小麦株高及其构成因子QTL定位研究
2018-01-05左文松侯大斌
余 马,张 洪,左文松,李 明,侯大斌*
(1.西南科技大学生命科学与工程学院,四川绵阳 621010;2.四川农业大学小麦研究所,成都 611130;3.泸州市古蔺县农业局,四川泸州 646500;4.绵阳市三台县立新镇人民政府,四川三台 621101)
小麦株高及其构成因子QTL定位研究
余 马1,2,张 洪1,左文松3,李 明4,侯大斌1*
(1.西南科技大学生命科学与工程学院,四川绵阳 621010;2.四川农业大学小麦研究所,成都 611130;3.泸州市古蔺县农业局,四川泸州 646500;4.绵阳市三台县立新镇人民政府,四川三台 621101)
【目的】系统剖析小麦株高与穗长及各茎节间长的遗传关系。【方法】利用包含有110个株系的ITMI重组自交系群体,和已构建的高密度遗传连锁图谱对小麦株高、穗长及各茎节间长进行了QTL定位。【结果】两个参试环境共检测到株高相关位点10个,穗长相关位点14个,倒一节间长相关位点19个,倒二节间长相关位点15个,倒三节间长相关位点17个,倒四节间长相关位点14个,倒五节间长相关位点22个,茎节数相关位点14个。所有位点中,2个株高相关位点,4个穗长相关位点,4个倒五节间长相关位点,4个倒四节间长相关位点,2个倒三节间长相关位点,1个倒二节间长相关位点,2个倒一节间长相关位点,2个茎节数相关位点在两个环境中都被检测到。【结论】共挖掘到125个与株高及各茎节间长相关的QTL位点。
小麦;株高;穗长;茎节间长;QTL
小麦株高是非常重要的农艺性状,该性状不仅影响植株的形态结构,还关系到小麦的田间产量。该性状等于主穗长与各茎节间长的总和,但小麦株高的遗传调控机制并非主穗长及各茎节间长遗传位点的简单叠加[1]。大量的遗传研究表明,小麦株高是由质量基因及微效数量基因共同控制的复杂性状[1-5]。小麦中大多数矮秆基因已经被发掘并开发出与之紧密连锁的分子标记应用于标记辅助选择育种中[1,5-6],其中 Rht-B1、Rht-D1 及 Rht8 矮秆基因在世界范围内应用最为广泛[1]。分别位于4B和4D染色体上的Rht-B1和Rht-D1基因都属于赤霉素不敏感基因,对小麦茎节伸长有重要影[5];Rht8位于2D染色体上,属于油菜素类固醇不敏感基因[7];携带Rht8矮秆位点的导入系比对照株系株高矮化,穗长变短,小穗数无变化,从而产生密穗型表型[8]。
