对枝蜈蚣藻的修订研究
——基于形态特征和基因序列分析
2017-11-29田伊林王宏伟
刘 芳 田伊林 王宏伟
(辽宁师范大学生命科学学院, 大连 116081)
对枝蜈蚣藻的修订研究
——基于形态特征和基因序列分析
刘 芳 田伊林 王宏伟
(辽宁师范大学生命科学学院, 大连 116081)
通过形态观察结合rbcL和COⅠ基因序列分析的方法, 对采自广东省汕头市、浙江省温州市和辽宁省大连市的对枝蜈蚣藻Grateloupia didymecladiaLi et Ding的11个标本进行了重新鉴定。结果表明: (1)藻体单生或丛生, 红褐色或深红色, 质地黏滑、肉质、衰老时软骨质, 成熟藻体直立, 高15—50 cm, 主枝明显, 扁平, 宽3—15 mm, 厚约1 mm, 末端渐尖, 1—3回羽状分枝。小枝对生或互生, 生长在主枝上或主枝边缘, 基部缢缩, 长短不一, 最长可达15 cm; 配子体为雌雄异体, 果胞枝生殖枝丛和辅助细胞生殖枝丛均为Grateloupia型, 果胞枝生殖枝丛主枝由5个细胞组成, 辅助细胞生殖枝丛主枝由4个细胞组成(5cpb-4auxb型); 四分孢子囊由四分孢子体的内皮层细胞产生, 呈十字形分裂。以上特征均与亚栉状蜈蚣藻G. subpectinataHolmes一致。(2)基于rbcL基因序列构建的系统树显示, 11个样本之间无碱基差异, 形成独立的进化支, 与产于韩国和日本的亚栉状蜈蚣藻G. subpectinataHolmes碱基差异分别为1 bp (0.08%)和2 bp (0.17%), 属于种内差异; 基于COⅠ基因序列构建的系统树显示, 11个样本之间无碱基差异, 形成独立的进化支, 与韩国产的亚栉状蜈蚣藻碱基差异为1 bp(0.18%), 属于种内差异。根据形态、结构及基因序列分析, 确定对枝蜈蚣藻与亚栉状蜈蚣藻为同一种, 根据优先法则, 将对枝蜈蚣藻G. didymecladiaLi et Ding作为亚栉状蜈蚣藻G. subpectinataHolmes的同物异名。亚栉状蜈蚣藻为中国新记录种。
红藻门; 海膜科; 对枝蜈蚣藻; 亚栉状蜈蚣藻; 形态特征; 基因序列分析
Q949.29
A
1000-3207(2017)06-1273-09
亚栉状蜈蚣藻(新拟)Grateloupia subpectinataHolmes隶属于红藻门Rhodophyta, 杉藻目Gigartinales, 海膜科Halymeniaceae, 蜈蚣藻属GrateloupiaC. Agardh, 主要特征为: 藻体丛生, 红褐色或深红色, 质地黏滑, 衰老后软骨质, 固着器盘状, 直径可达1 cm, 成熟藻体直立, 高15—40 cm, 主枝扁平, 宽4.5—10 mm, 厚1.3 mm, 末端渐尖, 1—3回羽状分枝, 小枝对生或互生, 生长在主枝边缘的小枝间隔为1—7 mm, 达17 cm长, 1—3 mm宽, 基部缢缩; 配子体为雌雄异体, 果胞枝生殖枝丛和辅助细胞生殖枝丛均为Grateloupia(5cpb-4auxb)型; 四分孢子囊由四分孢子体的内皮层细胞产生, 呈十字形分裂。主要分布于中低潮间带[1]。该种于1912年由Holmes[2]根据外部形态和简单的内部结构命名。1914年,Yendo[3]根据外部形态和内部结构的观察将其作为长枝蜈蚣藻G. prolongataJ. Agardh的同物异名。1936年, Okamura[4]将其作为日本产的蜈蚣藻G. filicina(Lamouroux) C. Agardh (现为亚洲蜈蚣藻G.asiaticaKawaguchi et Wang)的同物异名。1998年,Yoshida等[5]指出1914年Yendo的文章中长枝蜈蚣藻G. prolongata的凭证标本包括两种蜈蚣藻, 分别是蜈蚣藻G. filicina和舌状蜈蚣藻G. livida(Harvey)Yamada, 同时, 又将亚栉状蜈蚣藻G. subpectinata作为蜈蚣藻G. filicina的同物异名。2004年, Faye等[1]对分布于日本的该种样本的营养结构和生殖结构进行了详细的观察, 并对其rbcL基因序列进行了分析, 认为该种既不是长枝蜈蚣藻G. prolongata的同物异名, 也不是蜈蚣藻G. filicina的同物异名, 而是蜈蚣藻属一个独立的种, 沿用其最初的种名Grateloupia subpectinataHolmes。2005年, De Clerck等[6]对采自法国、意大利、西班牙、南非、澳大利亚、美国、墨西哥等地的蜈蚣藻G. filicina进行了研究, 发现其中采自法国、西班牙和澳大利亚的蜈蚣藻样本在进行rbcL基因序列分析时, 与日本产的亚栉状蜈蚣藻G. subpectinata形成独立的进化支,表明G. subpectinata也分布于法国、西班牙和澳大利亚。同年, Verlaque等[7]将采自澳大利亚、地中海和日本的G. luxurians(A. Gepp amp; E. S. Gepp) R.J. Wilkes和亚栉状蜈蚣藻G. subpectinata进行了详细的形态比较和rbcL基因序列分析, 将G. luxurians作为亚栉状蜈蚣藻G. subpectinata的异名, 表明G. subpectinata在澳大利亚、地中海和日本均有分布。2013年, Nelson等[8]在新西兰也发现该种。2015年, Yang等[9]对采自韩国的九种蜈蚣藻进行了分类研究, 其中包含亚栉状蜈蚣藻G. subpectinata,表明其在韩国也有分布。到目前为止, 中国没有该种的报道。
