17-AAG对人绒毛膜癌JAR细胞MMP-2和MMP-9表达的调控作用*
2018-05-09崔丽军刘百艺安国庆
谷 苗,崔丽军,刘百艺,安国庆,许 倩
(承德医学院,河北承德 067000)
人绒毛膜癌是一种来源于滋养细胞的恶性肿瘤,由于滋养细胞具有胚胎细胞的特性,导致绒癌早期即发生血道转移,波及全身,危及患者生命。热休克蛋白90(HSP90)是许多癌基因通路中的重要组成部分,因此HSP90是肿瘤治疗中非常有前景的分子靶点[1]。17-丙烯胺基-17-去甲氧基格尔德霉素(17-AAG)作为HSP90的抑制剂在骨肉瘤[2]、胆囊癌[3]、肺癌[4]及乳腺癌[5]等的研究中具有抑制肿瘤细胞生长、诱导肿瘤细胞凋亡的作用,但关于17-AAG对人绒毛膜癌JAR细胞的抗癌作用国内外鲜有报道。本研究以人绒癌JAR细胞为研究对象,观察了
17-AAG对人绒毛膜癌JAR细胞基质金属蛋白酶2(MMP-2)、MMP-9蛋白表达的调控作用,初步探寻17-AAG对JAR细胞侵袭、转移的干预作用,为17-AAG对人绒毛膜癌治疗作用的研究提供实验依据。
1 材料与方法
1.1 材料与主要仪器 人绒毛膜癌JAR细胞系(广州吉尼欧生物科技有限公司),胎牛血清(FBS)、DMEM培养基(美国Gibco公司);17-AAG(深圳生尔易美有限公司);鼠抗MMP-2单克隆抗体、兔抗MMP-9单克隆抗体、兔抗GAPDH单克隆抗体(美国Abcam公司);酶标仪(美国BioTek Cytation5)。
1.2 细胞培养 人绒毛膜癌JAR细胞用含15% FBS的DMEM培养基,置于37℃、5% CO2培养箱中培养,每2-3天更换培养基,取对数生长期的细胞进行后续实验。
1.3 MTT法检测绒癌JAR细胞的增殖抑制率 取细胞状态良好、对数生长期的JAR细胞,胰酶消化计数,按5×103/ml接种于96孔板中,设置对照组、实验组、空白组。待细胞长到密度为90%左右时,对照组细胞不加17-AAG,实验组细胞加入不同浓度的17-AAG,分别为5μg/ml、10μg/ml、20μg/ml、40μg/ml,每组设5个复孔。培养48h后,加入MTT(5μg/ml),每孔10μl,继续培养4h,吸掉培养液,每孔加入二甲基亚砜150μl,震荡15min,酶标仪于波长490nm处检测吸光度。计算JAR细胞的增殖抑制率。增殖抑制率=(1-实验组OD值/对照组OD值)×100%,并计算药物半数抑制浓度(IC50)。
1.4 Western blotting法检测绒癌JAR细胞MMP-2、MMP-9蛋白的表达 取细胞状态良好、对数生长期的JAR细胞接种于6孔板中,待细胞长到密度为90%左右时,分别加入0μg/ml(对照组)、5μg/ml、10μg/ml、20μg/ml、40μg/ml浓度的17-AAG。继续培养24h后,加入RIPA蛋白裂解液提取各组细胞的总蛋白,并检测蛋白浓度。60μg蛋白上样量进行12% SDS-聚丙烯酰胺凝胶电泳,采用PVDF膜进行转膜,5%脱脂奶粉封闭1h,加入5%脱脂奶粉稀释的一抗MMP-2(1:2000)、MMP-9(1:2000)4℃孵育过夜,GAPDH(1:4000)作为内参。然后加入5%脱脂奶粉稀释的二抗(1:1000)室温孵育2h,超敏发光液显影后使用Tanon5500全自动化学发光成像分析系统显影,并用Quantity One软件进行灰度分析,以目的蛋白条带与内参蛋白条带灰度的比值作为目的蛋白的相对表达水平。实验重复3次。
1.5 统计分析 采用SPSS 19.0软件进行统计分析,多组间计量资料的比较采用单因素方差分析法,P<0.05为差异有统计学意义。
2 结果
2.1 不同浓度17-AAG对人绒毛膜癌JAR细胞增殖的影响MTT结果显示,17-AAG对人绒毛膜癌JAR细胞生长具有明显的抑制作用,随着17-AAG浓度的升高,JAR细胞的增殖抑制率逐渐升高(P<0.05,图1)。药物作用48h细胞增殖抑制率IC50均值为5.736。
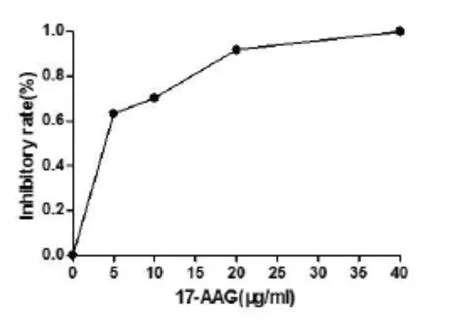
图1 不同浓度17-AAG对JAR细胞的抑制作用
2.2 不同浓度17-AAG对JAR细胞MMP-2和MMP-9蛋白表达的影响 分别用5μg/ml、10μg/ml、20μg/ml、40μg/ml的17-AAG处理细胞24h,与对照组(0μg/ml)相比,各实验组细胞MMP-2和MMP-9蛋白的表达明显降低,随着17-AAG浓度的升高,MMP-2和MMP-9蛋白的表达逐渐降低(P<0.05,图2)。
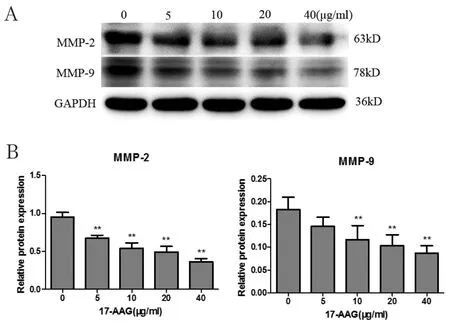
图2 不同浓度17-AAG对JAR细胞MMP-2、MMP-9蛋白表达的影响
3 讨论
基质金属蛋白酶(matrix metalloproteinase,MMP)的主要功能是降解细胞外基质、维持细胞外基质的平衡,且在肿瘤细胞的生长、分化、侵袭、转移等过程中起重要作用[6]。MMP-2和MMP-9属于基质金属蛋白酶家族,与多种肿瘤的发生发展有关。有研究表明[7],肽酰精氨酸脱亚氨酶1(PAD1)抑制剂通过调节MEK1-ERK1/2-MMP-2信号通路发挥抗人乳腺癌的作用;Liu[8]等研究发现,姜黄素通过激活miRNA-98抑制MMP-2和MMP-9的表达和活性,从而控制肺癌的侵袭和转移;Cheng等[9]研究发现,短指软珊瑚内酯能通过PI3K/AKT/mTOR信号通路下调MMP-2/-9和尿激酶的表达,从而抑制人膀胱癌细胞的侵袭和迁移;另外,番石榴叶的提取物丁醇成分通过抑制ERK1/2-MAPK信号通路抑制MMP-2和MMP-9的表达,从而抑制肺癌细胞的侵袭和转移[10]。可见,MMP-2和MMP-9在多种肿瘤的侵袭和转移中发挥重要作用。为此本研究观察了17-AAG对人绒毛膜癌JAR细胞MMP-2和MMP-9蛋白表达的调控作用,以分析17-AAG对JAR细胞的干预作用及机制。
