水稻株高、穗长和每穗颖花数的遗传研究
2016-06-21方兆伟刘金波王宝祥宋兆强卢百关周振玲秦德荣徐大勇
方兆伟,刘金波,王宝祥,迟 铭,宋兆强,李 健,杨 波,卢百关,刘 艳,周振玲,秦德荣,徐大勇
(江苏省连云港市农业科学院/江苏省现代作物生产协同创新中心,江苏 连云港 222006)
水稻株高、穗长和每穗颖花数的遗传研究
方兆伟,刘金波*,王宝祥,迟 铭,宋兆强,李 健,杨 波,卢百关,刘 艳,周振玲,秦德荣,徐大勇*
(江苏省连云港市农业科学院/江苏省现代作物生产协同创新中心,江苏 连云港 222006)
摘要:以两系不育系9311S和恢复系WHR2杂交产生的P1、P2、F1、B1、B2、F2六个世代为研究材料,利用主基因+多基因混合遗传模型分析了株高、穗长和每穗颖花数三个性状的遗传效应。结果表明:穗长和每穗颖花数的遗传均符合1对加性主基因+加性-显性多基因模型,株高表现为1对加性主基因+加性-显性多基因模型或1对加性-显性主基因+加性-显性多基因模型,且均以加性效应为主。讨论了9311S的应用前景。
关键词:水稻;株高;穗长;每穗颖花数;遗传
每穗颖花数是构成水稻产量的重要因子。适当提高植株高度,能够增加群体生物产量,从而提高产量[1-3]。水稻穗长是与水稻产量密切联系的重要农艺性状,对水稻产量影响较大[4]。Rao等对20个高产品种的产量及其各构成性状进行相关分析,发现穗长是对每个品种的产量具有最明显且直接影响的4个产量相关性状之一[5]。因此,研究每穗颖花数、穗长和株高等性状的遗传,对于提高作物产量具有非常重要的理论指导意义。应用现代遗传学的理论与方法,从不同角度对水稻产量相关性状进行深入探讨,是现阶段水稻超高产育种研究的重要内容[6]。前人大多采用NCII遗传试验设计来对性状进行配合力分析,探讨亲本在杂种优势利用或杂交育种中的利用价值[7-12]。虽然双列杂交试验实施简便,但只能利用亲代和F1代有限的遗传信息,无法区分或估计加性和显性以外的上位性遗传效应[13]。以前作者曾利用植物数量性状主基因+多基因混合遗传模型软件对P1、P2、F1、F2世代群体的株高、穗长、总粒数、粒长等性状进行了遗传分析[14-18]。这4个群体有限,所表达的信息有所限制。扩大遗传群体世代,能在较大程度上阐明信息量。两系光温敏核不育系9311S是由连云港市农业科学院自主选育的,该不育系植株较矮,穗大粒多,与众多恢复系测配,杂种F1代的株高介于双亲之间,抗倒伏,穗大。为了研究9311S株高的遗传及其高产理论,本研究选用两系不育系9311S和高秆大穗恢复系WHR2的杂交后代P1、P2、F1、B1、B2、F2六个世代为试验材料,利用主基因+多基因混合遗传模型,研究了水稻的株高、穗长和每穗颖花数的遗传及其基因效应,探讨了杂种优势产生的遗传机制,评价了各性状的遗传潜力和育种价值,以期为遗传改良、杂种优势的利用提供理论依据。
1材料与方法
1.1供试材料
2013年春天,在海南三亚,以两系光温敏核不育系9311S(P1)为母本,以恢复系WHR2(P2)为父本,配制杂交组合F1(9311S/WHR2)。2013年正季,在连云港市农业科学院玉带河试验基地,以9311S为母本,以F1为父本,配制B1(9311S//9311S/WHR2);同时,以F1去雄做母本,以P2作父本,配制B2(9311S/WHR2//WHR2)。成熟时,收获F1代植株上的种子,形成F2群体。
1.2试验方法
1.2.1田间种植与性状测定方法 2014年正季,将试验材料P1、P2、F1、F2、B1、B2六个世代种植于连云港市农业科学院玉带河试验基地。5月7日播种,6月13日移栽。亲本和F1代每材料种植5行,每行10株。F2、B1、B2代分别种植600、450和221株。株行距16.7 cm23.0 cm。单本种植,常规栽培管理。成熟时,测量株高。同时每株取主茎穗考察穗长和每穗颖花数。对于亲本和F1,均调查30株;对于其余世代,调查173~499株。
