxygen Minimum Drinking Water Reservoir Sediments Studied by Quantitative PCR and Pyrosequencing [J]. Microbial Ecology, 2015,69(3):618-629.
[28] Obi C C, Adebusoye S A, Ugoji E O, et al. Microbial Communities in Sediments of Lagos Lagoon, Nigeria: Elucidation of Community Structure and Potential Impacts of Contamination by Municipal and Industrial Wastes [J]. Frontiers in Microbiology, 2016,7(306):1213.
[29] 吴 鑫,奚万艳,杨 虹.太湖梅梁湾冬季浮游细菌的多样性 [J]. 生态学杂志, 2006,25(10):1196-1200.
[30] Haller L, Tonolla M, Zopfi J, et al. Composition of bacterial and archaeal communities in freshwater sediments with different contamination levels (Lake Geneva, Switzerland) [J]. Water Research, 2011,45(3):1213-1228.
[31] Saarenheimo J, Aalto S L, Rissanen A J, et al. Microbial Community Response on Wastewater Discharge in Boreal Lake Sediments [J]. Frontiers in Microbiology, 2017,8:1-12.
[32] 王 鹏,肖汉玉,张 华,等.鄱阳湖入湖河口沉积物细菌群落特征 [J]. 中国环境科学, 2018,38(4):1481-1489.
[33] 金 笑,寇文伯,于昊天,等.鄱阳湖不同区域沉积物细菌群落结构、功能变化及其与环境因子的关系 [J]. 环境科学研究, 2017,30(4): 529-536.
[34] Huang W, Chen X, Jiang X, et al. Characterization of sediment bacterial communities in plain lakes with different trophic statuses [J]. Microbiologyopen, 2017,6(5):e00503.
[35] Wu H, Li Y, Zhang J, et al. Sediment bacterial communities in a eutrophic lake influenced by multiple inflow-rivers. [J]. Environmental Science & Pollution Research, 2017:1-12.
Microbial community structure characteristics and its key driving factors in surface sediments along Nanfei River.
YANG Chang-ming1*, WU Ya-qiong1, WANG Yu-lai2, GUO Yun-hao1
(1.Key Laboratory of Yangtze Water Environment of Ministry of Education, Tongji University, Shanghai 200092, China;2.School of Energy and Environment, Anhui University of Technology, Maanshan 234000, China)., 2018,38(9):3552~3561
In order to explore the differences in bacterial community structure and its driving factors in the sediment of Nanfei River, the key physiochemical parameters including pH, CEC, TOC, DOC, and various forms of nitrogen were determined in the surface sediment samples along Nanfei River with different land uses such as urban, suburb, industrial park and rural areas. Specially, sediment bacterial community structure characteristics were analyzed by using MiSeq high-throughput sequencing technology. The relationships between sediment bacterial community structure and aforementioned physiochemical parameters were examined by redundancy analysis (RDA). The results showed that the major bacterial phylum is Proteobacteria in the surface sediments, followed by Chloroflexi, Bacteroidetes, Firmicutes. At the level of class, the major bacterial class are Anaerolineae, Gammaproteobacteria, Clostridia, Deltaproteobacteria besides the unclassified. Epsilonproteobacteria were detected mainly in sediment samples from downstream of the Nanfei River, suggesting that they were not ubiquitous in the sediment ecosystem of the Nanfei River. Most of the genus is unclassified, withandbeing predominant species. Different types of land uses along the Nanfei River led to different pollution sources and discharging load will result in different physicochemical properties of surface sediment, which in turn reshape the bacterial community structure of the sediment. The bacterial diversity in the sediments from urban and industrial areas of the middle reaches is generally higher than that in the upper and lower reaches. RDA analysis indicated that the sediment CEC, TOC, DOC, TP, nitrogen in weak acid extractable form (WAEF-N) and nitrogen in ion exchangeable form (IEF-N) were the key factors driving the differences in the structural characteristics of bacterial communities in the sediment of Nanfei River. The influence of TP was significant on Proteobacteria, Bacteroidetes and Firmicutes, and WAEF-N had a greater influence on Chloroflexi.
Nanfei River;surface sediments;bacterial communities;MiSeq high-throughput sequencing;redundancy analysis (RDA)
X522
A
1000-6923(2018)09-3552-10
杨长明(1973-),男,安徽巢湖人,研究员,博士,主要从事污染底泥和土壤生态风险评价与修复技术研究.发表论文60余篇.
2018-01-18
国家水污染控制与治理科技重大专项(2017ZX07603-003);国家自然科学基金资助项目(51608002)
* 责任作者, 研究员, cmyang@tongji.edu.cn