本研究结果显示,17-AAG对人绒毛膜癌JAR细胞的生长具有明显的抑制作用,且呈浓度依赖性;同时,17-AAG还可以下调JAR细胞MMP-2和MMP-9的表达,提示17-AAG可能通过降低MMP-2和MMP-9的表达水平,从而抑制JAR细胞的侵袭和转移。由于抗肿瘤细胞的侵袭和转移是一个多因素参与的复杂体系,17-AAG是通过调节MEK1-ERK1/2-MMP-2信号通路还是PI3K/AKT/mTOR信号通路对MMP-2和MMP-9进行调控,还需进一步的研究探讨。
【参考文献】
[1] Oh YJ, Seo YH. A novel chalcone-based molecule, BDP inhibits MDA-MB-231 triple-negative breast cancer cell growth by suppressing Hsp90 function[J]. Oncol Rep, 2017, 38(4): 2343-2350.
[2] Massimini M, Palmieri C, De Maria R, et al. 17-AAG and Apoptosis,Autophagy, and Mitophagy in Canine Osteosarcoma Cell Lines[J].Vet Pathol, 2017, 54(3): 405-412.
[3] Weber H, Valbuena JR, Barbhuiya MA, et al. Small molecule inhibitor screening identified HSP90 inhibitor 17-AAG as potential therapeutic agent for gallbladder cancer[J]. Oncotarget,2017, 8(16): 26169-26184.
[4] Woo JK, Jang JE, Kang JH, et al. Lectin, Galactoside-Binding Soluble 3 Binding Protein Promotes 17-N- Allylamino-17-demethoxygeldanamycin Resistance through PI3K/Akt Pathway in Lung Cancer Cell Line[J]. Mol Cancer Ther, 2017, 16(7): 1355-1365.
[5] Raja SM, Clubb RJ, Bhattacharyya M, et al. A combination of Trastuzumab and 17-AAG induces enhanced ubiquitinylation and lysosomal pathway-dependent ErbB2 degradation and cytotoxicity in ErbB2- overexpressing breast cancer cells[J].Cancer Biol Ther, 2008, 7(10): 1630-1640.
[6] Lubbe WJ, Zhou ZY, Fu W, et al. Tumor epithelial cell matrix metalloproteinase 9 is a target for antimetastatic therapy in colorectal cancer[J]. Clin Cancer Res, 2006, 12(6): 1876-1882.
[7] Qin H, Liu X, Li F, et al. PAD1 promotes epithelial-mesenchymal transition and metastasis in triple-negative breast cancer cells by regulating MEK1-ERK1/2-MMP2 signaling[J]. Cancer Lett,2017, 409: 30-41.
[8] Liu WL, Chang JM, Chong IW, et al. Curcumin Inhibits LIN-28A through the Activation of miRNA-98 in the Lung Cancer Cell Line A549[J]. Molecules, 2017, 22(6). pii: E929.
[9] Cheng TC, Din ZH, Su JH, et al. Sinulariolide Suppresses Cell Migration and Invasion by Inhibiting Matrix Metalloproteinase-2/-9 and Urokinase through the PI3K/AKT/mTOR Signaling Pathway in Human Bladder Cancer Cells[J]. Mar Drugs, 2017,15(8). pii: E238.
[10] Im I, Park KR, Kim SM,et al. The butanol fraction of guava(Psidium cattleianum Sabine) leaf extract suppresses MMP-2 and MMP-9 expression and activity through the suppression of the ERK1/2 MAPK signaling pathway[J]. Nutr Cancer, 2012, 64(2):255-266.
