ghr-miR160负调控棉花叶片衰老机理研究
2018-04-09段珊窦玲玲程帅帅郭亚宁庞朝友马启峰魏恒玲范术丽王寒涛喻树迅
段珊 ,窦玲玲 ,程帅帅 ,郭亚宁 ,庞朝友 ,马启峰 ,魏恒玲 ,范术丽 ,王寒涛,喻树迅*
(1.西北农林科技大学农学院,陕西杨陵712100;2.中国农业科学院棉花研究所/棉花生物学国家重点实验室,河南安阳455000)
植物microRNA (miRNA)是1类长度约为21 nt的内源性无编码功能的单链小分子RNA[1],通过降解靶基因或抑制其翻译在转录后的水平上调节基因表达[2],从而广泛参与植物生长发育的一系列重要进程,包括调控根的形成和发育、叶形态发生和极性及植物幼年期到生殖期转变。同时,miRNA在植物对外界环境应答和非生物胁迫方面发挥重要的调控作用[3]。
棉花是我国最重要的经济作物之一,叶片是棉花最重要的源器官,叶片早衰严重影响棉产量和纤维品质,进而造成棉花减产[4]。叶片衰老是叶片发育的最后阶段,多种激素参与了这一过程的调控,乙烯[5]、脱落酸[6]、茉莉酸[7]和水杨酸[8]可加速叶片衰老,细胞分裂素[9]、生长素[10]和赤霉素[11]则延缓叶片衰老。
miR160在植物的发育过程中起到了重要的调控作用。在拟南芥中,miR160通过靶向调控ARF10、ARF16和ARF17,参与调控植物生长发育及生物胁迫等[12]。在大豆中,过表达gm-MIR160A可增加叶绿素含量,同时其靶基因GmARF和衰老标记基因下调,表明gma-miR160通过抑制靶基因表达对叶片的衰老进程进行负调控[13]。将普通野生稻MIR160f前体转入拟南芥,可使莲座叶数目减少,首次抽薹时间提前,表明MIR160f可能参与了植物花期的调控[14]。而在龙眼中,miR160及其靶基因不仅参与激素信号传导,还参与体细胞胚胎发生过程[15]。番茄的miR160a通过抑制其靶基因SLARF10可影响叶片的长出和早期果实的发育[16]。
棉属A组、D组和AD组基因组测序已经完成[17-19],可通过 psRNATarget(http://plantgrn.noble.org/psRNATarget/)预测miR160的靶基因。经分析其为核酸外切体组分RRP41-like(Ribosomal RNA-processing protein 41),它的功能需要进一步验证。
本研究利用拟南芥内源miR319a前体骨架[20],构建了棉花miR160的过表达载体,通过农杆菌介导法转入拟南芥等方法,初步明确了ghr-miR160的作用,以期为棉花叶片衰老相关基因的研究提供新思路。
1 材料与方法
1.1 材料
棉花miR160及其靶基因GhRRP41L在叶片中的相对表达量分析所用材料为早熟早衰品种中棉所10号 (CCRI 10)和早熟不早衰品种辽4086(Liao 4086),种植于中国农业科学院棉花研究所(河南省安阳市)温室。拟南芥为哥伦比亚型,于本实验室温室种植,生长条件为22℃,16 h光照、8 h黑暗。pMD19-T载体购于TAKARA公司,pBI121载体为本实验室保存,大肠杆菌DH5α购于北京全式金有限公司,农杆菌菌株LBA4404A为本实验室保存。
1.2 取样方法
棉花子叶展平后,开始进行第1次取样,每隔1周取1次叶片,共取4次样,3个重复,于液氮中速冻后存于-80℃冰箱备用。
1.3 DNA及RNA提取
