过度进食肥胖症脑奖赏功能异常与运动治疗:脑功能成像证据
2018-08-13高峰焦广发董东
高峰 焦广发 董东
1河北师范大学体育学院人体运动生物信息测评省重点实验室(石家庄 050024)2河北体育学院运动人体科学系(石家庄 050041)3河北师范大学数学与信息科学学院(石家庄 050024)
遗传、饮食、行为和环境是肥胖症四大诱因。脑功能成像技术也进一步证实,进食行为、大脑奖赏功能和肥胖发生之间存在密切联系。本文旨在对过度进食者大脑奖赏功能异常的多模态脑功能成像证据进行梳理,对大脑奖赏功能与能量稳态功能在人类进食行为和肥胖发生发展中的调控作用进行评述,探讨基于脑功能成像技术开发奖赏功能异常诊断标准的可能性,为运动防肥减肥提供新的干预靶点。
1 脑功能成像技术发展
脑功能成像技术是将不同人群在从事特定任务事件中所激活的脑区以图像形式呈现,研究结构与功能关系的神经成像技术。目前用于人类进食行为研究的脑功能成像技术有:(1)基于电磁学理论的脑电图、相关事件电位和脑磁图,可检测大脑对诱发刺激响应的电或磁信号,其对脑功能活动区判定的时间分辨率高,但空间分辨率较低;(2)基于血流动力学的功能磁共振成像(functional magnetic resonance imaging,fMRI)、正电子发射断层扫描、单一正电子发射计算机断层扫描和近红外脑功能成像(functional near infrared spectroscopy,fNIRS)可检测“脑-进食线索”间奖赏信息的加工过程,具有良好的空间分辨率,但时间分辨率不及脑电图[1]。其中fNIRs技术和fMRI技术具有共同的神经生理学基础即神经血管耦合机制。神经血管耦合机制是指当大脑受到线索刺激或执行一项任务时,处于激活状态的相应脑区局部血流量和氧代谢率会显著增加,氧合血红蛋白和脱氧血红蛋白发生相应的改变。fMRI是利用核磁共振造影测量神经元兴奋引起的脑血流改变,通过脑血氧水平依赖指标推测脑皮层功能状态和皮层下如腹侧纹状体、苍白球、丘脑和下丘脑等信号加工过程。而fNIRS主要依赖于近红外光与脑组织中生色物质之间的吸收和散射关系,根据Beer-Lambert定律推算激活脑区血氧和血容量的变化,从而反映与进食行为密切相关的脑皮层区血流动力学变化,如前额叶皮层、眶额叶皮层、颞叶皮层和额颞叶等。
目前,借助不同脑功能成像技术可确定不同人群进食行为(食物偏好、进食频率和食物能量密度等)与特定脑功能状态的关联[2],针对这种结构与功能的关联采用统计技术(区域间相关、因素分析、动态因果建模等)获取进食行为相关的大脑奖赏功能激活模式,进而对过度进食肥胖症的发生、发展和转归进行预测和评价,则是目前的发展方向。
2 进食行为、肥胖与脑奖赏功能异常
