不同居群驼蹄瓣叶片解剖结构比较研究
2018-04-23任尚福
任尚福
(喀什大学 研究生处,新疆 喀什 844008)
南疆是指新疆天山以南,昆仑山以北的广大地区,地处干旱的荒漠地带。终年降雨稀少,蒸发强烈,水资源时空分配不均,植被稀疏,生境条件严酷,生态系统十分脆弱,荒漠占绝对优势,因而生活在南疆地区的植物是南疆绿洲的防线,是绿洲与沙漠之间的绿色生态屏障。在南疆分布的植物在长期与恶劣环境的斗争中,逐渐在结构上及生理上形成了适应特征。同时近年由于环境破坏日趋严重,土壤盐渍化成为当今世界困扰人类的重大问题之一,这更加剧了南疆生态的的脆弱性。
驼蹄瓣(ZygophyllumfabagoL.)在分类上属于蒺藜科霸王属多年草本植物[1]。驼蹄瓣生于荒漠、草原化荒漠及荒漠化草原带,属温带干旱地区的强旱生或旱生植物,是长期适应自然干旱环境的产物,具有耐干旱、抗盐碱、耐贫瘠、耐风蚀沙埋等优良性状,有较强的生态适应能力,作为南疆荒漠区系灌木片层中主要优势种和建群种,对维持南疆脆弱的生态系统具有及其重要的作用。近年来,对旱生植物的解剖学研究主要集中在对不同生态型或亲缘植物形态结构的比较研究,植物对于干旱环境采取种种适应对策,但并非每种植物都采用其中一种对策,而是有不少植物同时采用多种对策。在植物的结构与环境关系方面,历来研究最多的器官是叶,因为叶片是植物进化过程中对环境变化比较敏感且可塑性较大的器官,环境变化常导致叶片的形态及解剖结构的响应与适应[2-4]。目前,有关驼蹄瓣的研究相对较少,主要集中于化学成分、属内亲缘关系和遗传多样性、种子萌发条件研究等方面[5-7]。本文通过采集3个不同居群不同生境的驼蹄瓣的植物叶片进行比较解剖学研究,试图通过对其叶片解剖结构的比较研究,探讨驼蹄瓣叶的解剖结构对不同环境的适应性,为南疆大片盐碱化土地的改良和脆弱的生态环境改善提供一定的理论依据和参考。
1 材料与方法
1.1 研究区概况
南疆属典型的大陆性干旱气候,昼夜温差大、干旱、光照长、降水量少、蒸发强烈,年平均气温10~13 ℃,无霜期200~220 d。试验选取阿克陶县、塔什库尔干县、沙雅县作为研究区(各研究区域具体气候特征见表1)。
1.2 实验材料
在每个研究区内,选择3个3 m×3 m的样方,在每个样方内随机选取3株生长健壮、长势一致的植株(实验材料采集具体地点见表2)。在每颗植株中部取发育程度大致相同的一片正常生长叶片,用锋利的刀片从叶片中脉及周围部位切取3~5 mm的叶片组织,立刻放在FAA固定液中封存编号,并用注射器抽取其中的空气。
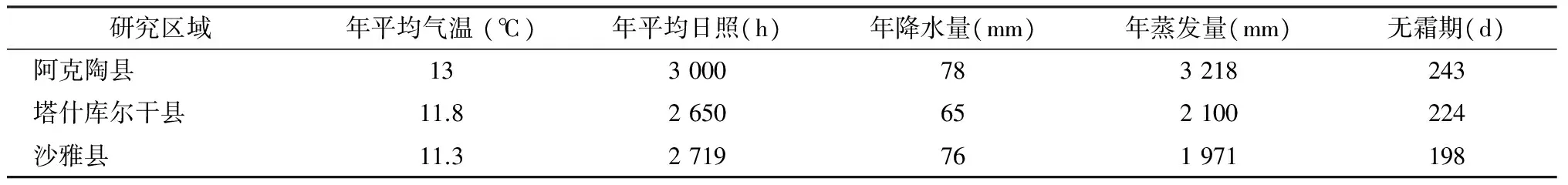
表1 各研究区域基本气候特征

表2 叶片采集地点
1.3 试验方法
每个处理选取3个材料,采用石蜡切片法制作切片,每个切片选5个观察视野,用DMB5-223I-5图像处理显微镜照相,并用Arcview3.3软件测定叶片厚度、上角质层厚度、上表皮厚度、栅栏组织厚度、海绵组织厚度、下表皮厚度及下角质层厚度,并计算出叶片栅海比值,栅海比值=栅栏组织厚度/海绵组织厚度。
1.4 数据分析
数据分析采用SPSS 17.0软件处理,对不同居群间驼蹄瓣叶片解剖结构特征比较在P<0.05水平上进行单因素方差分析,用最小显著差法(LSD)检验各平均值间的差异显著性,并采用Excel 2010绘制图表。
2 结果与分析
2.1 驼蹄瓣叶片解剖结构特征