采用改良的CTAB法提取拟南芥基因组DNA[21],并将其保存于-20℃冰箱备用。使用Small RNA Purification Kit(Sigma)进行 microRNA的提取,按照TransScript miRNA First-Strand cDNA Synthesis SuperMix(北京全式金公司)说明书进行反转录,microRNA及cDNA存于-20℃冰箱备用。RNA的提取参照多糖多酚类RNAprep pure plant kit(天根生化科技有限公司)说明书,应用微量分光光度计和1%(质量分数)琼脂糖凝胶进行浓度和质量检测,合格的样品使用 PrimeScriptRT reagent kit(TaKaRa)合成 cDNA模板后,于-20℃保存或直接用于反转录实时荧光定量聚合酶链式反应 (Quantitative reverse transcription polymerase chainreaction,qRT-PCR)。
1.4 MicroRNA160过表达载体的构建
从 miRBase (http://www.mirbase.org/,release 21)获取拟南芥pre-miR319a序列,将ath-miR319a和ath-miR319a*替换为ghr-miR160和ghr-miR 160*,再从拟南芥pre-miR319a上下游各取80 bp碱基,新的序列送于苏州金唯智生物科技有限公司合成。以合成序列为模板,用AtmiR319a引物及 Mighty Amp○RDNA Polymerase Ver.2(TaKaRa)酶获得所需序列后,回收目的片段并连接pMD19-T载体,转入大肠杆菌感受态细胞DH5α,用菌液PCR方法鉴定获得阳性克隆,送至金维智公司进行测序。
对序列正确的菌液提取质粒检测后与经过双酶切 (酶切位点分别是BamHⅠ和SacⅠ)的pBI121载体进行重组,转化大肠杆菌DH5α,用测序验证正确的大肠杆菌重组质粒转化农杆菌。
1.5 MicroRNA160过表达载体转化拟南芥
利用Bechtold等建立的蘸花法转化拟南芥[22]。待种子成熟后收获,为T0种子,消毒灭菌后种于具卡那霉素抗性的1/2 MS培养基,于4℃冰箱春化2 d后,移至16 h光照、8 h黑暗,21℃的培养箱中,生长14 d后,将其移栽至营养土中。待拟南芥生长约40 d,取莲座叶进行PCR检测,分单株收获阳性株,为T1种子,以此类推,直至T3纯合株系。选取阳性纯合株系,进行ghr-miR160的转录水平分析及表型观察。

表1 引物序列Table 1 The primer sequences in this study
1.6 qRT-PCR分析
利用软件Oligo7设计荧光定量引物(表1),以棉花叶片和拟南芥叶片的cDNA为模板,使用康为世纪的试剂盒UltraSYBR mixture with ROX在ABI7500 Real-time PCR system荧光定量仪上进行qRT-PCR分析,反应条件为3步法,扩增程序为:预变性 95℃ 10 min;95℃ 10 s;60℃ 30 s;72℃32 s;40个循环。每个样品技术重复3次,采用 2-△△Ct法,内参基因为GhU6 和GhActin。
2 结果与分析
2.1 MicroRNA160家族分析
在miRBase中共获得pre-miR160数据141条,miR160数据150条,分布在19个物种共41种植物中。利用DNAMAN对150条miR160数据进行多重序列比对 (图1-A),以棉花miR160为例,第11位、14位和16位的C、G和A碱基在所有miR160中高度保守。利用DNAMAN和i-TOL(http://itol.embl.de/index.shtml) 对 141 条pre-miR160数据进行系统进化分析 (图1-B),结果表明,棉花 miR160 和甜橙(Citrus sinensis)、木薯(Manihot esculenta)、葡萄(Vitis vinifera)、毛果杨(Populus trichocarpa)及番木瓜(Carica papaya)的miR160具有较高的同源性。
2.2 棉花miR160的表达模式分析