目前学术界公认的肥胖与进食行为关系的两大理论是“自体能量调节稳态失衡”和“大脑奖赏功能异常”理论[3]。自体能量调节稳态失衡理论认为,机体对能量储备不足的调节能力要远大于对能量过剩的调节能力,这种不对称性稳态调节易导致在美味食物丰富环境下的过度进食行为发生,促进肥胖的发生与发展。大脑奖赏功能异常理论认为,进食行为受到双信号通路调控,即内环境能量信号通路和外环境食物线索信号通路。食物线索刺激包括口味、颜色、形状、气味、能量密度等,食物线索刺激和进食后的体验(愉快/厌恶)促进人类在出生后逐步形成稳定的大脑食物奖赏调节信号系统[4]。美味可口的食物所带来的愉悦感是促使个体非饥饿或饱腹后过度进食强有力的内驱力。大脑奖赏信号系统在食物线索触发-进食行为-中枢愉快奖赏的刺激与强化的习服过程中,会出现奖赏过度现象,即大脑对食物线索产生奖赏期待过高或超敏感,导致正常进食引起的愉快感奖赏不能与预期相匹配,而出现进食过度的现象[5]。目前研究证实,大脑奖赏功能异常是奖赏期望(眶额叶、岛叶和扣带回)、学习记忆(海马、杏仁、纹状体和丘脑)、奖赏权衡(伏隔核和腹侧被盖区)和决策执行(前额叶)这4个脑功能区/核团间相互作用的结果[6]。
进食行为、进食奖赏和能量稳态是肥胖防治的3个关键调节靶点(见图1)。进食行为是生理性需求和心理性渴望驱动的人类行为。进食行为的生理需求满足是通过能量稳态调节功能实现的,而进食行为的心理需求满足则通过奖赏功能获取,可见人类进食行为受到双重信号调控[7]。肥胖的形成过程中多伴有进食行为的改变,这与大脑奖赏系统调节异常有关,但两者间的因果关系尚未探明。是丰富食物环境引发进食行为改变,进而大脑奖赏功能失常引发过度进食行为,还是肥胖引发大脑奖赏功能异常进而引发进食过度?这分别涉及学习记忆的正强化机制,大脑奖赏系统神经信号通路突触的可塑性变化机制,以及奖赏和冲动性进食的神经心理学机制,目前尚处于学术争鸣阶段。

图1 大脑奖赏功能和能量稳态功能在机体进食行为和肥胖发生中的双信号调控[6,7]
3 过度进食肥胖症脑奖赏功能异常:脑功能成像证据
过度进食肥胖症的临床判定主要借助主观测量法(食物渴望度和饱腹感的问卷测量)和客观测量法(进食量的测量和脑奖赏功能神经活性的脑成像测量)。研究显示过度进食肥胖症虽具共同的行为学特征,但是脑奖赏功能异常的表现形式却不尽相同。脑功能成像技术是研究进食行为相关的大脑奖赏脑区结构与功能的激活、多巴胺受体功能和多巴胺基因多态性的有效检测手段[8],近些年来基于脑功能成像技术的过度进食肥胖症成因与分型研究显示,过度进食肥胖症按成因可分为脑奖赏超敏感型、脑奖赏缺失型和认知反馈抑制受损型。而如何快速准确判定过度进食肥胖症成因,并依据不同肥胖亚型来制定相应防治方案,则是肥胖症治疗的瓶颈。
3.1 脑奖赏超敏感的脑功能成像证据