对枝蜈蚣藻G. didymecladiaLi et Ding记载于《中国海藻志》第二卷第三册[10], 是由李伟新等基于1963年2月2日采自广东省汕头市潮安海门标本(雌配子体一部分)的形态结构特征建立的新种。最近, 我们进行蜈蚣藻属Grateloupia分类研究时, 发现采自广东省汕头市(模式标本产地)、浙江省温州市和辽宁省大连市的对枝蜈蚣藻G. didymecladia的特征与产自日本的亚栉状蜈蚣藻G. subpectinata的描述几乎一致。为了判断对枝蜈蚣藻G. didymecladia在蜈蚣藻属中分类地位及与亚栉状蜈蚣藻G.subpectinata之间的关系, 对两者的藻体颜色、大小、分枝、质地以及营养结构和辅助细胞生殖枝丛结构、果胞枝生殖枝丛结构、受精后的发育过程、囊果的发育过程以及四分孢子囊的形成过程等生殖特征进行了详细比对。同时, 对两者的rbcL和COⅠ基因序列也进行了比较分析。该研究结果将为《中国海藻志》的修订及再版提供新信息。
1 材料与方法
1.1 标本的采集、处理及形态观察
样本分别于2013年3月至2016年6月间采集于广东省汕头市潮安海门(对枝蜈蚣藻G. didymecladiaLi et Ding模式标本产地)、浙江省温州市南麂岛、辽宁省大连市黑石礁和金石滩等地(表1), 此前均定名为对枝蜈蚣藻G. didymecladiaLi et Ding。在标本采集后, 去除杂质和附生藻类, 制成硅胶干燥标本用于提取DNA, 腊叶标本用于形态观察及10%福尔马林溶液液浸标本制作成冰冻切片, 观察其内部结构。冰冻切片用0.5%(w/v)甲基蓝染液染色, 用Olympus BH2数字显微镜观察, 并用Nikon-HFX-ⅡA照相机拍照记录。标本保存于辽宁师范大学生命科学学院植物标本室(LNU)。
1.2 分子分析
使用植物基因组DNA提取试剂盒(TIAGEN,Valencia, CA, Beijing)提取藻体总DNA, 以提取到的总DNA为模板进行PCR扩增。rbcL和COⅠ基因的PCR反应体系、反应过程和扩增产物的电泳检测方法参照Yang等[9]的设计, 扩增成功后的样品由上海生工生物公司纯化和测序。引物设计参照Wang等[6]的方法, 引物组合为rbcLF1-rbcLR1381(序列分别为: 5′-CAAGGATTAAGAATGAACG CTA-3′; 5′-ATCTTTCCATAAATCTAAAGC-3′)和COⅠF1-COⅠR1(序列分别为: 5′-GGTGGTTGTATGTCTATGC-3′; 5′-CCGGGTCGAAGAAAGAAG-3′),由上海生工生物公司合成。

表1 样本编号、采集信息及基因登录号Tab. 1 Specimen No., collection information and GenBank accession number
从GenBank下载并选取了蜈蚣藻属的17个种[6,8,11—16]的rbcL基因序列和14个种[9,17]的COⅠ基因序列, 与本研究的11个对枝蜈蚣藻G. didymecladia样本的rbcL和COⅠ基因序列分别进行比对分析。
应用软件Clustalx (1.83)[18]进行基因序列比对。使用Maximum likelihood (ML)、Neighborjoining (NJ)、Maximum parsimony (MP)法构建系统树。PAUP4.0[13]和MEGA5.0[19]软件用于碱基差异度分析以及系统树的构建, 计算遗传距离的模型为Number of differences和Kimura 2-Parameter,Bootstrap值为1000。
2 结果
2.1 对枝蜈蚣藻的外部形态
藻体(图1)单生或丛生, 红褐色或深红色, 质地黏滑, 肉质, 衰老时软骨质; 成熟藻体直立, 高15—50 cm, 主枝明显, 扁平, 宽3—15 mm, 其中, 四分孢子体(图1A)主枝宽3—8 mm, 雌配子体(图1B)主枝宽7—15 mm。藻体厚约1 mm, 末端渐尖,1—3回羽状分枝, 近栉状。小枝对生或互生(图2A), 生长在主枝上或主枝边缘, 基部缢缩(图2A、B), 横切面呈圆形或椭圆形。四分孢子体(图1A)的小枝最长可达15 cm, 雌配子体(图1B)的小枝略短。囊果(图2C)近球形(图2C中ch为囊果孔, 直径为10—15 μm), 散布于除固着器外的整个藻体, 稍突出藻体表面。
2.2 对枝蜈蚣藻的内部结构
营养结构: 藻体横切面观是由皮层和髓部构成, 厚度为300—400 μm。皮层由7—12层细胞组成, 背斜排列, 厚度为65—120 μm, 分外皮层和内皮层, 外皮层由3—5层圆形或长椭圆形细胞构成; 内皮层由4—6层不规则的多角形或星状细胞构成。髓部由不规则排列的髓丝构成, 髓丝排列错综交织(图3A、B)。
生殖结构: 藻体为雌雄异体, 果胞枝生殖枝丛与辅助细胞生殖枝丛均由内皮层细胞产生, 果胞枝生殖枝丛主枝包含5个细胞, 生长在一个较大的支持细胞上, 最上面是一个带有一条细长受精丝的较小果胞, 一个3—6个细胞组成的不育侧枝的下位细胞, 一个4—8个细胞组成的不育侧枝的亚下位细胞,与之对向生长的依次是2个分别带有一条不育侧枝的圆形细胞, 果胞枝上的所有细胞包括果胞都带有三级不育侧枝。果胞与直径较大的基细胞相邻, 受精丝有时伸直, 有时弯曲成各种角度指向藻体表面,果胞枝上的所有不育侧枝都弯向藻体表面, 形成瓶状体(图3C、D)。