叶片衰老最显著的变化是颜色由绿变黄,且叶片颜色越黄,衰老程度越严重[23]。观察CCRI 10和 Liao 4086 在 4 个时期(7 d、21 d、35 d、49 d)的子叶表型,CCRI 10子叶生长35 d时颜色明显变黄,而同时期Liao 4086子叶并无明显的黄化表型(图2-A)。因此,此时期CCRI 10的衰老程度比Liao 4086更为严重。qRT-PCR测定ghr-miR160(图 2-B)及其靶基因GhRRP41L(图2-C)的相对表达量表明,在35 d的Liao 4086子叶中ghr-miR160表达量上调,GhRRP41L表达量下调,说明ghr-miR160可能通过作用于靶基因延缓子叶的衰老。
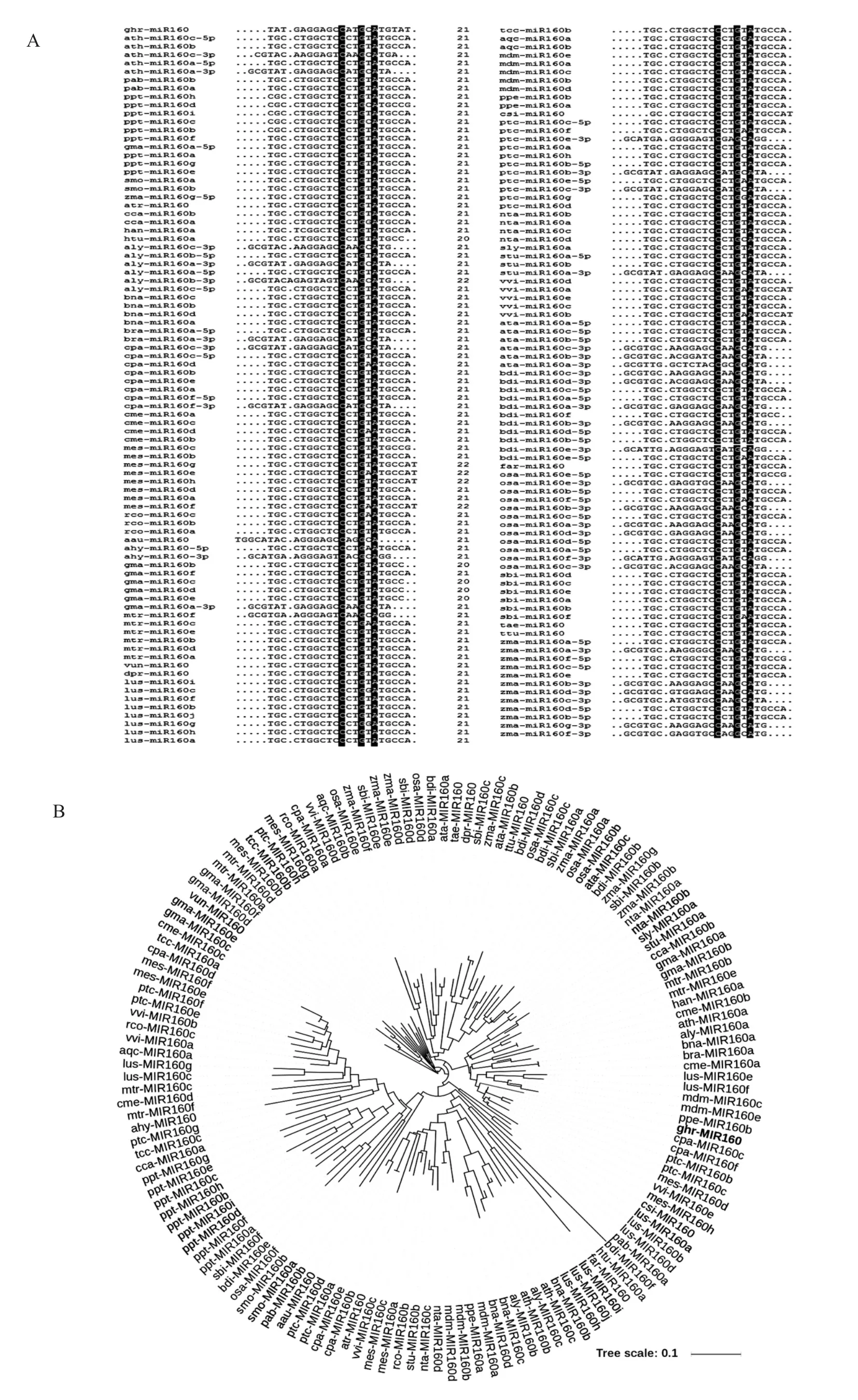
图1 miR160多重序列比对及进化关系分析Fig.1 Multiple sequence alignment and phylogenetic analysis of miR160 family

图2 棉花miR160及其靶基因GhRRP41L在CCRI 10和Liao 4086不同时期子叶中的相对表达量Fig.2 qRT-PCR analysis of ghr-miR160 andGhRRP41Lin CCRI 10 and Liao 4086 at different development stages
2.3 棉花miR160的功能验证分析
提取转ghr-miR160基因拟南芥株系LR-1、LR-2、LR-3和野生型拟南芥叶片的 DNA和RNA,进行PCR验证和表达量分析。在DNA水平,转ghr-miR160基因拟南芥的条带和阳性质粒条带位置相同,而阴性对照和野生型拟南芥无条带(图3-A),说明ghr-miR160基因已经插入拟南芥基因组中;在RNA水平,ghr-miR160在转该基因拟南芥株系中表达量显著上调45~390倍(图3-B),说明ghr-miR160已在拟南芥中转录表达。表型观察结果表明,转ghr-miR160基因拟南芥比野生型拟南芥要晚抽薹,晚开花,叶片衰老延缓(图 4-A)。 分别取生长 20 d、30 d、40 d和 50 d的转ghr-miR160基因和野生型拟南芥莲座叶,测定拟南芥衰老相关基因SAG12的表达量,同时测定叶绿素。结果表明,在拟南芥衰老进程中,SAG12的表达量总体呈逐渐上升趋势,在拟南芥生长的40 d和50 d,转该基因拟南芥SAG12的表达量低于野生型拟南芥(图4-B)。然而,拟南芥叶绿素含量随着叶片衰老总体呈下降趋势,在生长40 d和50 d时,转ghr-miR160基因拟南芥的叶绿素含量高于野生型拟南芥(图4-C)。

图3 转基因拟南芥阳性检测和ghr-miR160表达量变化Fig.3 The checking of transgenicArabidopsisby PCR and qRT-PCR of ghr-miR160
3 讨论
叶片衰老是植物叶片发育的最后阶段,在衰老过程中,叶绿素丧失,大分子物质如蛋白质、脂类、核酸等被降解,导致光合作用减弱,同时衰老组织的营养物质被运输到幼嫩组织和生殖器官中[24]。棉花叶片衰老是1个普遍性的问题,发生于各国的产棉区,导致棉花减产降质[4]。目前对于棉花叶片衰老的研究主要集中在生理生化指标上;因此,研究棉花早衰过程中的关键基因具有重要意义。
