小家鼠的探究
2018-04-02者金付
者金付

1 分類地位
小家鼠(Mus musculus Linnaeus,1758),别名小老鼠、小耗子、鼷鼠、小鼠等。分类学上属啮齿目(Rodentia),鼠形亚目(Myomorpha),鼠总科(Murioidea),鼠科(Muridae),鼠亚科(Muridae)的小鼠属(Mus)。
早在石器时代,小家鼠就出现在中亚。以后逐渐扩展到欧洲、亚洲和北美洲。现在已遍及全球。在国内,除了西藏少数地区外,各地均可见到。目前一般认为,小家鼠大约在 35~90 万年前起源于印度次大陆,继而向东、西、北三个方向分化出三个亚种,即欧洲亚种(M. m. domesticu)、华南亚种(M. m. castaneus Waterhouse,1843)和 指明亚种(M. m. musculus Linnaeus,1758)。欧洲亚种主要分布于东亚和东南亚,华南亚种主要分布于西欧、中东和北非,指明亚种主要分布于东欧和北亚。
2 分布与生境
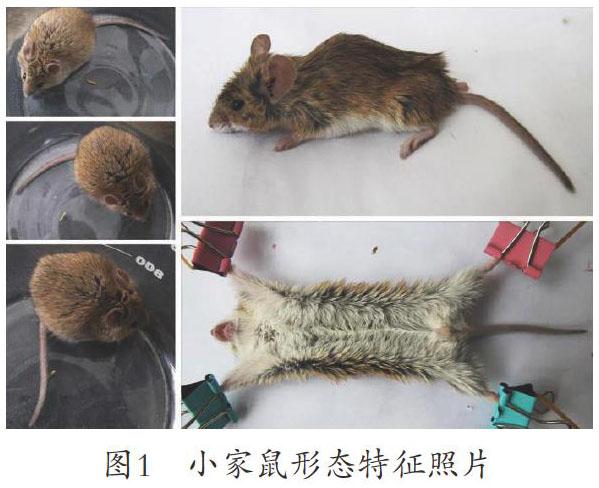
图1 小家鼠形态特征照片
(标本2015年4月19日采自宁夏回族自治区永年县黄河滩地杨树苗圃)
中国小家鼠亚种分化尚无定论。Schwarz and Schwarz(1943)以形态、行为、地理分布951)采用限制性内切酶mtDNA分析技术,发现中国北部分布指明亚种(M. m. musculus Linnaeus,1758)、南部和东南部分布华南亚种、西南部分布有棒杆亚种(M. m. bactrianus)、东南部分地区有指明亚种和华南亚种重复分布、南部部分地区也有华南亚种和棒杆亚种重复分布。Bonhomme(1989)以蛋白质多态型为依据显示我国东北部分布的是指明亚种、南部和东南部分布华南亚种、西南部分布棒杆亚种、南部部分地区为华南亚种和棒杆亚种混合分布区。Tsuehiya等(1994)以外形体尺指标作为分类标准,表明北方地区以甘肃亚种(M. m. gasnuesnis)分布为主,东北地区为甘肃亚种和指明亚种混合分布,中部地区主要分布喜马拉雅亚种(M. m. homourus Hodgson,1845),南部和东南部分布华南亚种。鲍世民等(1999)采用同工酶分析技术和免疫学分析技术测定,综合前人研究结果分析认为,中国小家鼠沿黄河—秦岭—长江为界分类为南、北两大亚种群,即,南方为华南亚种群,北方为指明亚种群。其中,华北亚种(M. m. wagneri Eversmenn,1848)在我国分布于内蒙古、山东、河北 河南、山西、陕西、宁夏。华南亚种(M. m. castaneus Waterhouse,1843)国内分布于江苏、浙江、安徽、江西、台湾、广东、广西。北疆亚种(M. m. decolor Argyropulo,1932)分布于新疆天山山地以北。喜马拉雅亚种(M. m. homourus Hodgson,1845)国内分布在西藏南部。四川亚种(M. m. tantillus G. Allen,1927)分布于四川万县。东北亚种(M. m. manchu Thomas,1909)分布于黑龙江、吉林、辽宁。西南亚种(M. m. urbanus Hodgson,1845)国内分布在贵州、四川、云南。宁夏全境均有小家鼠分布,居民区栖息在住宅、厨房、仓库、养殖场等,野外多在农田、菜园、林地、草甸、草原等生境栖息(图35,照片69)。
3 危害特征