驼蹄瓣叶片解剖结构由表皮、栅栏组织、海绵组织、叶脉等构成,其解剖结构整体呈现对称形分布,为等面叶(见图1)。表皮具有1层生活细胞,由表皮细胞和气孔器组成,表皮细胞排列紧密,不含叶绿体,具有保护内部的组织结构和贮存水分的作用;细胞外壁上有蜡质透光比较厚的角质层;叶肉具有栅栏组织和海绵组织分化,栅栏组织细胞成圆柱型,细胞长轴与叶表面垂直,含有大量的叶绿体,海绵组织细胞不规则,排列不紧密,细胞之间具有较多间隙。
2.2 不同居群驼蹄瓣叶片厚度特征
驼蹄瓣叶片厚度普遍比较厚,但不同居群叶片厚度差异性较大(见表3),T-1叶片厚度最厚达到530.986 μm,T-2为463.586 μm,T-3叶片厚度最薄为409.914 μm。方差分析表明,T-1叶片与T-2叶片和T-3叶片厚度差异性均显著,T-2叶片和T-3叶片厚度差异性显著。

表3 不同居群驼蹄瓣叶片解剖结构数据统计
2.3 不同居群驼蹄瓣叶片解剖表皮特征
驼蹄瓣叶片表皮细胞外壁具角质层,由单层长方形细胞构成,细胞大小不一,排列紧密,角质层较厚,上表皮细胞较下表皮细胞形状大,排列更为紧密。上表皮厚度大于下表皮厚度,相差比较小,T-1叶片的表皮层和角质层最厚,其上表皮、下表皮、上角质层和下角质层最大分别为:22.908 μm、17.41 μm、2.612 μm和17.41 μm。方差分析表明:T-1叶片上角质层厚度、上表皮厚度及下表皮厚度与T-2叶片和T-3叶片上角质层厚度、上表皮厚度及下表皮厚度均有明显差异,T-2和T-3上角质层厚度及下角质层厚度之间差异不显著。


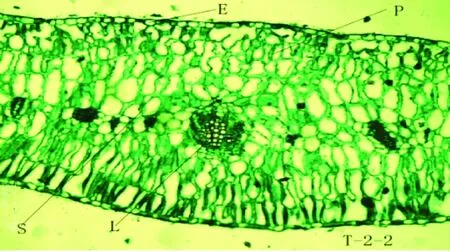

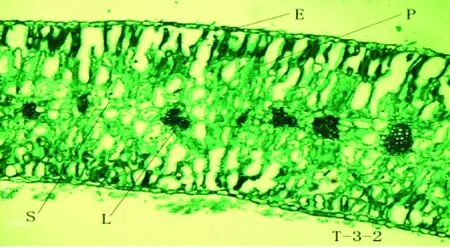
T-1-1、T-1-2驼蹄瓣-1;T-2-1、T-2-1驼蹄瓣-2;T-3-1、T-3-1驼蹄瓣-3;T-1-1、T-2-1、T-3-1 (10×10);T-1-2、T-2-2、T-3-2 (10×40)
2.4 不同居群驼蹄瓣叶片解剖叶肉特征
驼蹄瓣叶片中的叶肉分化为栅栏组织和海绵组织,栅栏组织位于上表皮下方,由一层柱状排列紧密的细胞组成,靠近上下表皮细胞均有栅栏组织的分化,且上下层栅栏组织厚度差异不大,海绵组织分布在栅栏组织之间,且不同居群之间海绵组织发达程度不同。栅栏组织和海绵组织厚度在不同居群之间表现出一定的差异性。T-1叶片的栅栏组织最厚为369.122μm,海绵组织厚度为165.318 μm。方差分析表明:T-1与T-2、T-3叶片的栅栏组织厚度差异显著,T-2及T-3叶片的栅栏组织厚度差异显著,T-1与T-2叶片的海绵组织厚度显著差异,T-2及T-3叶片的海绵组织厚度差异不显著。在这三个居群中,T-1叶片的栅海比值最大为2.233,T-2叶片次之为2.176,T-3叶片的栅海比值最小为2.003。
3 结论与讨论
因叶片是植物暴露生长环境中面积最大的器官,所以在植物进化过程中其形态解剖结构易受海拔、温度和降雨量等环境因子的影响,表现出明显的形态差异,并呈现一定的变化机制,形成了特定的形态特征,可为属内种间的分类提供重要依据[8],也可用于评价植株的抗逆性[9-10]。植物器官的形态结构是与其生理功能和生长环境密切相适应的,在长期外界生态因素的影响下,叶在形态结构上的变异性和可塑性是最大的,即叶对生态条件的反映最为明显。通过对不同居群驼蹄瓣叶片解剖结构的观察得出,其叶片由表皮、叶肉和叶脉三部分组成,解剖结构整体呈基本对称形分布,为等面叶。植物的表现性状主要是有遗传物质决定的,但环境因素可以使得遗传因素得以加强或减弱。南疆地区四面环山,地形封闭属暖温带大陆性干旱气候,其特点是干旱、光照长、少雨,蒸发强烈,并且南疆土壤具有高盐碱、高含砂量、土壤贫瘠、土层较薄等一系列特征。驼蹄瓣有较厚的叶片厚度,较厚的角质层和表皮层,这都与其生活在干旱的环境有关。