图1 对枝蜈蚣藻的外部形态Fig. 1 External morphology of G. didymecladia
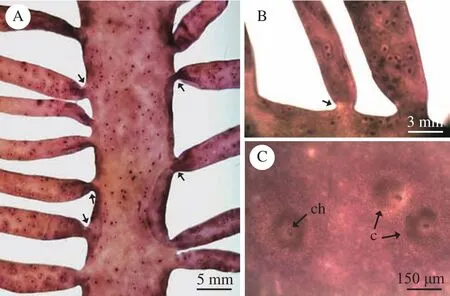
图2 对枝蜈蚣藻(LNU2016060502)的表面观Fig. 2 Surface observation of G. didymecladia(LNU2016060502)
辅助细胞生殖枝丛主枝包含4个细胞, 辅助细胞枝末端细胞为辅助细胞。辅助细胞枝上的4个细胞都带有一个由5—12个细胞组成的侧枝。辅助细胞枝上的所有侧枝都弯向藻体表面形成瓶状体, 与果胞枝生殖枝丛相似。辅助细胞位于瓶状体中心,成熟的辅助细胞形状为圆球形(图3E)。
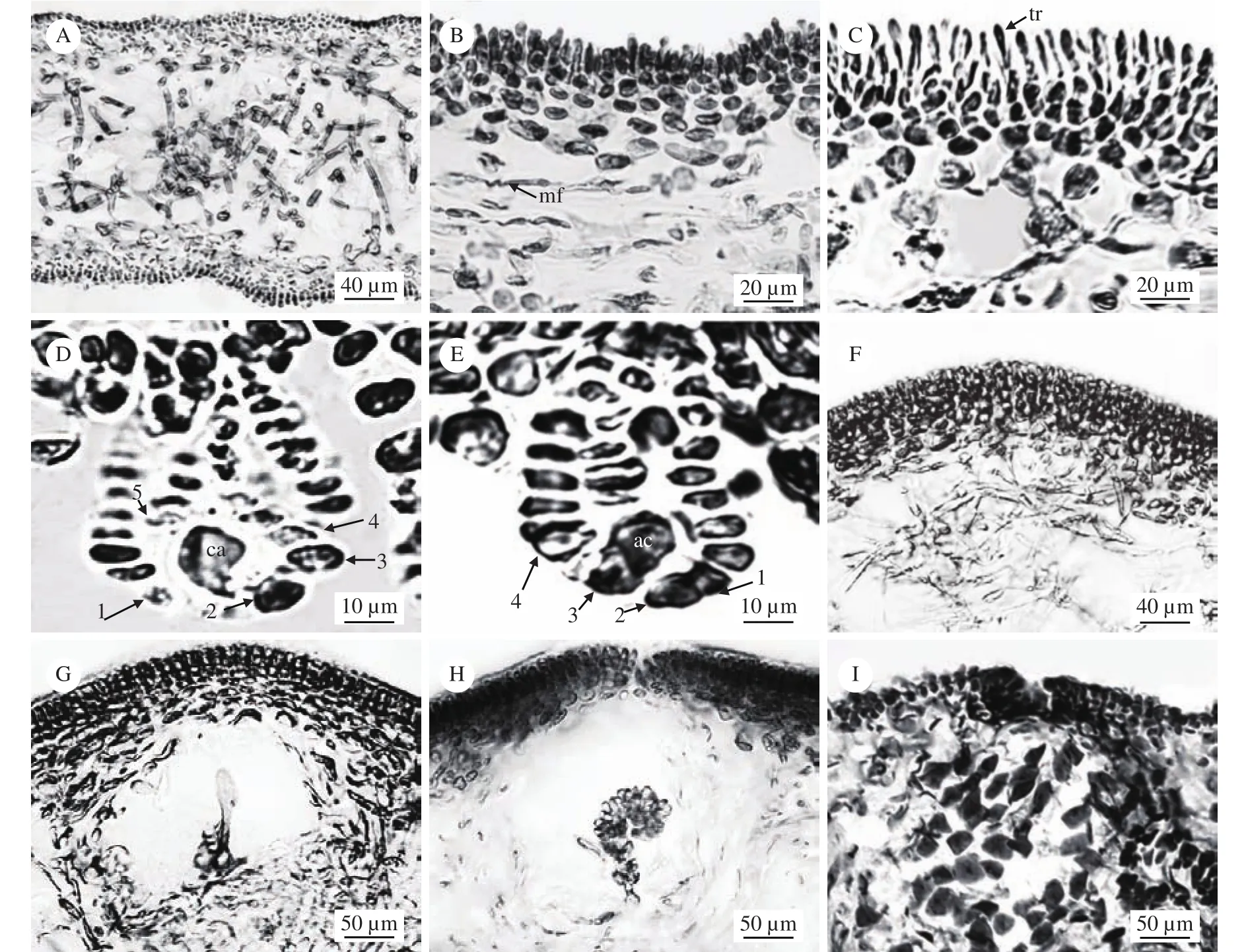
图3 对枝蜈蚣藻雌配子体内部结构Fig. 3 The internal structure of gametophy te of G. didymecladia
精子通过受精丝到达果胞, 受精后的果胞开始增大并与下位细胞融合, 融合细胞产生一条初级联络丝, 延伸并与果胞枝上的基细胞融合, 大融合细胞产生次级联络丝并向辅助细胞移动时, 当联络丝接近辅助细胞时, 它的顶部区域膨大与辅助细胞的近尖端部位立即融合, 融合了一个联络丝之后的辅助细胞扩大, 通过与相邻的瓶状体细胞合并形成一个横向延伸的融合复合体(图3F), 该复合体上产生分枝营养丝, 同时该复合体产生一个简单的初级产孢丝指向藻体表面(图3G), 该产孢丝上的细胞由顶到底长成孢子囊, 形成幼小囊果(图3H)。发育成熟的囊果近球形, 由3—5层髓丝包裹, 孢子囊的顶端有囊孔。果孢子是由产孢丝末端细胞形成的, 待成熟后由囊孔逸出。囊果微突出藻体表面且呈现肉眼可见的斑点状, 散落于除基部外的藻体表面, 囊果直径范围为180—240 μm (图3I)。
四分孢子囊由四分孢子体的内皮层细胞形成,略突出于藻体表面, 散落分布于除基部外的整个藻体 (图4A), 四分孢子囊母细胞(图4B)经减数分裂先形成二分体(图4C), 再形成四分体即为成熟的四分孢子囊, 呈十字形分裂, 成熟的四分孢子囊长为50—60 μm, 宽为15—20 μm (图4D)。
2.3 系统发育分析
rbcL基因序列分析对枝蜈蚣藻G. didymecladia11个样本rbcL基因序列长度均为1301 bp,比对校正后的基因序列长度是1196 bp, 简约信息位点192个。
序列比对结果显示, 11个样本的基因序列间无碱基差异, 与韩国和日本的亚栉状蜈蚣藻G. subpectinata(登录号为GU168549和KF156717)碱基差异分别为1 bp (0.08%)、2 bp (0.17%), 它们共同聚集成一个独立的小分支, 区别于其他种(图5)。对枝蜈蚣藻G. didymecladia与带形蜈蚣藻G. turuturu、稀疏蜈蚣藻G. sparsa和G. phuquocensis的碱基差异分别为27 bp (2.26%)、27 bp (2.26%)、41 bp(3.43%), 与亚洲蜈蚣藻G. asiatica的碱基差异为83 bp (6.94%), 与模式种蜈蚣藻G. filicina的碱基差异为83 bp (6.94%)。
COⅠ基因序列分析对枝蜈蚣藻G. didymecladia的11个样本COⅠ基因序列长度均为610 bp,比对校正后的基因序列长度是554 bp, 简约信息位点137个。
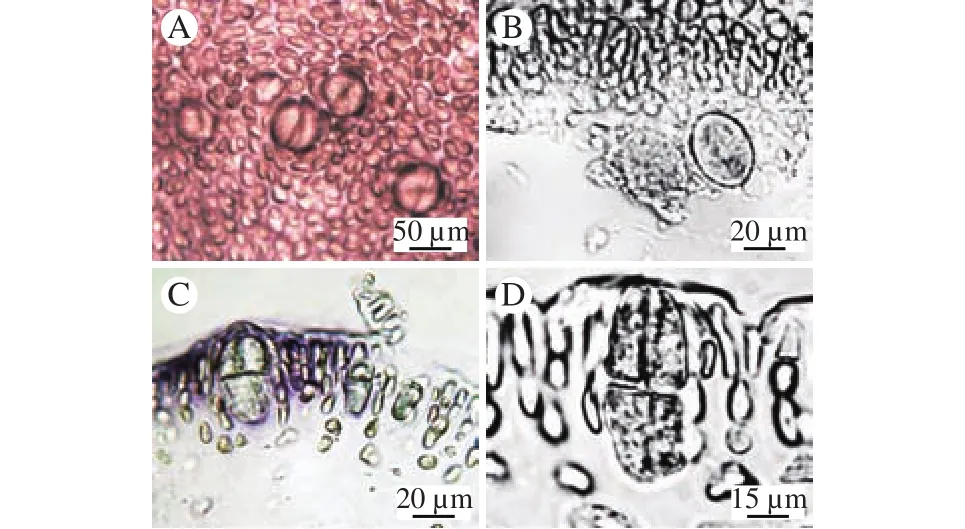
图4 对枝蜈蚣藻四分孢子体内部结构(LNU2014041919)Fig. 4 The internal structure of tetrasporophyte of G. didymecladia (LNU2014041919)
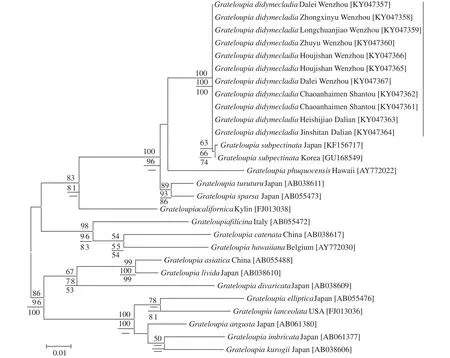
图5 基于rbcL基因序列构建的ML系统树Fig. 5 Maximum likelihood tree was based on partial rbcL sequences data
序列比对结果显示, 11个样本的基因序列间没有碱基差异, 与韩国的亚栉状蜈蚣藻G. subpectinata(登录号为KJ648542)碱基差异为1 bp (0.18%),它们共同聚集成一个独立的小分支, 区别于其他种(图6)。对枝蜈蚣藻G. didymecladia与G. phuquocensis、带形蜈蚣藻G. turuturu和加利福尼亚蜈蚣藻G. californica的碱基差异分别为29 bp (5.24%)、34 bp (6.15%)、63 bp (11.39%), 与亚洲蜈蚣藻G.asiatica的碱基差异为65 bp (11.75%), 与蜈蚣藻G.filicina的碱基差异63 bp (11.39%)。
3 讨论
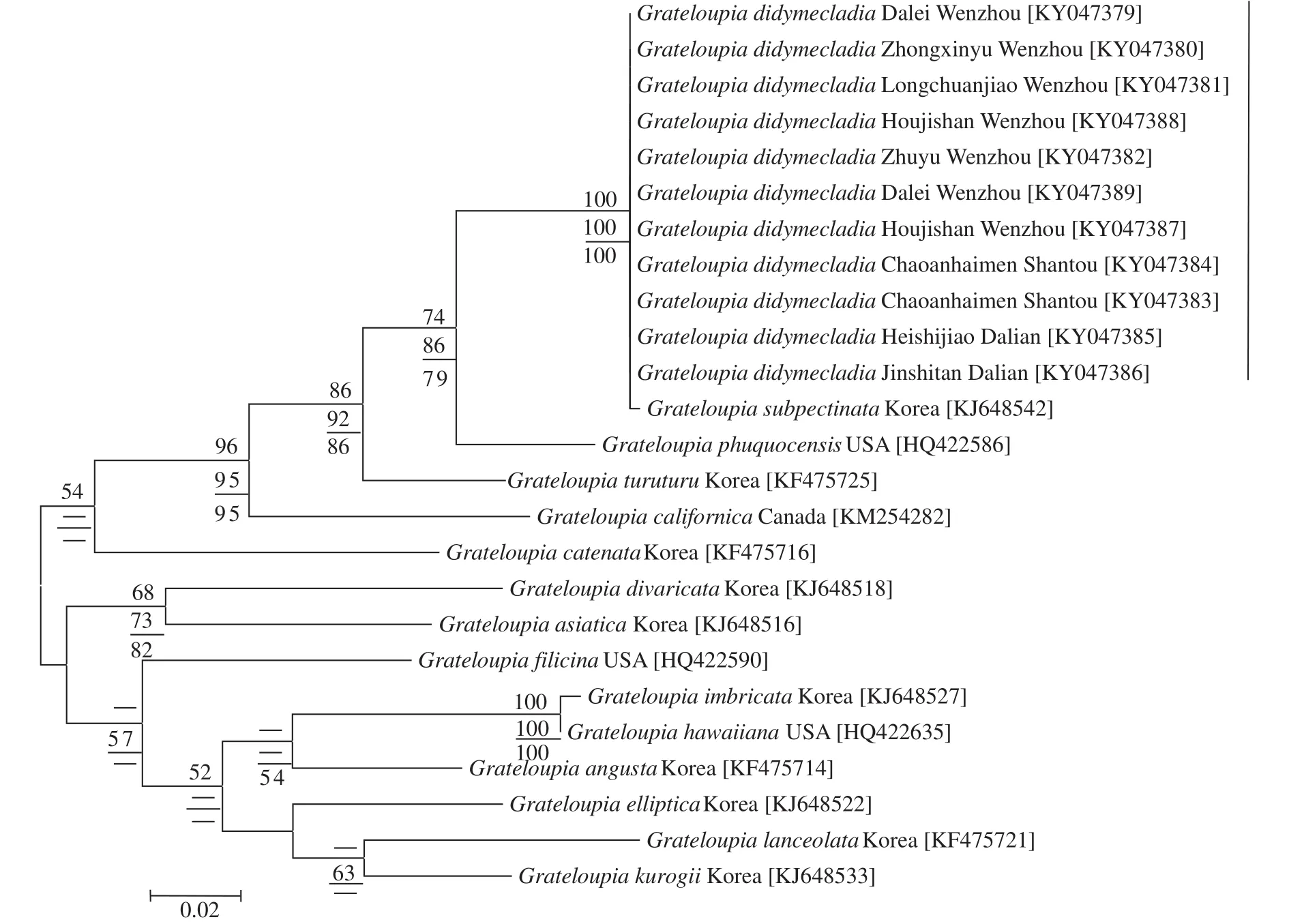
图6 基于COⅠ基因构建的ML系统发育树Fig. 6 Maximum likelihood tree was based on partial COⅠ sequences data