1.2.2数据分析方法 首先对试验数据进行次数分布分析,利用南京农业大学章元明教授提供的植物数量性状主基因+多基因混合遗传模型软件,分析性状的遗传模型,并根据模型估计主基因和多基因效应值及其方差等遗传参数[19]。试验数据处理在计算机软件Excel中进行。
2结果与分析
2.19311S×WHR2组合的P1、P2、F1、B1、B2、F2世代的株高表现
从表1可以看出:9311S的平均株高为(80.92±4.09)cm, WHR2的平均株高为(146.18±4.65)cm,双亲差异显著(P0.01); F1的平均株高为(128.48±5.41)cm,介于双亲之间,偏向于高亲; B1的平均株高为(107.64±11.76)cm,偏向于低亲; B2的平均株高为(130.23±13.42)cm,偏向于高亲,而且出现超高亲个体; F2的平均株高为(121.38±13.38)cm,偏向于高亲,而且出现超高亲个体; B1、B2和 F2群体均呈连续分布,只有1个峰,没有显示出有效应较大的主基因存在。
2.29311S×WHR2组合的P1、P2、F1、F2、B1、B2世代的穗长表现
从表2可以看出:9311S的平均穗长为(19.62±1.71)cm, WHR2的平均穗长为(27.03±1.79)cm,双亲差异显著(P0.01); F1的平均穗长为(25.42±1.41)cm,介于双亲之间,偏向于高亲; B1的平均穗长为(23.38±2.11)cm,接近于中亲值; B2的平均穗长为(25.14±2.42)cm,偏向于高亲; F2的平均穗长为(24.19±2.48)cm,偏向于高亲,而且出现超高亲个体。B1、B2和F2群体均呈连续分布,只有1个峰,没有显示出有效应较大的主基因存在。
2.39311S×WHR2组合的P1、P2、F1、F2、B1、B2世代的每穗颖花数表现
从表3可以看出:9311S的平均每穗颖花数为(233.03±44.30)粒, WHR2的平均每穗颖花数为(282.97±36.44)粒,双亲差异显著(P0.01); F1的平均每穗颖花数为(355.30±57.09)粒,超过高亲; B1的平均每穗颖花数为(301.54±71.88)粒,超过高亲; B2的平均每穗颖花数为(321.14±82.22)粒,超过高亲; F2的平均每穗颖花数为(325.60±111.00)粒,超过高亲。B1、B2群体均呈连续分布,只有1个峰,没有显示出有效应较大的主基因存在。F2群体呈连续分布,有3个峰,显示出有效应较大的主基因存在。
2.4遗传参数的估计
经遗传模型极大似然值和AIC值(赤池信息准则, Akaike’s Information Criterion)比较,似然比检验(LRT)(如表4),9311S×WHR2组合穗长和每穗颖花数的最适遗传模型均为D-2模型,即这2个性状均表现为1对加性主基因+加性-显性多基因混合遗传。株高的最适遗传模型为D-1和D-2模型,即该性状表现为1对加性-显性主基因+加性-显性多基因混合遗传和1对加性主基因+加性-显性多基因混合遗传。
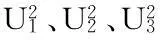
由遗传模型(图1)可以看出:在B1群体中株高存在2个成分分布,分布比例为1∶1,拟合后显示为1个峰;在B2群体中株高也存在2个成分分布,分布比例为1∶1,拟合后显示为1个峰;在F2群体中株高存在3个成分分布,分布比例为1∶2∶1,拟合后显示为2个峰;穗长和每穗颖花数的图形分布和拟合图形分布与株高相似(D-1与D-2一样结果,略)。
根据遗传模型中成分分布及相应的均值,估计出模型中的一阶参数和二阶参数(表5~表6)。