脑奖赏超敏感的成因理论认为,由于食物是包含进食前线索认知刺激和进食后代谢信号刺激的双重外部刺激物,美味可口食物的反复摄入可促成食物线索与可口食物间的脑内关联形成,进而造成大脑奖赏功能区对食物线索信号产生过度神经活化反应,即大脑奖赏超敏感或奖赏预期过高,诱发过度进食肥胖症的形成[9]。脑功能成像的横向比较研究证据一致显示,大脑的奖赏区对食物线索反应虽具个体差异,但是成人和青少年过度进食肥胖症患者与健康体质者相比,奖赏功能区(腹侧纹状体、杏仁核和前额叶皮质等)均表现出更高食物线索敏感,这是造成其心理性奖赏期望过高的神经生理学基础[10,11]。近些年几项前瞻性研究进一步围绕脑奖赏功能异常是过度进食的前因还是后果展开研究,美食图片、食物广告、食物形状和食物的风味均可激活奖赏相关的脑区(见表1)。这些证据提示,检测多巴胺能奖赏系统(伏隔核、杏仁核和眶额叶皮质)对食物线索的神经功能反应特征,是预测个体过度进食肥胖症发生和评价治疗效果的有效方法。2016年一项荟萃分析研究表明,少年儿童的脑奖赏功能与成年人相比存在显著差异[12],提示饮食行为对脑奖赏功能具有可塑性影响。

表1 脑奖赏功能对食物线索超敏性的前瞻性脑功能成像证据
3.2 脑奖赏缺失的脑成像证据
脑奖赏缺失型肥胖的成因理论认为,脑内基于多巴胺的脑奖赏环路的合成/分泌障碍会造成脑内奖赏值下降,因此机体通过过度进食以弥补降低的奖赏值,进而诱发过度进食肥胖症[12]。借助PET、SPECT和fMRI技术,探究多巴胺系统在进食行为中的作用,结果显示肥胖程度与多巴胺信号释放量呈负相关,即随着体脂肪率或体重身高指数(BMI)增加,背侧纹状体多巴胺合成水平降低[17];肥胖还引发纹状体多巴胺 D2/D3受体结合率较低[18]。有研究还显示肥胖引发脑内μ-阿片受体减少,但多巴胺 D2受体利用率未改变,推测除多巴胺启动的奖赏调节环路外还有其它神经环路参与大脑摄食奖赏[19]。但一项动物研究发现,长期高脂饮食喂养降低小鼠脑内多巴胺奖赏环路对胃内脂质信号油酰乙醇胺(OEA)的反应,导致低脂饮食难以使小鼠获得奖赏满足,进而形成高脂饮食偏好[20]。但是,奖赏缺失与肥胖之间不是简单的因果关系,“脑奖赏缺失型肥胖"是环境、饮食、情绪和行为交互作用的结果,奖赏缺失只是其中的一个环节。
3.3 认知反馈抑制受损的脑功能成像证据
认知反馈抑制受损的肥胖成因理论则认为,在非生理性饥饿下,长期处于美味食物环境下而诱发的过度进食,会使脑内认知控制功能异常,主要是前额叶和顶叶,造成摄食行为反馈抑制受损引发过度进食肥胖症[21]。目前该理论成因的前瞻性研究证据尚不充分。Batterink等[22]关于认知反馈抑制与BMI关系的fMRI证据研究显示,超重肥胖青少年对美食线索具有更强的进食冲动,但是其认知反馈抑制区(眶额叶、前额叶皮质内侧、额上回、额中回)神经兴奋性则低于体重偏低者,且腹内侧前额叶对美食线索的低激活与1年后BMI增加呈反比。另一项关于脑执行功能对体重增加的预测作用的前瞻性fMRI证据研究显示,肥胖者大脑额前回、额中回和额下回对不同难度的延迟折扣刺激任务的低激活应答,可以预测其未来1.3~2.9年间体重呈增加趋势[23]。