蜈蚣藻属是海膜科中最大的一个属, 运用传统的形态学分类方法对蜈蚣藻属进行准确的分类鉴定较困难。1970年, Chiang[20]根据助细胞的位置及在其周围联络丝所形成的助细胞瓶状体特点, 将Grateloupia型的助细胞生殖枝丛结构作为蜈蚣藻属的重要鉴别特征。1984年, 李伟新等[21]关于蜈蚣藻属生殖结构的报道支持了Chiang的观点。2004年, Kawaguchi等[22]进一步证实了果胞枝生殖枝丛结构与辅助细胞生殖枝丛结构均对蜈蚣藻的分类鉴定具有重要价值。2013年, Gargiulo等[23]在关于蜈蚣藻属的研究中, 根据组成辅助细胞生殖枝丛主枝和果胞枝生殖枝丛主枝细胞数量的不同, 将其分为6cpb-5auxb型、5cpb-4auxb型和4cpb-3auxb型。随着分子生物学技术的飞速发展和不断应用, 越来越多的国内外藻类分类学家采用传统的形态观察与分子分析相结合的方法作为蜈蚣藻分类鉴定的依据。例如, 2000年Wang等[11]对蜈蚣藻中空变型G. filicinavar.lomentariaHowe、蜈蚣藻节荚变型G. filicinavar.porraceaf. lomentaria(Howe) Okamura和管型藻Sinotibimorpha porracea(Martens ex Kützing) Li et Ding进行了形态观察和rbcL基因序列分析, 认为它们是同物异名, 既不是蜈蚣藻G. filicina的变型也不是新种, 而是蜈蚣藻属Grateloupia中的一个独立的种, 即链状蜈蚣藻Grateloupia catenataYendo。2001年, Kawaguchi等[13]对亚洲和意大利产的蜈蚣藻G. filicina进行了形态结构和rbcL基因序列比对研究, 发现二者是完全独立的种,亚洲产的蜈蚣藻应为一新种, 将其命名为亚洲蜈蚣藻G. asiaticaKawaguchi et Wang。上述学者的研究表明, 运用分子分析结合形态学观察, 可以对外部形态相似, 易造成混淆的蜈蚣藻进行更准确的鉴定。作者团队运用分子分析结合形态学观察已经发现并报道了莺歌海蜈蚣藻G. yinggehaiensisWang et Luan[24]、大连蜈蚣藻G. dalianensisWang et Zhao[24]、细弱蜈蚣藻G. tenuisWang et Luan[25]、博鳌蜈蚣藻G. boaoensisWang et Luan[26]、黄海蜈蚣藻G. huanghaiensisWang et Zhao[27]、多枝蜈蚣藻G. ramosaWang et Luan[28]、异枝蜈蚣藻G. variataWang[29]等新种, 并对阳江蜈蚣藻G. yangjiangensisLi et Ding进行了再鉴定, 支持其作为新种的成立[30]。2016年, 李芳等[18]通过形态观察和rbcL基因序列分析, 认为帚状蜈蚣藻G. fastigiataLi et Ding与亚洲蜈蚣藻G. asiatica为同一种, 将帚状蜈蚣藻G. fastigiata作为亚洲蜈蚣藻G. asiatica的异名。本研究通过rbcL和COⅠ基因序列分析, 结果显示对枝蜈蚣藻G. didymecladia与产于韩国和日本的亚栉状蜈蚣藻G. subpectinata碱基差异很小, 分别为0.0—0.17%和0.0—0.18%。2016年, 李芳等[18]提出蜈蚣藻属rbcL基因序列分析种内遗传变异范围为0.0—1.0%。2015年, Yang等[9]提出蜈蚣藻属COⅠ基因序列分析种内遗传变异范围为0.0—1.6%。因此, 对枝蜈蚣藻G. didymecladia与亚栉状蜈蚣藻G. subpectinata之间的碱基差异应在种内遗传变异范围, 属于种内差异。
形态观察结果表明: 本研究的标本与亚栉状蜈蚣藻G. subpectinataHolmes藻体均为直立, 红褐色或深红色, 高度在15—50 cm, 主枝扁平, 1—3回羽状分枝, 小枝对生或互生, 基部缢缩; 质地黏滑, 衰老后软骨质; 皮层数为7—12层; 髓部非中空; 配子体为雌雄异体(表2)。2004年, Faye等[1]认为亚栉状蜈蚣藻G. subpectinata的生殖结构为5cpb-4auxb型,果胞枝主枝由5个细胞组成, 辅助细胞枝主枝由4个细胞组成, 果胞仅与下位细胞融合, 辅助细胞没有融合现象; 四分孢子囊为长椭圆形[长(55—60) μm×宽(18—20) μm]; 囊果为近球形(直径220—250 μm)。本研究的标本生殖结构与其完全一致(表2)。
对枝蜈蚣藻G. didymecladiaLi et Ding记载于2004年出版的《中国海藻志》中(图7)[10], 亚栉状蜈蚣藻G. subpectinata于1912年由Holmes[2]命名。结合形态特征和分子分析研究结果, 我们认为对枝蜈蚣藻G. didymecladia与亚栉状蜈蚣藻G. subpectinata为同一种, 对枝蜈蚣藻G. didymecladia不成立。根据优先法则, 将对枝蜈蚣藻G. didymecladia作为亚栉状蜈蚣藻G. subpectinata的异名, 亚栉状蜈蚣藻G. subpectinata为中国新记录种。
致谢:
感谢中国科学院海洋研究所孙忠民副研究员及辽宁师范大学生命科学学院曹翠翠、姜朋和赵树雨同学帮助采集标本。
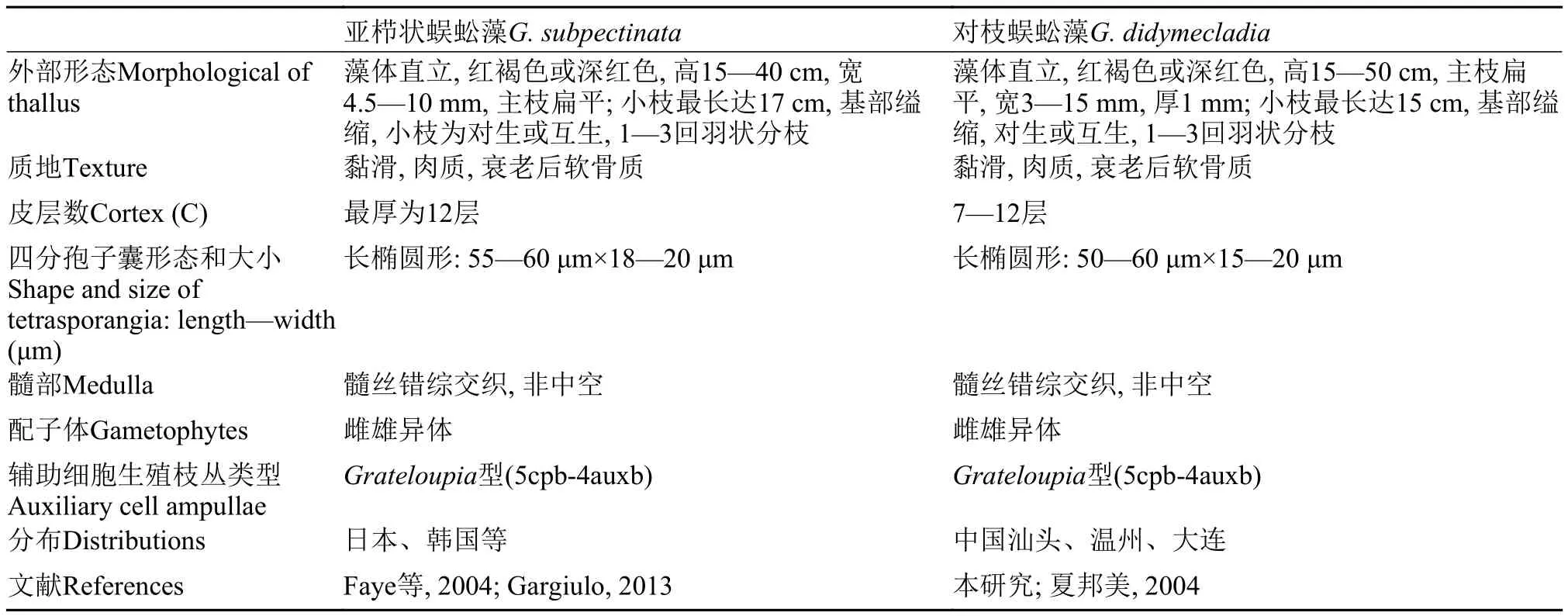