随着小麦基因组学研究技术及分子标记开发技术的不断提高,全球对小麦株高相关QTL位点的报道也日益增加,这类研究多集中于影响最终株高的遗传位点发掘[9-11]。从生物学角度上分析,株高等于小麦穗长和地上部分所有节间长之和,各节间和穗长相互作用,并最终影响和决定小麦产量。因此株高的遗传机制尚待深入剖析研究。本研究拟通过对小麦重组自交系群体及其亲本为材料,利用已构建的高密度遗传图谱对该群体株高、穗长及各节间长度进行QTL定位及条件QTL定位,以期系统探索调控小麦株高的遗传机制。
1 材料和方法
1.1 材料及实验设计
本研究采用由国际小麦作图组织(ITMI)构建的112份F10重组自交系群体(Opata85×W7984)。ITMI群体母本W7984为人工合成小麦,父本Opata85为推广品种,该群体构建参见V.Deynze等发表论文[12]。
ITMI群体及亲本于2008—2010年间种植于雅安市。实验设计采用单粒播种,种植行长1.5 m,株距0.1 m,行距0.3 m,每株系种植5行,常规肥水管理及病虫害防治。小麦收获时各株系取具代表性的5株测量其株高、穗长和各茎节间长度;株高和穗长测定参照李立会编写标准[13],其中株高为地表到穗顶部的长度(不包括芒长);穗长为取样植株中主穗穗基部到顶部的长度(不包括芒长)。
1.2 统计分析
方差分析和协方差分析参照Zhu J.提出的混合线性模型[14]。株高(1)与其组成性状(2)的遗传相关系数计算公式为:

Covg(1,2)为生长期与组成性状的遗传协方差,σ2g(1)为生长期的遗传协方差,σ2g(2)为组成性状的遗传协方差。
1.3 QTL定位分析
ITMI群体以BARC图谱为定位图谱(http://wheat.pw.usda.gov),该图谱包含1410个位点(SSR和AFLP),图谱全长2.541 cM,标记密度1.72 cM每个位点[15]。
所有的表型鉴定值和条件分析表型值都被用于QTL定位分析。QTL定位分析采用IciMapping 2.0软件[16];分析采用基于逐步回归法的ICIM作图法(inclusive composite interval mapping),分析中所有QTL步移速度为1.0 cM,LOD阈值为2.5。
2 结果与分析
普通小麦“Opata85”在两年参试环境中倒一节间及倒二节间显著高于人工合成小麦“W7984”(见表1)。“W7984”株高、倒四节间长、倒五节间长及茎节数显著高于“Opata85”。“Opata85”倒三节间长在参试环境2009—2010年显著低于“W7984”,而在参试环境2008—2009年中差异不显著。此外两个群体亲本穗长差异不显著。