本实验中不同居群的驼蹄瓣叶片解剖特征指数之间具有显著性差异,这与刘世彪提出的叶片是植物营养器官中对环境变化最为敏感的器官是一致的[11]。张红霞研究浑善达克沙地榆指出同一种植物由于它们长期处于不同的生境条件下,其叶片的解剖结构也相应产生了差异[12]。陈庆诚研究指出生长在环境条件差异程度很大的同一种植物,它们在形态与解剖结构上,常表现了明显的趋异现象,生长在不同生境条件下的植物通过各种改变来适宜环境[13]。不同地区由于气候特点、地理环境、土壤条件和生物因素等不同,在长期的进化过程使不同生态环境条件下的驼蹄瓣逐步顺应生长环境,形成了与气候环境因素相适应的分布格局。随着海拔的升高,其光强增加但温度随之降低,上角质层厚度、下角质层厚度、上表皮厚度、下表皮厚度、栅栏组织厚度及海绵组织厚度和海拔成正相关,上述叶片指标与所在地的纬度和经度无明显相关性。研究表明,随干旱程度的加剧,叶片小而厚,这种变化既减少叶片的蒸腾面积,提高了贮水性,但又不影响植物同化作用的正常进行,是植物从形态结构上对干旱少雨环境作出的适应性改变[14]。
叶肉组织中栅栏组织与海绵组织的分化程度反映了生境中的水分状态,研究结果表明叶肉组织在一定环境条件下存在着一个最佳的栅栏组织厚度与海绵组织厚度的比值(P/S,栅海比),即一定程度上表现为P/S值相对越大,则植物净光合速率越大,耐寒耐旱性越强[15]。由实验所测数据分析可知:栅海比值与海拔高度呈正相关。这与沈宁东等研究的不同海拔间藏茴香叶片厚度均达到差异显著水平,并且不同海拔间上表皮的长度和厚度随海拔的升高而逐渐增加[16]。研究发现:驼蹄瓣叶片具有旱生植物的典型性特征,如叶片厚度较大,T-1叶片厚度可达530.986 μm,这与研究区的降水量少和蒸发量强相适应,较厚的叶片可以延长水分从叶脉向表皮扩散的距离,从而防止水分过度蒸腾,因叶肉细胞增大造成的叶片增厚符合盐生植物的特征,这与赵秀明等学者研究提出的植物叶片越厚,储水能力越强相一致[17]。一般认为,小叶型是耐旱的特征,细小的叶型可以减少水分蒸发面积,驼蹄瓣叶表皮外均具有较厚的角质层,较厚的角质层既可以防止高温下叶片内水分的过度散失又可以防止强烈的日照对叶内部细胞的灼伤,是旱生植物结构的特征之一。
杨春雪研究也已经指出随着生长环境的改变,植物会发生一些结构上的适应性变化[19-20]。不同居群驼蹄瓣叶片解剖结构在本质上没有区别,但不同居群大果白刺各测量指标如叶片厚度、上角质层厚度、上表皮厚度、下表皮厚度、栅栏组织厚度和海绵组织厚度之间存在着显著差异,造成差异的原因可能是不同居群的植物的异速生长,是植物对其生存环境做出的生态对策。探讨不同生境对植物叶片的影响,除地形环境和气候外,还应跟土壤含水量有关,有学者研究指出叶片解剖结构指标均与土壤含水量间无显著相关性,说明土壤含水量对叶片解剖结构建成的影响不大[18],在南疆干旱荒漠环境中土壤含水量相差不大且比较低,因此本研究没有考虑土壤含水量对驼蹄瓣叶片解剖结构指数的影响。本研究3个居群之间驼蹄瓣叶片结构指数具有的明显的差异,这种差异性说明了叶片形态和解剖结构对不同生境的生态适应的响应。通过驼蹄瓣叶片的解剖结构观测得出3个不同居群适应环境顺序为T-1>T-2>T-3,这同时体现了驼蹄瓣叶片结构对不同生境表现出较大的变异性以及多样的适应性。
参考文献:
[1] LIU Y X,ZHOU L H.Nitariaceae,Peganaceae,Zygophyllaceae[M]//WU Z Y,RAVEN P H.Flora of China.Beijing: Science Press , 2008,11:41-50.