表2 亚栉状蜈蚣藻和对枝蜈蚣藻的形态比较Tab. 2 Comparison of morphological features between G. subpectinata and G. didymecladia

图7 《中国海藻志》中对枝蜈蚣藻的模式标本Fig. 7 Typical specimen of G. didymecladia in Flora Algarum Marinarum Sinicarum
[1]Faye E J, Wang H W, Kawaguchi S,et al. Reinstatement ofGrateloupia subpectinata(Rhodophyta, Halymeniacae)based on morphology andrbcL sequences [J].Phycological Research, 2004, 52(1): 59—67
[2]Holmes E M. A new JapaneseGrateloupia[J].The Scottish Botanical Review, 1912, 1: 208
[3]Yendo K. Notes on Algae New to Japan. II [M]. Tokyo:Botanical Magazine. 1914, 263—281
[4]Okamura K. Japanese Algae [M]. Tokyo: Uchida-rokakuho. 1936, 964
[5]Yoshida T, Kawaguchi S. Marine Algae of Japan [M].Tokyo: Uchida-Rokakuho. 1998, 697—734
[6]De Clerck, Gavio O B, Fredericq,et al. Systematics ofGrateloupia filicina(Halymeniaceae, Rhodophyta), based onrbcL sequence analysis and morphological evidence,including the reinstatement ofG. capensissp. nov [J].Journal of Phycology, 2005, 41(2): 391—410
[7]Verlaque M P, Brannock M, Komatsu T,et al. The genusGrateloupiaC. Agardh (Halymeniaceae, Rhodophyta) in the Thau Lagoon (France, Mediterranean): a case study of marine plurispecific introductions [J].Phycologia, 2005,44(5): 477—496
[8]Nelson W A, Kim S Y, Boo S M,et al. The first record ofGrateloupia subpectinatafrom the New Zealand region and comparison withG. prolifera, a species endemic to the Chatham Islands [J].Botanica Marina, 2013, 56(5-6):507—513
[9]Yang M Y, Kim M S. Taxonomy ofGrateloupia(Halymeniales, Rhodophyta) by DNA barcode marker analyis and a description ofPachymeniopsis volvitasp.nov [J].Journal of Applied Phycology, 2015, 27(3):1373—1384
[10]Xia B M. Flora Algarum Marinarum Sinicarum[A]. In:Tomus II Rhodophyta. Part III. Gelidiales Cryptonemiales Hildenbrandiales [C]. Beijing: Science Press. 2004,59—81 [夏邦美. 中国海藻志. 见: 第二卷 红藻门, 第三册 石花菜目 隐丝藻目 胭脂藻目. 北京: 科学出版社.2004, 59—81]
[11]Wang H W, Kawaguchi S, Horiguchi T,et al. Reinstatement ofGrateloupia catenata(Rhodophyta, Halymeniaceae) on the basis of morphology andrbcL sequences[J].Phycologia, 2000, 39(3): 228—237
[12]Sherwood A R, Presting G G. Universal primers amplify a 23S rDNA plastid marker in eukaryotic algae and cyanobacteria [J].Journal of Phycology, 2007, 43(3):605—608