根据Zhang等[25]对71个物种37个miRNA家族研究表明,至少存在于10个不同物种的miRNA家族是高度保守的。本研究发现植物miR160基因在进化上高度保守,这与Zhang等[25]的研究一致。保守的miRNA基因往往形成复杂的调控网络,功能具有多样性[26]。在所有miR160分布中,玉米(Zea mays)、亚麻(Linum usitatissimum)、 小立碗藓 (Physcomitrella patens)、 木薯(Manihot esculenta)中分别发现 13 个、10 个、9 个和 8 个;而在菊芋(Helianthus tuberosus)、向日葵(Helianthus annuus)和小麦(Triticum aestivum)中分别发现1个。由此可见,miR160在低等植物和高等植物中都存在,是1个相对古老的家族;但是各个植物之间的存在数量有明显差别,说明miR160基因在进化中,有些旧的基因消失,也有些新的基因出现,导致其在基因组呈分散分布,也进一步说明miR160家族调控的复杂性和研究价值。
为了探究棉花miR160的表达模式,本研究利用qRT-PCR分析了CCRI 10和Liao 40864各时期子叶的ghr-miR160及靶基因GhRRP41L的表达量。ghr-miR160在Liao 4086在子叶生长35 d中表达量显著高于CCRI 10。Xu等[27]也发现在水稻灌浆后期,miR160在不早衰品种的叶片中优势表达。这表明ghr-miR160可能调控棉花叶片的衰老。
目前,对于miR160的研究主要集中于其在种子萌发、根冠发育及响应逆境等方面的作用[12]。为了进一步验证miR160在叶片衰老中的功能,用ghr-miR160过表达载体转化拟南芥。在叶片衰老过程中,一些基因被激活,表达增强,此类基因为衰老相关基因[28]。qRT-PCR检测拟南芥衰老相关基因SAG12表达量结果显示,生长40 d的野生型拟南芥的SAG12表达量约为转ghr-miR160基因拟南芥的2倍,而至50 d时高达5倍。并且拟南芥生长40 d、50 d时,转ghr-miR160基因拟南芥叶绿素含量约为野生型拟南芥的18倍。以上结果表明,转ghr-miR160基因拟南芥的衰老速度远远低于野生型拟南芥。张停停[29]发现miR160的靶基因ARF16突变体可使拟南芥提前衰老。以上研究表明,miR160负调控叶片衰老。
4 结论
通过对ghr-miR160进行生物信息学分析、棉花子叶表达模式分析及过表达载体转化拟南芥验证分析,结果表明:其在进化过程中高度保守,且在35 d的早熟不早衰品种辽4086子叶中优势表达;转该基因拟南芥出现晚抽薹、晚开花和叶片衰老延迟现象。测定拟南芥叶片SAG12相对表达量和叶绿素含量结果进一步表明,转该基因拟南芥出现衰老延缓现象。以上结果初步证明棉花miR160基因可能对叶片衰老过程进行负调控,为miRNA在叶片衰老进程中发挥调控作用机制提供了新的思路,也为棉花分子育种提供了潜在的候选基因。
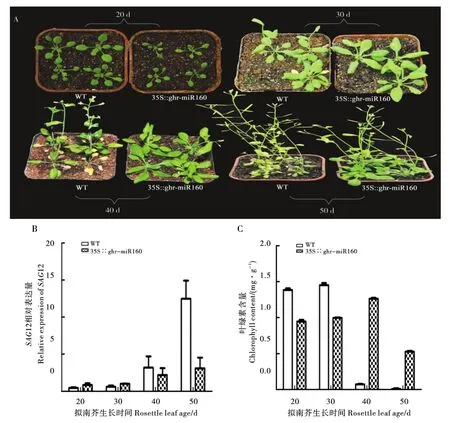
图4 转ghr-miR160基因拟南芥衰老延缓表型及相关指标测定Fig.4 PhenotypefordelayedleafsenescenceandresultsofrelateddetectioninArabidopsisoverexpressedghr-miR160
参考文献:
[1]Bartel D P,Bartel B.MicroRNAs and their regulatory roles in plant[J].Annual Review of Plant Biology,2006,57:19-53.
[2]Bartel D P.MicroRNAs:Genomics,biogenesis,mechanism,and function[J].Cell,2004,116(2):281-297.
[3]Mallory A C,Vaucheret H.Functions of microRNAs and related small RNAs in plants[J].Nature Genetics,2006,38(Suppl):31-36.