但是,一项基于认知干预法人随机对照研究也显示,通过脑认知控制干预提高肥胖者前额叶皮质对高脂膳食的反应,以抵抗脑内奖赏区对食物线索的超敏性,可以短暂减少过度进食,但长期减肥效果不佳。研究提示,奖赏认知抑制行为干预可对进食行为产生抑制,但是肥胖者脑内奖赏区对食物线索的超敏一旦形成,难以单纯依靠认知干预恢复其正常的大脑奖赏功能[24]。从人类进化角度看,人类的能量稳态调节功能在面对丰富美食的环境时,反馈抑制的调节能力弱化[25]。
4 过度进食肥胖症运动治疗与相关脑成像证据
运动与食欲和进食行为之间的关系也一直是过度进食肥胖症运动治疗领域的研究焦点。进食和运动是决定人体能量平衡的两个即相互独立又相互关联的行为变量。从理论上看,保持机体趋向负能量稳态平衡是降低体重的关键。运动锻炼和饮食限制是最常用的诱导能量缺失的手段。20世纪90年代至今运动对人体食欲和运动后能量摄入影响的相关研究充分说明,急性运动引发的能量消耗(energy expenditure,EE)和运动后能量补偿(energy compensatory,EC)之间存在微弱关联,大量针对健康人群、有训练经历者和肥胖人群的实验研究均支持,虽然运动增加能耗的同时也会提高个体对食物的渴望[25],但是从中等强度、大强度到大强度间歇急性运动均不会引发个体运动后能量补偿增加[26-29]。从神经内分泌视角探究运动、食欲和能量稳态调控关系的研究也证实,食欲相关的脑肠激素在保持运动后即刻负能量稳态调节中起到关键作用[30-32]。健康成年男子在承受不同诱因但相近能量缺失时(饮食限制诱导能量缺失 vs.运动诱导能量缺失)会有不同的EC变化,饮食限制诱导能量缺失增加EC而运动诱导能量缺失则没有增加EC。而这与运动诱导的能量缺失可降低血浆促食欲激素ghrelin和增加抑食欲肽PYY(3-36)水平密切相关[33]。这说明食欲相关的脑肠激素对于胃肠内能量摄入和利用的即刻反应很敏感,但对运动引起的能量负平衡则反应迟缓至运动后数小时[32]。肥胖者与体重正常者具有相一致的急性运动后食欲相关的激素反应,肥胖者急性运动后40 min~2 h内均比体重正常组更显著呈现能量摄入下降[34.35]。研究表明急性运动降低促食欲素分泌主要与运动造成血流再分配密切相关,而其增加抑食欲素分泌则受到交感神经兴奋性、胃肠蠕动、细胞因子释放和糖脂代谢间接调控[36-38]。但是对运动减肥的长期效果研究显示,长期有氧运动对肥胖者减体重效果具有很大的个体差异[39],急性运动抑制食欲作用尚不能作为长期体重控制效果的有效预测因子[40]。而肥胖者12周有氧运动减脂效果不佳与运动增强了其对食物心理奖赏的渴望有关,具有高脂饮食偏好的肥胖者在运动后即刻对食物的喜爱程度和对高脂甜食的需求动机均显著提高,这种稳定的饮食偏好的心理特征不会随12周规律运动引起的体重下降而弱化[17]。