[13]Kawaguchi S, Wang H W, Horiguchi T,et al. A comparative study of the red algaGrateloupia filicina(Halymeniaceae) from the Northwestern Pacific and Mediterranean with the description ofGrateloupia asiaticasp.nov [J].Journal of Phycology, 2001, 37(3): 433—442
[14]Wang H W, Kawaguchi S,et al. A morphological and molecular assessment of the genusPrionitisJ. Agardh(Halymeniaceae, Rhodophyta) [J].Phycological, 2001,49(3): 251—261
[15]Miller K A, Hughey J R, Gabrielson P. Research note:First report of the Japanese speciesGrateloupia lanceolata(Halymeniaceae, Rhodophyta) from California, USA[J].Phycological, 2009, 57(3): 238—241
[16]Lee J I, Kim H G, Geraldino P J L,et al. Molecular classification of the genusGrateloupia(Halymeniaceae,Rhodophyta) in Korea [J].Algae, 2009, 24(4): 231—238
[17]Sherwood A R, Sauvage T, Kurihara A,et al. The Hawaiian Rhodophyta Biodiversity Survey (2006-2010):a summary of principal findings [J].Bmc Plant Biology,2010, 10(1): 258
[18]Li F, Jiang P, Zhao S Y,et al. Research on the revision ofGrateloupia fastigiataLi et Ding (Rhodophyta,Halymeniaceae) based on morphological observations andrbcL sequence analyses [J].Acta Hydrobiologica Sinica,2016, 40(6): 1249—1256 [李芳, 姜朋, 赵树雨, 等. 帚状蜈蚣藻的修订研究—基于形态观察和rbcL序列分析.水生生物学报, 2016, 40(6): 1249—1256]
[19]Koichiro T, Daniel P, Nicholas P,et al. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods [J].Molecular Biology and Evolution, 2011,28(10): 2731—2739
[20]Chiang Y M. Morphological studies of the red algae of the family Cryptonemiaceae [J].University of California Publications in Botany, 1970, 58: 1—95
[21]Li W X, Fu P F, Ding Z F,et al. Studies on the reproductive organs ofGrateloupia filicina(MU LF) C.AG [J].Tropic Oceanology, 1984, 3(4): 35—40 [李伟新, 付鹏飞,丁镇芬, 等. 蜈蚣藻及其生殖器官的研究. 热带海洋,1984, 3(4): 35—40]
[22]Kawaguchi S, Shimada S, Wang H W,et al. The new genusYonaguniaKawaguchi et Masuda (Halymeniaceae,Rhodophyta), based onY. tenuifoliaKawaguchi et Masuda sp. nov. from southern Japan and includingY. formosana(Okamura) Kawaguchi et Masuda comb. nov.from Southeast Asia [J].Phycologia, 2004, 40(1):180—192
[23]Gargiulo G M, Morabito M, Manghisi A. A reassessment of reproductive anatomy and postfertilization development in the systematics ofGrateloupia(Halymeniales,Rhodophyta) [J].Cryptogamie,Algologie, 2013, 34(1):3—35