[2] 李芳兰,包维楷.植物叶片形态解剖结构对环境变化的响应与适应[J].植物学通报,2005,22(增刊):118-127.
[3] 韦存虚,张军,王建军,等.星星草营养器官适应盐胁迫的结构特征[J].植物资源与环境学报,2006,15(1):51-56.
[4] 王勋陵,王静.植物的形态结构与环境[M].兰州:兰州大学出版社,1989:105-138.
[5] 冯育林,吴蓓,李云秋,等.蹄瓣茎的化学成分研究[J].中草药,2009,40(4):536-538.
[6] 宛涛,燕玲,史雪松,等.内蒙古驼蹄瓣属植物与其近缘种霸王遗传多样性的比较研究[J].干旱区资源与环境,2006,20(4):199-203.
[7] 张辰波,史雪松,宛涛,等.内蒙古驼蹄瓣属植物亲缘关系的RAPD分析[J].中国草地学报,2006,28(5):86-90.
[8] 常雪刚,徐柱,易津,等.6种鹅观草属植物叶片形态解剖特征比较[J].草地学报,2011,19(3):443-450.
[9] 寇云玲,罗青红,宋锋惠,等.新疆杂交榛叶片解剖结构研究[J].果树学报,2012,29(3):1063-1068.
[10] 王玉,洪永聪,丁兆堂,等.利用茶树叶片解剖结构指数预测茶树种质材料的抗寒性[J].中国农学通报,2009,25(9):126-130.
[11] 刘世彪,胡正海.遮阴处理对绞股蓝叶形态结构及光合特性的影响[J].武汉植物研究,2004,22(4):339-344.
[12] 张红霞,刘果厚,崔秀萍.干旱对浑善达克沙地榆叶片解剖结构的影响[J].植物研究,2005,25(1):39-44.
[13] 陈庆诚,孙仰文,张国梁. 疏勒河中、下游植物拿落优势种生态一形态、解剖特性的初步研究[J].兰州大学学报,1961(3):61-96.
[14] 马全林,刘世增,严子柱.沙葱的抗旱性特征[J].草业科学,2008,25(6):56-61.
[15] HIGUCHI H,SAKURATANI T,UTSUNOMIYA N.Photosynthesis,leaf morphology,and shoot growth as affected by temperatures in cherimoya(Annona cherimola Mill.)[J]. Trees Sci Hort, 1999,80:91-104.
[16] 沈宁东,韦梅琴,李宗仁,等.不同海拔藏茴香叶片解剖结构比较研究[J].北方园艺,2014(16):31-34.
[17] 赵秀明,王飞,韩明玉,等.新引进苹果矮化砧木的叶片解剖结构及抗旱性[J].西北农林科技大学学报(自然科学版),2012,40(5):136-142.
[18] 黄文娟,焦培培,黄金花,等.塔里木河流域胡杨叶片解剖结构比较研究[J].植物研究,2016,36(5):669-675.
[19] 杨春雪,卓丽环,柳参奎.植物显微及超微结构变化与其抗逆性关系的研究进展[J].分子植物育种,2008,6(2):341-346.
[20] 李志勇,李鸿雁,师文贵,等.牧草种质资源营养器官解剖结构及抗旱性的研究进展[J].安徽农业科学,2010,38(11):5583-5585.