Alkahtani等[41]进一步研究还发现,4周中等强度间歇运动可增加肥胖者对高脂无糖食物的喜爱程度,而大强度间歇运动(90%VO2max 30 sec+间歇30 sec)则会降低这一偏好。大强度运动以糖供能为主,而中等强度虽然能增加运动中的脂代谢供能比例,但是由于肥胖者脂肪储备为过剩状态,及其在运动态和静息态的脂肪利用率远低于正常体重者,因此仅从能量代谢和神经内分泌角度很难全面解释运动对肥胖者对食欲抑制的强度依赖性和食物喜爱偏好的影响机制,还需从神经认知角度关注大脑奖赏功能对肥胖者运动后进食补偿的认知调控作用。
近10年来,随着脑成像测量评价技术的迅猛发展,大量脑功能成像证据显示,肥胖者不但具有对食物线索刺激的脑内奖赏功能异常激化(主要岛叶和眶额叶皮质),而且在没有食物奖赏信号刺激下,肥胖者脑奖赏功能相关脑区的功能连接(额叶、岛叶、扣带回、海马、杏仁核、纹状体、伏隔核和丘脑等)也表现出随着体脂量的增加而发生异常且伴有高能食物偏好的过度进食行为[42]。基于“运动对肥胖者食欲和能量稳态影响”的相关研究充分说明,急性运动可降低肥胖者食欲和诱导能量负平衡,但是运动的长效机制则具有很大的个体差异,可见长期运动后的能量负稳态变化受到个体对食物奖赏认知调控的影响要远高于能量代谢的生理调控作用[26,39]。
而关于运动是否可对脑进食奖赏功能产生有益于体重控制的影响这一问题,两项基于fMRI脑功能成像技术的研究给出了肯定回答。Evero等[43]研究显示,经常进行运动锻炼的成年人在完成60分钟大强度功率车运动后即刻(83% ±1.0%HRmax),大脑对食物奖赏期待值(左右岛叶、右侧尾壳核和右侧罗兰帝克岛盖)显著降低,食物线索的视觉注意(枕中回和枕下回)显著下降,且奖赏认知抑制脑区(眶额叶)对食物线索的神经反应增强。这项研究首先证实,运动不但对能量稳态调节相关的食欲激素表达具有调控作用,而且急性运动对健康体重者的奖赏刺激动机认知和奖赏信息加工也具有有益作用,这提示持久的运动锻炼有助于在丰富美食环境下保持脑奖赏功能稳态,这可能是长期运动控制体重的神经认知机制之一。Cornier等[44]进一步检验中等强度急性运动和24周运动训练对肥胖者食物信号脑神经反应和进食补偿的影响,结果显示急性和长期中等强度有氧运动可以显著降低肥胖者大脑岛叶(为脑奖赏期待功能区)对食物刺激的神经兴奋性,岛叶兴奋变化与个体体脂变化成正相关,而与体重变化无显著关联。这项研究进一步证实,长期中等强度有氧运动改善肥胖者过度进食行为与其具有降低肥胖者大脑岛叶对食物奖赏期待异常激活密切相关。现有的大脑奖赏功能异常与肥胖症关系的脑成像证据已证实过度进食肥胖症具有进食行为异常(高热量偏好、享乐和压力型等)和脑奖赏功能异常(食物线索神经敏感性升高、多巴胺信号调节下降和认知反馈神经兴奋性下降等)[45],而运动对肥胖者脑奖赏功能可塑性影响的脑功能成像证据(表2)也充分支持运动可对过度进食肥胖症脑奖赏功能异常产生有益影响。