小家鼠对农业的危害很严重,在大发生年代,常给农业造成很大损失。小家鼠危害所有农作物和水果,盗食粮食和各类干果,对贮存苹果啃食很大;初春也啃食麦苗、树皮、蔬菜等。作物收获季危害时一般不咬断植株,只盗食谷穗,受害株很少倒伏。在城市,最大的损失可能不是它吃掉的东西,而是它污染食物和咬坏珍藏的书画、公文、衣物等。虽然小家鼠造成的经济损失难以估测;但几乎所有的人都能意识到小家鼠的存在而造成损失的严重性。此外能传播鼠疫、土拉伦斯病、蜱性斑疹伤寒、丹毒、脉络丛脑膜炎、传染性肾炎、副伤寒、恙虫病、钩端螺旋体病、旋毛虫病、狂犬病、李氏杆菌病等自然疫源性疾病。
9.2 鉴别特征 小家鼠也就是人们常见的小老鼠。体长不到褐家鼠的一半。吻部尖而长,耳朵较大,被毛柔软,无刺毛,体背呈灰褐色或黄褐色。尾细长,鳞环明显(图1)。
9.2.1 形态鉴别
l 测量指标/mm 体重7~20 g。体长50~100,尾长36~87,足长14~16,耳长10~15.5。
l 形态特征 小家鼠为鼠科中的小型鼠,尾与体长相当或略短于体长。头较小,吻短,耳圆形,明显地出毛被外。毛色随季节与栖息环境而异。体背呈现棕灰色、灰褐色或暗褐色,毛基部黑色。体腹面灰黄色到白色,体侧面毛色有时界线分明。足暗褐色或污白色,有的个体白色。尾1色,有些个体尾上面黑褐色,下面为沙黄色(图1)。乳头胸部3对,鼠鼷部2对。
1—上颌骨侧面;2—上颌骨腹面;3—上颌骨背面;4—上门齿和臼齿
9.2.2 头骨鉴别
l 测量指标/mm 颅长19~23,颧宽9.5~11.6,乳突宽8.5~10,眶间宽3~3.65,鼻骨长6.5~7.7,上颊齿列长3~3.7。
l 头骨特征 颅小,呈长椭圆形;吻短;眶上嵴低,鼻骨前端超出上门齿前缘,后端略被前颌骨后端所超出。顶间骨宽大。门齿孔甚长,其后端可达M1中部水平。腭后孔位于M2中部,下颌骨冠状突较发达,略为弯曲,明显指向后方。上门齿斜向后方,其后缘有1缺刻;M1长超过M2和M3之和。M1和M2齿突与鼠属(Rattus)相似。M3很小,内外侧各具有1齿突(图2)。
9.3 发生规律 小家鼠是一种家栖兼野栖的鼠类,其栖息范围甚为广泛,凡人所至之处均能见到它的活动。住家、场院、仓库、农田以及戈壁荒漠等地方都是它的栖居地,尤以住家、场院以及收割后的麦草垛、稻草堆和玉米秸秆堆下更是小家鼠最适宜的栖居场所,这些地方既是它的繁殖地带,又是它越冬的良好场所。在北方生活于农村房屋内的小家鼠到了夏天往往迁至邻近田野、山地或果园中。在谷物或禾草堆下也常发现这种家鼠。此外,小家鼠还常利用轮船、火车为栖居场所。
9.3.1 洞穴 小家鼠的洞穴比较简单,洞穴的结构往往与栖息环境有关(图25)。室内小家鼠通常在地板下面和墙壁空洞中做窝,有时也会在衣箱、抽屉、粮柜、杂物橱中营巢;窝巢常以破布、乱棉絮等物铺垫制成,呈半圆型;在柴草垛或粮食垛下,则在地上或地表浅层建造一个明洞,且有跑道通向各方。在草原或空旷地带,特别是入冬后,小家鼠便集群栖居在较深的洞穴中。野外小家鼠营穴居生活,其洞穴有短浅无巢的临时洞和较长而有巢的居住洞,通常有2~3个洞口,洞道长10~100 cm或更长。雌鼠产仔后,大部分洞口堵塞,只留1个洞口。秋收季节,在田中挖临时洞。
夏季洞穴比较简单,巢室小而浅,呈圆型或卵圆型。巢的成分不定,以掘居洞周围环境及植物生长而异。在草原上,则以杂草筑成,在农田中,以谷物叶茎筑成,在草堆下,以草叶筑成,草堆下多在地表挖掘洞道,筑巢栖居,形成半明半暗的洞穴。在地下筑洞也非常之浅,有1~2个洞口,洞口直徑为2.5~3.0 cm,洞口前多有小土堆。一般洞道长60~80 cm,但在草垛下的明洞,则长短不定,有的可长达200 cm以上。冬季洞穴,巢室较深,洞道略复杂。