由表5~表6可知,在9311S×WHR2组合中株高适合的遗传模型为D-1,在该遗传模型中,控制株高的1对主基因的基因显性度为0.49,说明控制株高的1对主基因的加性效应和显性效应都同等重要;植株高度高对矮呈部分显性。在B1群体中,控制株高的主基因的遗传率为12.4%,多基因的遗传率为71.84%,说明株高以多基因遗传为主。在B2群体中,控制株高的主基因的遗传率为38.56%,多基因的遗传率为49.33%,说明株高以多基因遗传为主。在F2群体中,控制株高的主基因的遗传率为44.89%,多基因的遗传率为42.93%,说明株高主基因和多基因遗传同等重要。
注: m为中亲值; d为主基因加性效应; [d]为多基因加性效应; h为主基因显性效应; [h]为多基因显性效应。
在D-2模型中,控制株高的1对主基因以加性效应为主。在B1群体中,控制株高的主基因的遗传率为12.4%,多基因的遗传率为71.84%,说明株高以多基因遗传为主。在B2群体中,控制株高的主基因的遗传率为38.56%,多基因的遗传率为49.33%,说明株高以多基因遗传为主。在F2群体中,控制株高的主基因的遗传率为44.89%,多基因的遗传率为42.93%,说明株高主基因和多基因遗传同等重要。
同样,在9311S×WHR2组合中控制穗长的1对主基因以加性效应为主。在B1群体中,控制穗长的主基因的遗传率为39.1%,多基因的遗传率为14.61%,说明穗长以主基因遗传为主。在B2群体中,控制穗长的主基因的遗传率为64.85%,多基因的遗传率为0.0%,说明穗长以主基因遗传为主。在F2群体中,控制穗长的主基因的遗传率为23.58%,多基因的遗传率为42.93%,说明穗长以多基因遗传为主。
在9311S×WHR2组合中控制每穗颖花数的1对主基因以加性效应为主。控制每穗颖花数的多基因的平均显性度为-23.01,超亲偏向于母本。在B1群体中,控制每穗颖花数的主基因的遗传率为66.34%,多基因的遗传率为5.69%,说明每穗颖花数以主基因遗传为主。在B2群体中,控制每穗颖花数的主基因的遗传率为66.39%,多基因的遗传率为0.0%,说明每穗颖花数以主基因遗传为主。在F2群体中,控制每穗颖花数的主基因的遗传率为45.79%,多基因的遗传率为38.95%,说明每穗颖花数以主基因遗传为主。

3讨论
水稻杂种优势主要表现在每穗颖花数的增加上[20]。了解每穗颖花数的遗传,可以有针对性地进行亲本改良和水稻高产育种。笔者、陈献功、江建华等研究认为每穗颖花数性状均受2对主基因+多基因共同控制[17-18,21]。笔者等利用籼粳交材料研究发现粳稻每穗颖花数的遗传表现为1对加性-显性主基因+加性-显性多基因遗传[15]。而本研究发现每穗颖花数性状的遗传符合1对加性主基因+加性-显性多基因模型,以加性效应为主,这可能与本研究的遗传材料差异大有关。而且在B1、B2和F2三个群体中每穗颖花数均表现为主基因遗传,因此在这3个群体中按照穗粒数性状进行选择是有效的,这为育种工作者选育大穗型品种提供了理论依据。
当稻穗较长时,籽粒排列疏松,籽粒灌浆充实,病害尤其是稻曲病发生较轻。朱春杰等利用籼粳群体分析了杂交后代穗部性状的变异,发现穗粒重由3个主成分决定,粒数因子、穗型因子和粒重因子分别贡献了45.49%、17.6%和12.22%,穗较长有使粒数增多、粒重增加的趋势[22]。邹文广等研究发现,穗长受基因的加性效应和显性效应的影响,以加性效应为主,而且遗传率低,适于在晚代选择[23]。本研究发现穗长性状的遗传符合1对加性主基因+加性-显性多基因模型,以加性效应为主。在B1和B2群体中穗长均表现为主基因遗传,在F2群体中表现为多基因遗传,可见回交育种能加速穗长性状的育种进程。