[4]王丹.棉花叶片衰老相关基因的数字化表达谱分析[D].杨凌:西北农林科技大学,2011.Wang Dan.Identification and analysis of senescence-associated genes in cotton leaves[D].Yangling:Northwest A&F University,2011.
[5]Abeles F B,Dunn L J,Morgens P,et al.Induction of 33-kD and 60-kD peroxidases during ethylene-induced senescence of cucumber cotyledons[J].Plant Physiology,1988,87(3):609-615.
[6]Zeevaart J A,Creelman R A D.Metabolism and physiology of abscisic acid[J].Annual Review of Plant Biology,1988,39:439-473.
[7]He Y,Fukushige H,Hildebrand D F,et al.Evidence supporting a role of jasmonic acid inArabidopsisleaf senescence[J].Plant Physiology,2002,128(3):876-884.
[8]Van der Graaff E,Schwacke R,Schneider A,et al.Transcription analysis ofArabidopsismembrane transporters and hormone pathways during developmental and induced leaf senescence[J].Plant Physiology,2006,141(2):776-792.
[9]Balibrea L M E,Gonzalez G M C,Fatima T,et al.Extracellular invertaseis an essential component of cytokinin mediated delay of senescence[J].Plant Cell,2004,16(5):1276-1287.
[10]Lim P O,Kim H J,Nam H G.Leaf senescence[J].Annual Review of Plant Biology,2007,58:115-136.
[11]Li J R,Yu K,Wei J R,et al.Gibberellin retards chlorophyll degradation during senescence ofParis polyphylla[J].Biologia Plantarum,2010,54(2):395-399.
[12]Rhoades M W,Reinhart B J,Lim L P,et al.Prediction of plant microRNA targets[J].Cell,2002,110(4):513-520.
[13]李小平,曾庆发,张根生,等.大豆 microRNA基因Gm-MIR160A负调控植物叶片衰老进程[J].广西植物,2015,35(1):84-91.Li Xiaoping,Zeng Qingfa,Zhang Gensheng,et al.Gm-MIR160A,a class of soybean microRNA gene,negetively regulates progress of leaf senescence[J].Guihaia,2015,35(1):84-91.