小家鼠在居室栖居时,多数只建造巢室,很少挖洞,只在泥土结构房内可见到一浅洞。
在草地中,小家鼠的巢呈球、碗两种形状,筑巢构料较广泛。野栖鼠巢多为球形,家栖鼠巢多为碗状。雄鼠巢小而结构疏松,巢材粗糙,常用谷子、黍子、芦苇、豆叶等组成;雌鼠巢大,且结构紧密,材料也柔软,多为马唐(Digitaria sanguinalis)、莎草(Cyperus microiria)、白草(Pennisetum flaccidum)等制成。球形巢在靠下方留有直径为2.5 cm小孔作为出入通道。巢体积为130 cm3;碗状巢高6~8 cm,深3~5 cm,内径7~9 cm。巢重85~175 g(郭全宝等,1984)。
小家鼠一般在夏季多数分散居住,一洞一鼠。冬季则多集群栖居,10~20只在一个洞系内栖居。如有人在一洞系内曾挖出小家鼠17只,其中成体7只,雄性成体2只,雌性成体5只,亚成体10只,其中雄鼠4只,雌鼠6只。
9.3.2 食性 小家鼠为杂食性,以盗食粮食为主,如玉米、稻子、小麦、高粱以及胡麻、花生等。尤为喜食面粉或面制食品。一只体重14~15 g的小家鼠一昼夜能吃4~5 g面粉或3g 多碎米。初春食源贫乏时,也咬毁青苗;夏季在野外也食草籽和昆虫,数量多时啃食树皮、棉桃和瓜果蔬菜等。其食性与季节、栖居环境食源有关。在高数量时,能取食各种可食之物。室内饲养表明,小家鼠对食物的嗜食程度,直接受它已采食过的食物的影响。小家鼠日食量为3.30±0.25 g。在有饮水的情况下,平均饥饿3.5 d后死去,雄鼠耐饥能力为雌鼠的4倍。小家鼠习惯小量多餐,平均每天取食193次之多,每次仅吃食10~20 mg。其取食场所常不固定,往往在一天之内遍及可能取食的所有地点。
9.3.3 活动 小家鼠活动性强,能主动趋利避害。当适宜空间增加时就扩散,栖息地生态条件恶化就迁出,优化则迁入。这种极强的机动灵活性,不仅使该鼠具有明显的季节迁移特征,而且使之得以随时占据最有利的生活地段,成为富于暴发性的优势种。评价小家鼠栖息地优劣的生态条件可归结为食物和隐蔽条件及其稳定性,以及空间大小、土壤的紧实度等。在天山北麓的老农业区,4月份小家鼠密集地是稻茬地和田间荒地,6月份是小麦地,8月份是水稻田和胡麻地,10月和11月份是水稻田和玉米地。稻茬地、小麦地及苜蓿地,水稻田和玉米地分别是各阶段的最适生境,至于房舍,则是冬季迁入,夏季迁出。
在一般情况下,小家鼠昼伏夜出,在20∶00~23∶00和3∶00~4∶00有两个活动高峰,且以前半夜活动更为频繁。但其昼夜节律在不同地区、不同季节和不同生境可能有些差别。冬季,小家鼠多在雪下穿行,形成四通八达的雪道,并有通向雪面的洞口。当新雪再次覆盖后,小家鼠又由旧雪层到新雪层下活动,久而久之,整个雪被中鼠道层层叠叠,纵横交错。但小家鼠作长距离流窜时,并不在雪被下穿行。春季从居民住宅、粮库、场院麦垛、稻草垛等地方迁往野外。入冬前,除部分栖居玉米秸秆堆放地内越冬外,大部分迂回原处。除季节性迁移外,还随作物生长情况做短距离的迁移。开春后,小家鼠从越冬场所迁往小麦、苜蓿等早春作物地内,以后又随着季节和各种不同作物生长郁闭、开花、结果情况,逐步转向胡麻、小麦、玉米、水稻等作物地集中。