张忠旭等研究认为,在以生物产量的突破为主攻目标,选育超高产中高秆弯曲穗型杂交粳稻时,可以适当放宽对株高的限制,强化缩短第1、2节间长度,增加穗及穗茎节间长度在株高中的比例,以及加强基部茎秆各性状的物理强度,即对抗倒性状进行优化组合,提高水稻本身的耐肥抗倒性[24]。熊振民等认为,在构成株高的诸因素中,与每穗总粒数相关最密切的性状是株高,其次是第II节间长。在矮化育种中并非是植株高度越矮越好,保证有一定的植株高度和适当的第I节间长度是很有必要的[25]。而且,熊振民等研究发现,2个籼稻矮秆品种其矮生性均受与sd1等位的一对隐性矮生基因控制,而l个粳稻矮秆品种受到多基因控制[25]。樊叶杨等研究发现在协青早/密阳46的F2群体中,控制株高性状的基因存在部分显性作用,以加性-显性为主,两个群体中均未检测到显性-显性互作[26]。本研究发现株高性状的遗传符合1对加性主基因+加性-显性多基因模型或1对加性-显性主基因+加性-显性多基因模型,以加性效应为主。而且在B1、B2和F2三个群体中株高均主要表现为多基因遗传,说明控制9311S株高的遗传受多基因控制,这可能与9311S的半矮秆性有关。
从3个性状基因的遗传来看,3个性状均表现为加性效应遗传,说明加性效应为该组合的优势起主导作用,也反映出这3个性状的一般配合力都好。为了充分利用杂种优势,需要在一般配合力高的基础上选择特殊配合力好的组合,才能适应生产的需要。为了发挥9311S的矮秆部分显性作用,可将其与遗传距离较远、丰产性好的恢复系杂交,选育高产、优质、抗倒杂交组合。在半矮秆基因的遗传背景下,通过“矮中求高”,适当提高水稻植株的高度,选育抗倒、高产、优质杂交稻组合[27]。这点在实际育种操作中得到了体现。
参考文献:
[1] 许乃霞,杨益花,苏祖芳.抽穗后水稻株型与高光效群体形成关系的研究[J].耕作与栽培,2009(5):17-18.
[2] 苏祖芳,许乃霞,孙成明,等.水稻抽穗后株型指标与产量形成关系的研究[J].中国农业科学,2003,36(1):115-120.
[3] 袁隆平.选育水稻亚种间杂交组合的策略[J].杂交水稻,1996,16(2):1-3.
[4] 郭小蛟,张涛,蒋开锋,等.水稻籼粳交F1、F2群体穗长QTL比较分析[J].中国农业科学,2013,46(23):4849-4857.
[5] Rao S, Khan M, McNeilly T, et al. Cause and effect relations of yield and yield component in rice (OryzasativaL.)[J]. Journal of Genetics and Breeding, 1997, 51: 1-5.
[6] 林志强,郑燕,蔡英杰,等.水稻长穗大粒RIL群体产量、穗部和谷粒性状的遗传分析[J].福建农林大学学报,2011,40(5):449-454.
[7] 廖伏明,周坤炉,盛孝邦,等.籼型三系杂交水稻主要农艺性状配合力研究[J].作物学报,1999,25(5):622-631.
[8] 朱镇,赵凌,宗寿余,等.籼型两系杂交水稻主要农艺性状的配合力分析[J].江苏农业学报,2004,20(4):207-212.
[9] 周军,邹小云,周瑶敏.籼型杂交水稻产量及其相关性状的配合力及遗传分析[J].江西农业学报,2008,20(12):20-23.
[10] 谭美林,冯明友,张家洪,等.杂交水稻亲本主要农艺性状配合力及遗传力分析[J].贵州农业科学,2009,37(3):6-8.
[11] 刘金波,宋兆强,王宝祥,等.几个新选杂交水稻不育系和恢复系的主要农艺性状的配合力分析[J].西南农业学报,2015.
[12] 洪德林,杨开晴,潘恩飞.粳稻不同生态类型间F1的杂种优势及其亲本的配合力分析[J].中国水稻科学,2002,16(3):216-220.
[13] 顾兴友,郑少玲,严小龙,等.水稻苗期耐盐性遗传的世代平均数分析[J].作物学报,1999,25(6):686-690.
[14] 刘金波,胡文德,王宝祥,等.水稻谷粒外观性状和粒重的遗传研究[J].浙江农业学报,2014,26(3):543-548.