[24]Zhao D, Wang H W, Sheng Y W,et al. Morphological observation andrbcL gene sequences studies of two new species,Grateloupia dalianensisH. W. Wang et D. Zhao sp. nov. andG. yinggehaiensisH. W. Wang et R. X. Luan sp. nov. (Halymeniaceae, Rhodophyta) from China [J].Acta Oceanological Sinica, 2012, 31(2): 109—120
[25]Yu L, Wang H W, Luan R X.Grateloupia tenuisWang et Luan sp. nov. (Halymeniaceae, Rhodophyta): a new species from South China Sea based on morphological observation andrbcL gene sequences analysis [J].Biomed Research International, 2013, Dec 23. doi: 10.1155/2013/560163
[26]Liu M, Wang H W, Luan R X. Morphological observation andrbcL sequence analysis of a new species from China,Grateloupia boaoensisWang et Luan sp. nov.(Halymeniaceae, Rhodophyta) [J].Acta Hydrobiologica Sinica, 2014, 38(5): 939—944
[27]Guan Y, Wang H W, Zhao F Q,et al.Grateloupia huanghaiensissp. nov. (Halymeniaceae, Rhodophyta), a peculiarnew species from China [J].Marine Biology Research,2015, 11(4): 396—404
[28]Cao C C, Liu M, Guo S R,et al.Grateloupia ramosaWang amp; Luan sp. nov. (Halymeniaceae, Rhodophyta)a new species from China based on morphological evidence and comparativerbcL sequences [J].Chinese Journal of Oceanology and Limnology, 2016, 34(2):283—294
[29]Cao C C, Li Y Z, Wang H W. Morphological observation,life history andrbcL gene sequence analysis of a new species,Grateloupia variatasp. nov. (Halymeniaceae,Rhodophyta) from Qingdao, China [J].Marine Biology Research, 2016, 12(4): 443—453
[30]Wang H W, Guo S R, Zhang X M,et al. Re-identifyingGrateloupia yangjiangensis(Rhodophyta, Halymeniaceae) based on morphological observations, life history andrbcL sequence analyses [J].Acta Oceanological Sinica, 2014, 33(4): 77—84
[31]Zhang L B. International Code of Botanical Nomenclature [M]. St. Louis: Missouri Botanical Garden Press.2007 [张丽兵. 国际植物命名法规. 圣路易斯: 密苏里植物园出版社. 2007]
RESEARCH ON THE REVISION OF GRATELOUPIA DIDYMECLADIA LI ET DING BASED ON MORPHOLOGICAL OBSERVATIONS AND MOLECULAR ANALYSES
LIU Fang, TIAN Yi-Lin and WANG Hong-Wei
(College of Life Sciences, Liaoning Normal University, Dalian 116081, China)