小家鼠具有攀爬能力(但不及巢鼠或黄胸鼠,也不如黄胸鼠机警),可沿铁丝迅速爬上滑下,在农田中,可沿作物茎秆攀援而上,并在穗间奔跑,如履平地。小家鼠也能利用粗糙的墙面向上爬,到梁、天棚上活动。在新疆,土坯房多用壁纸糊顶,冬夜小家鼠常在纸顶上奔跑打闹,影响住户休息。小家鼠从2.5 m高处跳下不会受伤,甚至可以从梁上跳下,准确地落在盛装食物的容器上盗食。
9.3.4 雌雄比 小家鼠在正常情况下雌多雄少,随着窝仔数的增高,雄性有增多的倾向。小家鼠的年龄组成在不同密度下有所不同,在高数量年,亚成年组比重高,在低数量年成年组比重偏高。此种现象反映了种群特征在不同密度水平下的变化。前者是前期刺激种群大发生的有利因素和后期高密度抑制效应双重作用的结果;后者则基于前期出生率低而后期生长发育快速的双重作用。
9.3.5 繁殖 小家鼠的繁殖力极强,条件适宜,一年四季均能繁殖,以夏、秋两季繁殖力最高。年产6~8胎,妊娠期约20~26 d,产后又能马上交配受孕,每胎产仔5~8只,最多14只以上。一般体重达7 g时即能性成熟。雌的体重7~11 g,体长66~71 mm者为亚成体;体重在11 g以上,体长7 2mm以上者为成体。雄的体重7~10 g,体长62~7l mm者为亚成体,体重超过10 g,体长超过71 mm以上者即为成体。幼鼠14~15 d睁眼。不到三星期就能独立活动。一般2个月即性成熟。实验小白鼠系由小家鼠白化而来。
在天山北麓,6~10月是田野小家鼠的繁殖盛期,怀孕率在50%以上,雄性睾丸下降率在90%以上(10月份除外)。而11月到3月下旬,怀孕率在30%以下,平均胎仔数和睾丸下降率也较低。小家鼠妊娠期约19d,平均胎仔数为7.86只,幼鼠在2.5月龄时即达性成熟,产仔间隔和年产窝数都随生境不同而发生变异。如在新疆北部,产仔间隔平均38.9(23~80)d,平均年产9.4窝;而在西宁市,产仔间隔平均50.9(25~102)d,平均年产7.1窝。
小家鼠实验种群胎仔成活率(母腹中胎儿的存活率)为94.2%,初生到性成熟的存活率为47.65%(初生至25 d为57.88%,26 d至2.5月为82.76%)。自然种群胎仔存活率为76.9%,初生至性成熟存活率从30%~35%到87%(朱盛侃等,1993)。
小家鼠营家庭式生活,在繁殖季节,由一雌一雄组成家庭,双方共同抚育仔鼠。待仔鼠长成,则家庭解体,有时是双亲先后离去,有时是仔鼠离巢出走。在繁殖盛期,也可发现亲鼠已孕,仔鼠仍在,甚至有几代仔鼠与亲鼠同栖一洞者(最多可超过15只),每一家庭,有不超过数平方米的领域。
9.3.6 种群数量调节 小家鼠有特别强大的生殖潜能,但其潜能的发挥受到其自身种群密度和多种环境因素的制约。种群密度的改变可导致个体极显著的生理变化和行为改变,在高密度的种群中,观察到肾上腺皮质增生,幼体胸腺萎缩和雌雄个体生殖腺的萎缩,表现出繁殖受到强烈的抑制。加上气候、农业收成和疾病的影响,使得小家鼠种群动态十分复杂多变。在个别年份,其数量可猛增千倍左右。如新疆天山北麓于1967年,伊犁谷地于1970年,都曾发生过小家鼠的大暴发,造成极大的危害。
小家鼠繁殖指数与密度呈显著的负相关。因此,小家鼠种群在一年中的生殖动态,在较大程度上受控于其数量水平,并且反应灵敏。种群增长率与其前一时段种群基数关系密切而直接。在北方,季节性抑制发生在8~10月份。