[15] 刘金波,王宝祥,刘晓丽,等.粳稻穗角、着粒密度及每穗颖花数的遗传[J].江苏农业学报,2014,30(5):936-942.
[16] 刘金波,刘晓丽,王宝祥,等.水稻一次枝梗数和二次枝梗数的遗传[J].江苏农业学报,2013,29(6):1187-1192.
[17] 江建华,张启武,洪德林.粳稻穗部性状遗传分析[J].植物学报,2010,45(2):182-188.
[18] 刘金波,洪德林.粳稻穗角和每穗颖花数的遗传分析[J].中国水稻科学,2005,19(3):223-230.
[19] 盖钧镒,章元明,王建康.植物数量性状遗传体系[M].北京:科学出版社,2003:224-260.
[20] Virmani S S, Aquino B C, Khush G S. Heterosis breeding in rice (OryzasativaL.)[J]. Theor Appl Genet, 1982, 63(4): 373-380.
[21] 陈献功,刘金波,洪德林.粳稻直立穗与弯曲穗3个杂交组合6个世代穗角和每穗颖花数的遗传分析[J].作物学报,2006,32(8):1143-1150.
[22] 朱春杰,徐海,郭艳华,等.籼粳稻杂交后代穗部性状变异及其相互关系研究[J].沈阳农业大学学报,2007,38(4):457-461.
[23] 邹文广,许旭明,张受刚,等.水稻长穗大粒种质资源产量性状配合力及遗传力分析[J].福建农业学报,2014,29(4):324-328.
[24] 张忠旭,马兴全,李全英,等.北方超级杂交粳稻育种研究[J].沈阳农业大学学报,2002,43(6):667-672.
[25] 熊振民,闵绍楷,俞桂林,等.云南稻种矮源的进一步筛选及其矮生性的遗传研究[J].江苏农学院学报,1989,10(3):1-5.
[26] 樊叶杨,庄杰云,李强,等.水稻株高QTL分析及其与产量QTL的关系[J].作物学报,2001,27(6):915-922.
[27] 陈俊宇,张振华,庄杰云.水稻株高QTL及其与产量性状和抽穗期关系的研究进展[J].福建稻麦科技,2012,30(1):67-71.
(责任编辑:黄荣华)
Research on Inheritance of Plant Height, Panicle Length and Number of Spikelets per Panicle in Indica Rice (Oryzasativa)
FANG Zhao-wei, LIU Jin-bo*, WANG Bao-xiang, CHI Ming, SONG Zhao-qiang, LI Jian,YANG Bo, LU Bai-guan, LIU Yan, ZHOU Zhen-ling, QIN De-rong, XU Da-yong*
(Lianyungang Academy of Agricultural Science/Collaborative Innovation Center for Modern Crop Production of Jiangsu Province, Lianyungang 222006, China)
Abstract:The genetic effects of plant height, panicle length and number of spikelets per panicle in P1, P2, F1, B1, B2, F2six generations derived from the crossing of the male sterile line 9311S and restorer line WHR2 were studied based on the major gene-polygene mixed inheritance model. The results showed that the inheritances of the panicle length and spikelets per panicle were controlled by one major gene with additive effect plus polygene with additive-dominant effects, and plant height was mainly controlled by one major gene with additive effect plus polygene with additive-dominant effects, or one major gene with additive-dominant effect plus polygene with additive-dominant effects. For three traits, additive effects were principal. The application prospects of 9311S were discussed.
Key words:OryzasativaL.; Plant height; Panicle length; Spikelets per panicle; Inheritance
收稿日期:2015-10-07
基金项目:国家水稻产业体系(CARS-01-01A);江苏省科技支撑项目(BE2014314);江苏省农业三新工程项目[SXGC(2015)138];
作者简介:方兆伟(1963─),男,江苏连云港人,副研究员,从事水稻遗传育种研究工作。*通讯作者:刘金波、徐大勇。
中图分类号:S511.032
文献标志码:A
文章编号:1001-8581(2016)05-0014-06
中央财政农业技术推广资金项目[TG(14)062];连云港科技攻关项目(CN1415)。