相关研究还显示,成功减重肥胖者在运动干预前后的主观进食渴望要显著低于运动减重抵抗的肥胖者[17],且成功体重控制者比减重后反弹者具有高的脑内进食奖赏的自我认知控制[27]。运动对非稳态的奖赏性进食行为和药物成瘾行为影响的脑成像研究还显示,运动对奖赏系统多巴胺通路的作用可能与物质成瘾具有共同的奖赏通路[3],过度进食和运动成瘾可触发相似的奖赏通路,运动有可能替代食物刺激成为外源性奖赏刺激,起到减弱或消除其他成瘾行为的作用[46]。
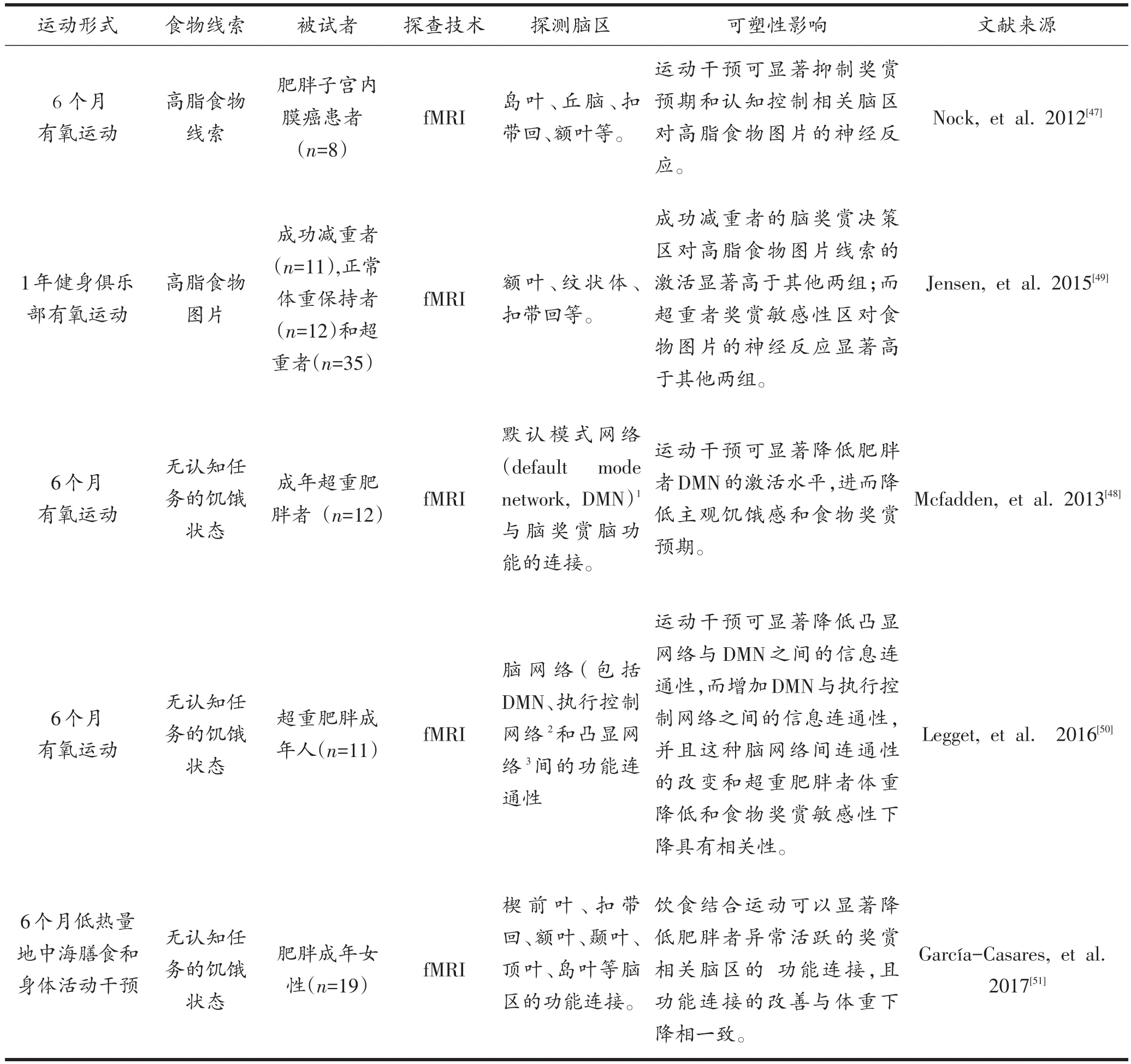
表2 运动对肥胖者脑奖赏功能可塑性影响的脑功能成像证据
5 总结
综上分析,长期的过度进食和静态生活方式相伴的脂肪积累可对肥胖者大脑的奖赏区产生结构和功能的可塑性影响[10,11,22,23,36,],而长期运动也可对肥胖者异常的奖赏期望(眶额叶、岛叶和扣带回)和决策执行(前额叶)结构和功能联结产生有益的可塑性[47-52]。由于受到fMRI和ERP等脑成像检测技术测试条件的限制,目前运动对过度进食肥胖症脑奖赏功能异常可塑性影响的研究才刚刚起步,现有的脑功能成像研究主要围绕运动后肥胖者对食物信号加工的功能变化展开,尚无法对运动过程中和自然身体活动状态下的肥胖者脑奖赏功能的动态变化进行探究。但是2017年1项对417名成年人为期1年的纵向描述性研究显示[53],成年人运动量与主观享乐性食欲感知之间的相关性受到锻炼形式和性别影响,而长期运动对男性主观食欲控制效果更加明显。因此,进一步探究自然身体活动状态下不同性别、不同饮食偏好的过度进食肥胖者对不同运动强度、形式和运动量干预后的脑奖赏功能应答的时相性证据显得更为迫切。
fNIRS技术是近些年发展起来的一种有效探查食物线索刺激对大脑皮层功能区影响的脑功能成像技术,可用于额叶对美食线索的认知研究[54],以及额叶和颞叶皮层对不同食物信号刺激(食物颜色、气味、风味、食物营养成分、食物形状和美味可口图片)激活范式的研究[55]。近些年来,随着fNIRS信号分析技术发展,其对摄食的神经行为学研究的补充作用日益显著。2016年有研究报道显示,fNIRS监测中运动伪差信号去噪能力进一步提高,可实现在功率车运动状态下实时监测前额叶区血氧动力学变化[56],且fNIRS与fMRI空间分辨率的测量误差仅为4.4±1 mm,低于脑回与脑沟之间平均间距[57]。因此,fNIRs技术是一种在静态和运动(步行、骑行和跑步)状态下,探查环境线索(食物和运动等)刺激对大脑皮层功能区影响的有效手段。基于fNIRs的研究结果具有较高生态效度,同时还可通过图像数据处理技术获得实时光学脑地图(topography)[16],这为今后借助fNIRS的脑成像技术,揭示自然身体运动下奖赏渴望驱动的过度进食者大脑皮层奖赏功能异常的运动反应特征,筛选过度进食肥胖症的神经认知学检测指标,为运动治疗效果反馈提供了新的研究范式。