9.3.7 种群动态 小家鼠的数量,在北方属典型的后峰型,每年到一定时期就会迅速增长,数量曲线陡然上升,具有指数式增长的特征,一旦受环境阻力或其他限制,又会立刻停止增长或骤然下降,表现为变幅很大,极不稳定。在天山北麓,其数量低谷在4月份,高峰在10月份,除自身的生殖抑制外,这主要是冬季严寒造成的。在珠江三角洲,农田小家鼠的数量波动曲线仍为单峰型,其最低点出现在6~7月份,峰期在冬季,这是由于6~7月暴雨盛行,寄生虫感染率高,而冬季气候温和,食物充裕所致。可见少家鼠在不同地域季节消长的时序虽有不同,但基本形态是相同的。
9.3.8 种群爆发
l 爆发周期 小家鼠数量的年间变化幅度也很大,并无一定周期,但并非没有规律。如在高数量年后,一般紧接着一个或几个低数量年,而且前一年数量越高,随后的数量越低,影响越久。根据其数量水平和危害特点,可将小家鼠的数量分为大暴发年、小暴发年、中暴发年和低数量年。
l 爆发进程 小家鼠为R-对策者,具有大暴发的固有特征。仅20世纪在北美、澳大利亚、欧洲和前苏联就发生将近20次,在我国新疆,1932年、1937年和l967年(天山北麓农区),以及l955年、1970年(伊犁谷地)共发生5次。1957年后我国学者进行了广泛调查,1970年大暴发时夏武平等专家亲临现场,目睹其惊人数量,进行了深入的调查研究。大暴发进程的特点是初期种群数量增长快,中期密度特高和后期种群数量急剧下降。朱盛侃等(1993)对小家鼠种群大暴发的主要特点作了总结:
(1)数量高 各主要栖息地捕获率均超过50%。由于夹日法的固有缺点,不能反映高密度种群的数量,所以其实际密度更高。
(2)发生早,持续期长,消退急骤 两大暴发年5月份鼠密度很高,6~10月份成群危害,到下第一场雪时则突然消失。
(3)行为改变 集结流窜,白天也活动,无所不食。
(4)危害烈,破坏力特强 可以成片毁灭庄稼,咬毁室内各种物品,酿成地区性特大灾害。
(5)鼠个体趋小,抗逆性变弱 数量中常年份小家鼠平均体重17.2 g,每千克58只,大暴发年平均体重不足14 g,每千克72只。中常年份雪后小家鼠仍很活跃,在野外也能保持相当数量,大暴发年的头场雪后鼠群骤逝,表明其耐寒性极弱。
(6)生理改变 生殖腺萎缩,10月上旬即全部停止繁殖,雌成鼠无一怀孕。
(7)种群崩溃 大暴发后次年种群数量必降至最低点,即种群“爆炸”以后出现“崩溃”现象。
l 数量动态 小家鼠数量变动非常大,在大发生年代,数量猛增,分布区蔓延擴大。数量猛增后,又急剧下降,转入低潮,雄性个体副睾无精子,雌体子宫变为白细,然后再逐步恢复到正常数量。除地域因素外,其数量又与季节有关,在居民住宅、粮库等地,冬春数量高,夏季数量偏低,秋季以后数量又逐渐上升到一定水平,即在作物收割完毕、气温亦逐渐变寒时,小家鼠则由野外田地迁入居民住宅、场院、粮库、稻草垛内栖居越冬;农田和野外则是冬春两季数量低,夏季数量最多,10月份达到最高峰。
9.3.9 共栖关系 在新疆北部农区,小家鼠分别与灰仓鼠、红尾沙鼠、小林姬鼠、根田鼠可并存于同一生境,特别是小家鼠与灰仓鼠,为农村与农田中之恒有种,但其数量变动有各自的规律。在小家鼠大发生时,灰仓鼠、小林姬鼠等的生存条件也受到一定影响。
小家鼠和褐家鼠常常可并存于一个栖息环境中。在谷草垛中,这两种鼠分开栖息,小家鼠主要在下部,而褐家鼠在上部。在同一谷草垛中,两者的数量都高。但两者直接相遇时,褐家鼠能咬死小家鼠。因此,当褐家鼠的密度上升时,小家鼠的密度往往相对下降,但在褐家鼠密度下降后,它的密度又可以上升。在许多地区大量灭鼠之后,常常出现褐家鼠减少,而小家鼠相对增多的情况。