表型相关分析中,株高与其所有构成因素均显著正相关(见表2)。穗长与倒二节间长、倒二节间长、倒四节间长及株高显著正相关。倒一节间长与株高、倒二节间长及倒三节间长显著正相关,与倒五节间长及茎节数显著负相关。除与茎节数相关性不显著外,倒二节间长和倒三节间长与所有参试目标性状均显著正相关。倒四节间长与倒一节间长相关性不显著,与其他所有参试目标性状均显著正相关。倒五节间长与穗长相关性不显著,与倒一节间长显著负相关,与其他参试性状均显著正相关。茎节数与倒一节间长显著负相关,与株高及倒四节间长显著正相关。遗传相关结果与表型相关结果基本一致。
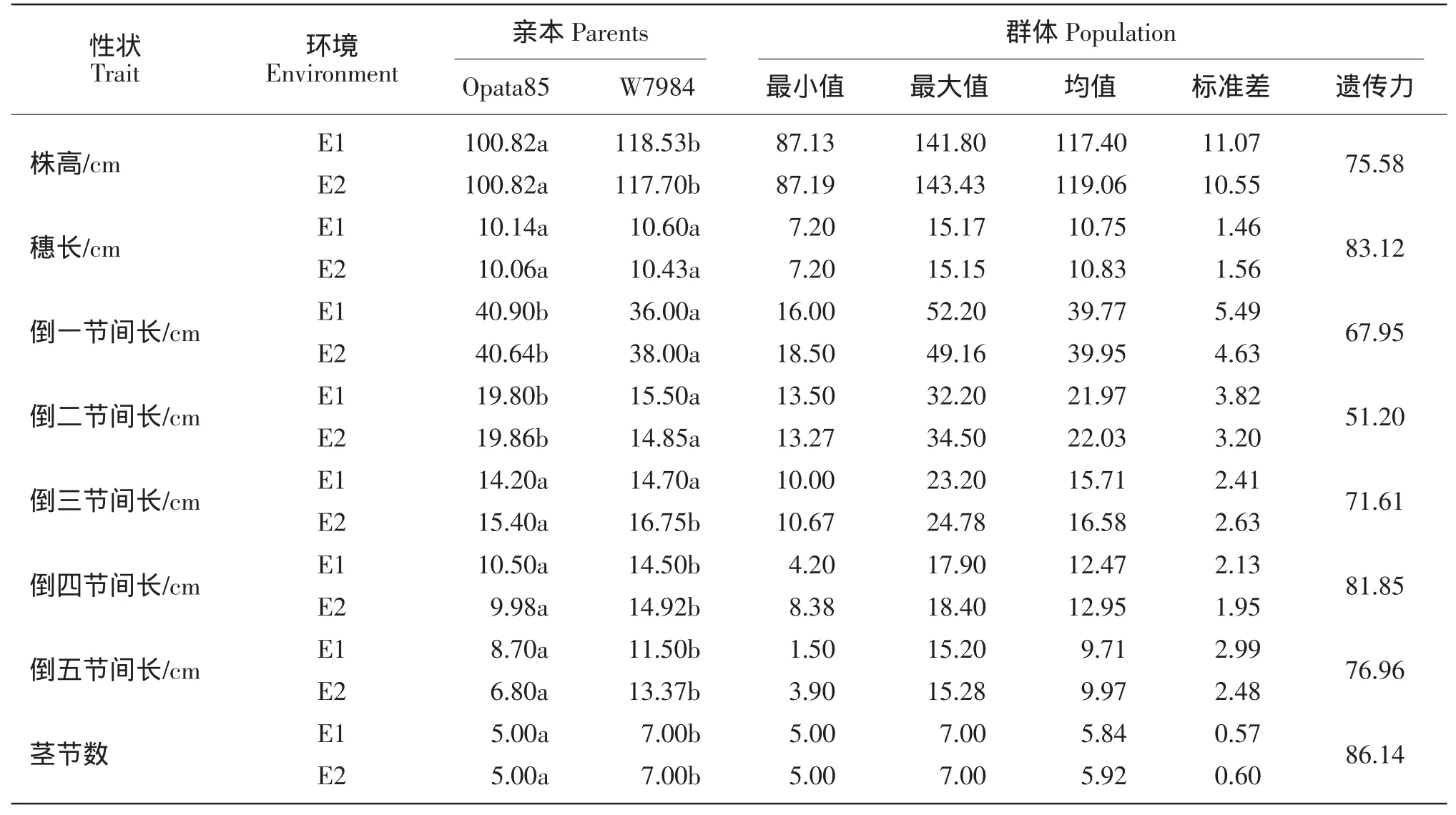
表1 ITMI群体及其亲本目标性状表型测定值Table1 Phenotypic values for objective traits of ITMI population and their parents

表2 株高及其构成因素间的表型相关系数及遗传相关系数Table2 Phenotypic and genetic correlations among plant height and plant height components
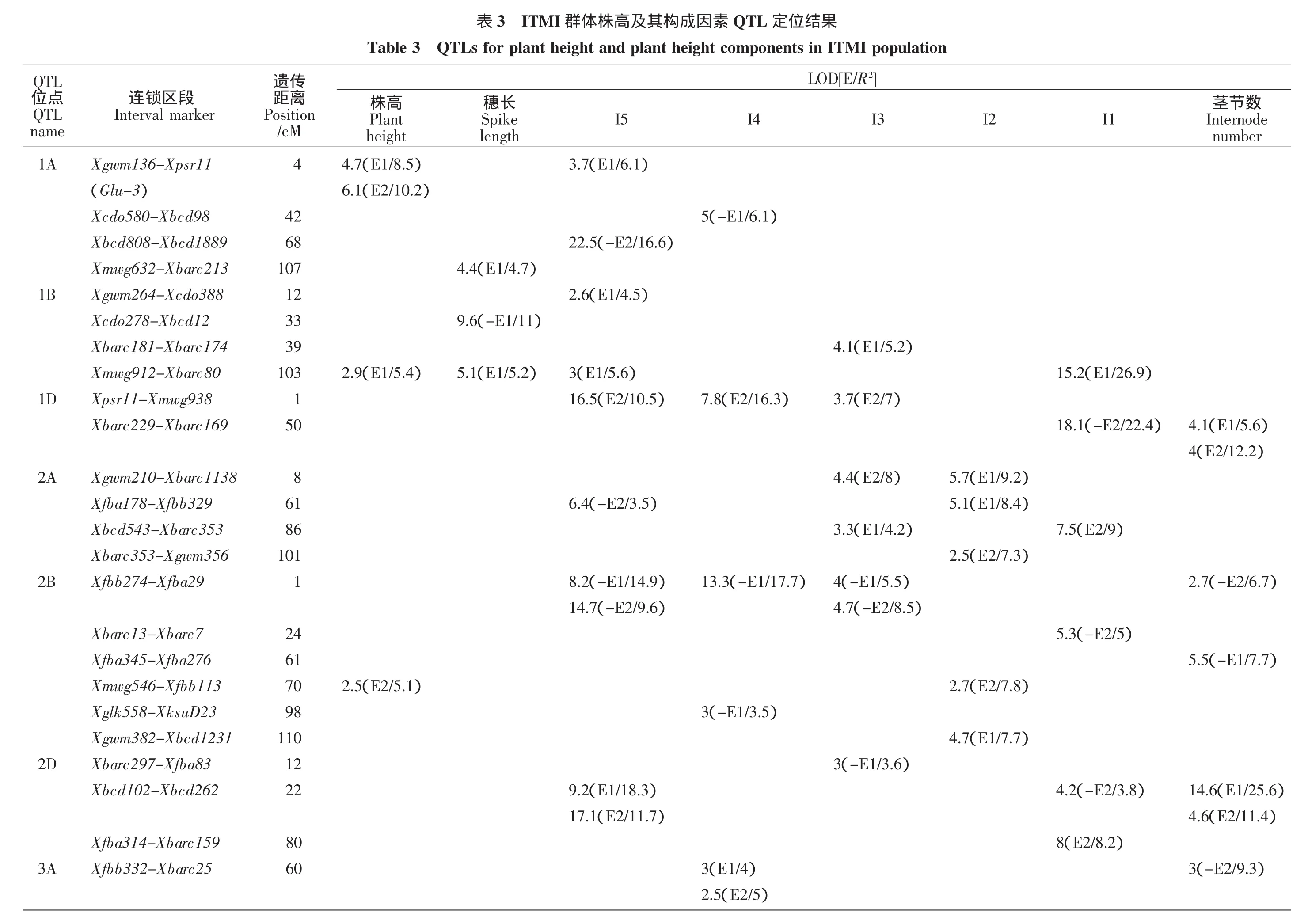




两个参试环境共检测到125个目标性状相关QTL位点,分别分布于除4B以外的所有染色体上(见表3、图1)。其中株高相关遗传位点10个,解释表型变异率5.1%~27.8%,其中1A和5B染色体上两个位点在两个参试环境被检测;穗长相关遗传位点14个,解释表型变异率3.3%~25.0%,其中3D、4A、5A和7A染色体上4个位点在参试环境中都被检测到;倒一节间长相关位点19个,解释表型变异率3.6%~26.9%,其中3D和4A染色体上2个位点在参试环境中都被检测到;倒二节间长相关位点15个,解释表型变异率4.4%~19.7%,其中2A染色体上位点在参试环境中都有检测;倒三节间长相关位点17个,解释表型变异率3.6%~24.3%,其中2B和5B染色体上2个位点在参试环境中都被检测到;倒四节间长相关位点14个,解释表型变异率4.0%~24.7%,其中3A、4D、5A和5B染色体上4个位点在参试环境中都被检测到;倒五节间长相关位点22个,解释表型变异率1.4%~18.3%,其中2B、2D、4A和5A染色体上4个位点在参试环境中都被检测到;茎节数相关位点14个,解释表型变异率4.5%~25.6%,其中1D和2D染色体上2个位点在参试环境中都被检测到。