With the combination of morphological observation andrbcL (ribulose-1, 5-bisphosphate carboxylase/oxygenase) andCOⅠ (Partial cytochrome c oxidaseⅠ) gene sequence analyses, eleven samples ofG.didymecladiaLi et Ding, collected from Shantou, Wenzhou and Dalian in China, were re-examined. The results indicated that: (1) the thalli were characterized as solitary or caespitose habit, in red-brown or dark-red color, fleshy and gelatinous in texture;they became cartilaginous at an older age with the height of 15—50 cm. The flattened main axes were erect, 3—15 mm in width, 1 mm in thickness, pinnately branched 1—3 times, bearing abundant branches with opposite or alternated arrangement. The end portion of the main axes was tapering. Numerous proliferous branchlets were on the surface and margin of the main axes, constricted at the base, of various lengths reaching as long as 15 cm; Gametophytes dioecious,the samples had five-celled carpogonial branches and four-celled auxiliary branches (5cpb-4auxb model), they are typicalGrateloupia-type. Tetrasporangia were formed from the inner cortex cells, cruciately divided. These morphological features were the same asG. subpectinataHolmes. (2) According to the phylogenetic tree constructed byrbcL gene sequences, the elevenG. didymecladiasequences in this study had no pairwise divergence and formed a single monophyletic subclade. Intraspecific divergence exists between the eleven samples andG. subpectinatafrom Korea and Japan, ranged from 1 bp (0.08) to 2 bp (0.17%). According to the phylogenetic tree constructed byCOⅠgene sequences,the eleven sequences were identical and formed a single monophyletic subclade. The pairwise distance ofCOⅠsequences between the samples andG. subpectinatafrom Korea was 1 bp (0.18%), also belonging to intraspecific difference.G. didymecladiaandG. subpectinatawere demonstrated to be the same species based on the morphological observation and molecular analysis. Based on the priority rule,G. didymecladiawas classified as the synonym ofG. subpectinata. This is to date the first discovery ofG. subpectinataexisting in China, indicating a new record species.
Rhodophyta; Halymeniaceae;Grateloupia didymecladia;Grateloupia subpectinata; Morphological observations; Molecular analyses
2016-12-30;
2017-05-14
国家自然科学基金(31270251、30870161、31570209)资助 [Supported by the National Natural Science Foundation of China(31270251, 30870161, 31570209)]
刘芳(1991—), 女, 吉林省辽源市人; 硕士研究生; 主要从事海藻分类学研究。E-mail: 172307320@qq.com
王宏伟(1963—), 男, 辽宁北镇人; 博士, 教授, 博士生导师; 主要从事海藻分类及分子系统学研究。E-mail: kitamiwang@163.com
10.7541/2017.158