[14]杨松楠,王姣,陈宗祥,等.普通野生稻miR160f的克隆和功能分析[J].生物技术通报,2014(11):114-118.Yang Songnan,Wang Jiao,Chen Zongxiang,et al.Clone and function analysis miR160f in common wild rice(Oryza rufipogonGriff.)[J].Biotechnology Bulletin,2014(11):114-118.
[15]Lin Yuling,Lai Zhongxiong,Tian Qilin,et al.Endogenous target mimics down-regulate miR160 mediation ofARF10,-16,and-17 cleavage during somatic embryogenesis inDimocarpus longanLour.[J/OL].Frontiers in Plant Science,2015,6:956(2015-11-25)[2016-12-01].https://doi.org/10.3389/fpls.2015.00956.
[16]Hendelmen A,Buxdorf K,Stav R,et al.Inhibition of lamina outgrowth followingSolanum lycopersicumAUXIN RESPONSE FACTOR10(SIARF10)derepression[J].Plant Molecular Biology,2012,78(6):561-576.
[17]Wang Kunbo,Wang Zhiwen,Li Fuguang,et al.The draft genome of a diploid cotton[J].Nature Genetics,2012,44(10):1098-1103.
[18]Li Fuguang,Fan Guangyi,Wang Kunbo,et al.Genome sequence of the cultivated cottonGossypium arboreum[J].Nature Genetics,2014,46(6):567-572.
[19]Li Fuguang,Fan Guangyi,Lu Cairui,et al.Genome sequence of cultivated upland cotton(Gossypium hirsutumTM-1)provides insights into genome evolution[J].Nature Biotechnology,2015,33(5):524-530.
[20]李文超,赵淑清.人工microRNAs对拟南芥At1g13770和At2g23470基因的特异沉默[J].遗传,2012,34(3):348-355.Li Wenchao,ZhaoShuqing.Specific gene silencing ofAt1g13770 andAt2g23470 by artificial microRNAs inArabidopsis[J].Hereditas(Beijing),2012,34(3):348-355.
[21]宋国立,崔荣霞.改良CTAB法快速提取棉花DNA[J].棉花学报,1998,10(5):273-275.Song Guoli,Cui Rongxia.A rapid improved CTAB method for extraction of cotton genomic DNA[J].Cotton Science,1998,10(5):273-275.
[22]Bechtold N.In planta agrobacteriummediated gene transfer by infiltration of adultArabidopsis thalianaplants[J].Comptes Rendus De Lacademie Des Sciences Serie Sciences De La Vie,1993,316(10):1194-1199.
[23]张宝来.水稻叶片衰老的研究进展[J].天津农业科学,2013,19(4):19-24.Zhang Baolai.Advances of research on leaf senescence in rice[J].Tianjin Agricultural Sciences,2013,19(4):19-24.
[24]Lim P O,Kim Y,Breeze E,et al.Overexpression of a chromatin architecture-controlling AT-hook protein extends leaf longevity and increases the post-harvest storage life of plants[J].The Plant Journal,2007,52(6):1140-1153.
[25]Zhang Baohong,Pan Xiaoping,Charles H C,et al.Conservation and divergence of plant microRNA genes[J].The Plant Journal,2006,46(2):243-259.
[26]魏强,梁永宏,李广林.植物miRNA的进化[J].遗传,2013,35(3):315-323.Wei Qiang,Liang Yonghong,Li Guanglin.Evolution of miRNA in plant[J].Hereditas(Beijing),2013,35(3):315-323.
[27]Xu Xiangbin,Bai Haiqi,Liu Chaoping,et al.Genome-wide analysis of microRNAs and their target genes related to leaf senescence of rice[J/OL].PLoS ONE,2014,9(12):e114313(2014-12-05)[2016-12-01].https://doi.org/10.1371/journal.pone.0114313.
[28]Gan S,Amasino R M.Making sense of senescence(molecular genetic regulation and manipulation of leaf senescence)[J].Plant Physiology,1997,113(2):313-319.
[29]张停停.拟南芥生长素响应转录因子ARF16生物学功能的初步探究[D].淮北:淮北师范大学,2013.Zhang Tingting.The function of auxin response transcription factorARF16 inArabidopsis thaliana[D].Huaibei:Huaibei Normal University,2013.