图1 ITMI群体株高及其构成因素QTL定位结果Figure1 QTLs for plant height plant height components in ITMI population
本研究共检测到33个公共位点区段,分别携带2~5个QTL位点,其中5A上Xgwm293-Xbarc40区段及Xgwm617-Xfba190区段携带了QTL位点数最多。Xgwm293-Xbarc40区段对株高、倒二节间长、倒三节间长、倒四节间长及倒五节间长均影响显著(见表3、图1)。Xgwm617-Xfba190区段对株高、穗长、倒二节间长、倒三节间长及倒四节间长均影响显著。此外,1B上Xmwg912-Xbarc80区段,2B上Xfbb274-Xfba29区段,3A上Xgwm666-Xmwg30区段,5B上Xgwm66-Xfbb121区段,6A上Xgwm494-Xtam36区段对4个性状影响显著。
3 讨论与结论
前人对株高的遗传定位研究报道较多,目前报道的矮秆基因有25个[5,17]。Zhang K.P.等在168份DH群体中(Huapei 3/Yumai 57)发现株高相关位点Qph5A-1与Xgwm186紧密连锁[5]。ITMI图谱中Xgwm186与Xbarc40遗传距离为5 cM,因此Xgwm293-Xbarc40区段内株高相关位点与Qph5A-1具有较高等位性。A.Singh等利用包含261份株系的DH群体将株高相关QTL位点定位于4B和6D染色体上[18]。S.Griffiths等利用4个DH群体进行QTL元分析共识别到16个株高相关元QTL[19]。T.Würschum等利用410份冬小麦群体进行全基因组关联作图分析,将7个株高相关 QTL 位点定位于 1D、2A、4D、5A、5B、6A 及 7D染色体上[4]。此外,影响小麦株高的QTL位点在21条染色体上都有报道[1]。本研究在 1A、1B、2B、3B、5A、5B、6A及7D染色体上共检测到10个株高相关QTL位点,这些位点与前人报道位点可能位于同一区间或具有较高等位性。
相对产量性状而言,小麦穗长遗传力较高,环境敏感性更低[20]。在过去的20年中,穗部形态相关QTL位点被定位于21条小麦染色体上[1]。其中1B、2D、4B、5A及7A染色体上的穗长相关QTL位点在多个遗传背景及环境下都有报道[1,21,22]。本研究也发现ITMI群体中染色体 1A、1B、3D、4A、5A、5D、6A 和7A上都有穗长相关QTL位点分布,这些位点对改良小麦穗部性状具有重要意义。
除穗长以外,目前对小麦茎节与株高遗传关系的报道较少,逯腊虎等在2D染色体上Xwmc111-Xcfd36区段内发掘到第一节间长相关位点1个,第五节间长位点 1个,5A染色体上 Xgwm156-Xgwm328区段内发掘到第四节间长相关位点1个[23]。以上位点与本研究中2D染色体上Xbcd102-Xbcd262区段内倒一节间长相关位点及倒五节间长相关位点,5A染色体上Xgwm293-Xbarc40区段内倒四节间长相关位点具有较高等位性。此外Yu M.等利用ITMI群体及SHW-L1×川麦32重组自交系群体进行了排除穗颈节影响过后的株高条件QTL定位[24]。Cui F.等利用Weimai 8×Jimai 20重组自交系群体及Weimai 8×Yannong 19重组自交系群体进行排除各茎节影响后的株高QTL定位[1]。
小麦株高等于主穗长与各茎节间长的总和,但并非所有控制小麦株高的遗传位点也对主穗长及各茎节间长起主导作用。本研究发掘到的10个株高相关QTL位点中,3个位点被检测到穗长相关QTL位点表达,4个位点被检测到倒一节间长相关QTL位点表达,3个位点被检测到倒二节间长和倒三节间长相关QTL位点表达,2个位点被检测到倒四节间长相关QTL位点表达,4个位点被检测到倒五节间长相关QTL位点表达,1个位点被检测到茎节数相关QTL位点表达。遗传相关分析和表型相关分析结果表明,所有参试性状与株高都有显著相关性,且穗长、倒二节间长、倒三节间长、倒四节间长及倒五节间长相关性较强,这与QTL定位结果基本一致。本研究发掘到的株高相关QTL位点及株高构成因素相关QTL位点对改良小麦株高及产量提升具有重要意义。
[1]CUI F,LI J,DING A M,et al.Conditional QTL mapping for plant height with respect to the length of the spike and internode in two mapping populations of wheat[J].Theoretical Applied Genetics,2011,122(8):1517-1536.
[2]LAW C N,SNAPE J W,WORLAND A J.The genetical relationship between height and yield in wheat[J].Heredity,1973,40:133-151
[3]SNAPE J W,LAW C N,WORLAND A J.Whole-chromosome analysis of height in whea[tJ].Heredity,1977(38):25-36.
[4]WURSCHUM T,LANGER S M,LONGIN C F H.Genetic control of plant height in European winter wheat cultivars[J].Theoretical and Applied Genetics,2015,128(5):865-874.
[5]ZHANG K P,TIAN J C,ZHAO L,et al.Mapping QTLs with epistatic effects and QTL×environment interactions for plant height using a doubled haploid population in cultivated wheat[J].Journal of Genetics and Genomics,2008,35(2):119-127.
[6]ELLIS M H,REBETZKE G J,AZANZA F,et al.Molecular mapping of gibberellin-responsive dwarfing genes in bread wheat[J].Theoretical Applied Genetics,2005,111(3):423-430.
[7]GASPERINI D,GREENLAND A,HEDDEN P,et al.Genetic and physiological analysis of Rht8 in bread wheat,an alternative source of semi-dwarfism with a reduced sensitivity to brassinosteroids[J].Journal of Experimental Botany,2012(63):4419-4436
[8]KOWALSKIA M,GOODING M,FERRANTE A,et al.Agronomic assessment of the wheat semi-dwarfing gene Rht8 in contrasting nitrogen treatments and water regimes[J].Field Crops Research,2016(191):150-160.
[9]MAO S L,WEI Y M,CAO W G,et al.Confirmation of the relationship between plant height and Fusarium head blight resistance in wheat(Triticum aestivum L.) by QTL meta-analysis[J].Euphytica,2010,174(3):343-356.
[10]WANG Z,WU X,REN Q,et al.QTL mapping for developmental behavior of plant height in wheat(Triticum aestivum L.)[J].Euphytica,2010,174(3):447-458.
[11]WU X S,WANG Z H,CHANG X P,et al.Genetic dissection of the developmental behaviours of plant height in wheat under diverse water regimes[J].Journal of Experimental Botany,2010,61(11):2923-2937.
[12]DEYNZE V,DUBCOVSKY J,GILL K S,et al.Moleculargenetic maps for group 1 chromosomes of Triticeae species and their relation to chromosomes in rice and oa[tJ].Genome,1995,38(1):45-59.
[13]李立会,杨欣明,李秀全.小麦种质资源描述规范和数据标准[M].北京:中国农业出版社,2006:124-211.
[14]ZHU J.Mixed model-approaches for estimating genetic variances and covariances[J].Journal of Biomathematics,1992,7(1):1-11.
[15]SONG Q J,SHI J R,SINGH S,et al.Development and mapping of microsatellite(SSR)markers in whea[tJ].Theoretical and Applied Genetics,2005(110):550-560.
[16]LI H H,YE G Y,WANG J K.A modified algorithm for the improvement of composite interval mapping[J].Genetics,2007,175(2):361-374.
[17]WORLANDA J,KORZUN V,RODER M S,et al.Genetic analysis of the dwarfing gene Rht8 in wheat.Part II.The distribution and adaptive significance of allelic variants at the Rht8 locus of wheat as revealed by microsatellite screening.Theoretical Applied Genetics,1998,96(8):1110-1120.
[18]SINGH A,KNOX R E,DEPAUW R M,et al.Genetic mapping of common bunt resistance and plant height QTL in wheat[J].Theoretical and Applied Genetics,2016,129(2),243-256.
[19]GRIFFITHS S,SIMMONDS J,LEVERINGTON M,et al.Meta-QTL analysis of the genetic control of crop height in elite European winter wheat germplasm[J].Molecular Breeding,2012,29(1):159-171.
[20]MA Z Q,ZHAO D M,ZHANG C Q,et al.Molecular genetic analysis of five spike-related traits in wheat using RIL and immortalized F2 populations[J].Molecular Gene and Genomics,2007,277(1):31-42.
[21]XU Y,WANG R,TONG Y,et al.Mapping QTLs for yield and nitrogen-related traits in wheat,influence of nitrogen and phosphorus fertilization on QTL expression[J].Theoretical and Applied Genetics,2014,127(1):59-72.
[22]ZHAI H,FENG Z,LI J,et al.QTL analysis of spike morphological traits and plant height in winter wheat(Triticum aestivum L.)using a high-density SNP and SSR-based linkage map[J].Frontiers in Plant Science,2016(7):16-17.
[23]逯腊虎,魏强,王飞,等.普通小麦农大3338×京冬6号DH系群体株高及节间长度的QTL分析[J].中国农业大学学报,2014,19(1):1-8.
[24]YU M,MAO S L,CHEN G Y,et al.QTLs for uppermost internode and spike length in two wheat RIL populations and their affect upon plant height at an individual QTL leve[lJ].Euphytica,2014,200(1):95-108.
QTL Mapping for Plant Height and Its Components in Recombinant Intercross Lines Population of Wheat
YU Ma1,2,ZHANG Hong1,ZUO Wen-song3,LI Ming4,HOU Da-bin1*
(1.College of Life Science and Engineering,Southwest University of Science and Technology,Mianyang 621010,Sichuan,China;2.Triticeae Research Institute,Sichuan Agricultural University,Chengdu 611130,China;3.Agriculture Bureau of Gulin County,Luzhou 646500,Sichuan,China;4.Lixin Town Government of Santai Country in Sichuan,Santai 621101,Sichuan,China)
【Objective】To system dissect genetic basic of plant height,spike length,and internode length.【Method】An ITMI recombinant intercross lines population contain 110 lines were used to perform QTL analysis based on a high density genetic map.【Results】A total of 125 QTLs for objective traits were identified.For these loci,10 of them were identified for plant height,14 were identified for spike length,19 were identified for the first internode length from the top (IL1),15 were identified for the second internode length from the top(IL2),17 were identified for the third internode length from the top(IL3),14 were identified for the fourth internode length from the top(IL4),22 were identified for the fifth internode length from the top (IL5),14 were identified for internode number.In addition,2 plant height related loci,4 spike length related loci,2 IL1 related loci,1 IL2 related locus,2 IL3 related loci,4 IL4 related loci,4 IL5 related loci,and 2 internode number related loci were expressed in both investigated environments. 【Conclusion】Taken together,all of the 125 QTLs can help to breed out commercial variety with suitable plant height.
wheat;plant height;spike length;internode length;QTL
S512.1
A
1000-2650(2017)04-0465-11
10.16036/j.issn.1000-2650.2017.04.002
2017-02-15
西南科技大学博士基金项目(13ZX7155);国家自然科学基金项目(31601582)。
余马,讲师,博士。*责任作者:侯大斌,教授,博士,主要从事药用植物学研究,E-mail:dbhou@126.com。
(本文审稿:武 晶;责任编辑:刘诗航;英文编辑:刘诗航)
