眼斑拟石首鱼重复DNA序列的染色体定位
2017-11-29朱齐春刘贤德王志勇蔡明夷
朱齐春 郑 娇 张 静 刘贤德 王志勇 蔡明夷
(集美大学水产学院, 农业部东海海水健康养殖重点实验室, 厦门 361021)
眼斑拟石首鱼重复DNA序列的染色体定位
朱齐春 郑 娇 张 静 刘贤德 王志勇 蔡明夷
(集美大学水产学院, 农业部东海海水健康养殖重点实验室, 厦门 361021)
针对眼斑拟石首鱼Sciaenops ocellatus染色体标记匮乏的问题, 利用荧光原位杂交(FISH)定位了眼斑拟石首鱼的18S rDNA、5S rDNA和端粒序列。结果显示, 眼斑拟石首鱼的核型公式为2n=48t; 仅有1对18S rDNA位点, 位于第1对染色体的次缢痕部位; 有2对5S rDNA位点, FISH信号强度不等, 强信号位于第8对染色体的近着丝粒端, 弱信号位于第3对染色体的臂间。端粒FISH信号出现于所有染色体的两端, 但表现出染色体两端信号不平衡的特点, 着丝粒端FISH信号明显强于远端信号。这一特点为判定染色体的方向提供了便利。结合其他石首鱼的核型数据可以推断, 2n=48t的核型及单对近着丝粒分布的18S rDNA位点是石首鱼的共同祖征; 在石首鱼进化过程中, 曾发生活跃但不影响宏观核型的小规模重排。研究结果丰富了眼斑拟石首鱼染色体的辨识标记, 并为研究石首鱼染色体进化提供了基础数据。
眼斑拟石首鱼; 核型; 染色体; 荧光原位杂交; 核糖体DNA; 端粒
染色体是遗传物质的载体。染色体的形态和结构特征是研究真核生物的基因组结构、系统发生、进化、分类和育种的重要资料。1970年以来,鱼类染色体核型数据不断积累, 至今报道过核型的鱼类大约有3425种或亚种, 约占现存鱼类的12.2%[1,2]。近年来, 分子克隆、PCR和荧光原位杂交等新技术逐步应用于鱼类染色体研究, 极大地拓展了鱼类染色体研究内容和深度。
眼斑拟石首鱼(Sciaenops ocellatus), 也称红姑鱼或黑斑红鲈, 属鲈形目, 石首鱼科, 拟石首鱼属,主要分布在从新泽西到佛罗里达的大西洋和墨西哥湾沿岸。1991年引入我国, 因其抗病力强、生长迅速、适温和适盐性广、存活率高、耐低温, 非常适合高密度养殖, 很快成为我国重要海水养殖鱼类[3,4]。尤锋等[3]和王晓艳等[5]分别分析了眼斑拟石首鱼的核型(2n=48t, NF=48), 并将核仁组织区(NOR)定位于1号染色体的近着丝粒区, 但目前尚未见FISH定位相关报道。为了进一步揭示眼斑拟石首鱼的核型特征, 本研究应用FISH技术在眼斑拟石首鱼染色体上定位18S rDNA、5S rDNA和端粒序列3种重复序列, 以期为眼斑拟石首鱼的染色体识别提供稳定有效的细胞遗传学标记, 也为石首科鱼染色体进化研究提供了基础数据。
1 材料与方法
1.1 样品和染色体制备
用于制备染色体的眼斑拟石首鱼雌雄各5尾,购自福建省宁德市金铃水产有限公司。鱼体注射秋水仙碱和植物血凝素后, 取头肾细胞制备染色体,具体的操作方法按陈紫瑩所描述的方法[6]。同时采集眼斑拟石首鱼的鳍条固定于无水乙醇中备用。
1.2 探针制备
用DNA提取试剂盒(上海捷瑞生物工程有限公司)从无水乙醇固定的鳍条中提取全基因组DNA。18S rDNA部分编码序列经PCR扩增获得, 其上、下游引物序列分别为5′-CGCGCAAATTACCCAC TCCC-3′和5′-CTGAACGCCACTTGTCCCT-3′。5S rDNA的全部编码序列和间隔序列同样也用PCR扩增获得, 所用上、下游引物序列为5′-GTCAGGC CTGGTTAGTACTTGGAT-3′和5′-GGGCGCATTCAGGGTGGTAT-3′。端粒重复序列(TTAGGG),通过引物(TTAGGG)5和(TAACCC)5无模板扩增获得[7]。
18S rDNA、5S rDNA和端粒重复序列的PCR扩增均为20 μL反应体系[10×buffer (含Mg2+) 2 μL、2.5 mmol/L dNTPs 1.6 μL、10 mmol/L的上下游引物各1 μL、5 U/μL的EasyTaqDNA聚合酶 0.2 μL和50 ng基因组DNA 1 μL, 加ddH2O至20 μL]。不同目标产物对应PCR扩增条件略有不同。18S rDNA扩增条件如下: 94℃预变性4min; 94℃变性30S, 54℃退火1min, 72℃延伸1min, 循环30次; 72℃充分延伸5min。5S rDNA的扩增条件为: 94℃预变性2min, 54℃退火1min, 循环30次。端粒重复序列扩增条件为: 94℃预变性90S; 94℃变性45S, 52℃退火30S, 72℃延伸30S, 循环30次; 72℃充分延伸10min。
上述PCR扩增产物用缺口平移法制备生物素或地高辛标记探针, 操作方法参见试剂盒说明书(Roche)。
1.3 染色体载玻片制备和染色
采用蒸汽滴片法获得染色体制片, 室温风干后在60℃恒温箱中老化3h后, 室温存放备用, 具体操作同陈紫瑩等[8]的描述。部分制片用吉姆萨染液、4′,6-二脒基-2-苯基吲哚(DAPI)和碘化丙啶(PI)染色, 方法同郑娇等[9]的描述。
1.4 原位杂交、信号检测及观察
FISH操作参考前人描述[10,11], 包括制片和探针的变性、杂交过夜、严谨性洗涤、信号放大等步骤。其中, 生物素标记用亲和素-Alexa fluor-488(Introgen)显示绿色荧光信号, 地高辛标记用抗地高辛-诺丹明(Roche)显示红色荧光信号。最后, 滴加含PI或DAPI的抗褪色剂封片。
1.5 显微观察、测量与核型排列
在正置荧光显微镜(Olympus BX53)下观察染色体, 用电荷藕合器件图像传感器(CCD, Olympus DP80)拍照。选取5个染色体完整、分散良好的中期相(PI染色)的数码图像, 应用Image-pro plus 6.0(IPP 6.0)软件辅助排列并测量染色体的长度、宽度、面积和累积光密度(IOD), 方法参考前人的描述[6,9]。核型图中的染色体按相对长度降序排列。
2 结果
2.1 核型特征
选取形态清晰、分散良好及数目完整的眼斑拟石首鱼的中期分裂相显微观察统计, 结果显示眼斑拟石首鱼的细胞含有48条染色体, 核型公式为2n=48t (图1), 且雄鱼和雌鱼的染色体数目没有差异。根据染色体的形态特征和信号特征对其进行配对, 将配对的染色体按照相对长度降序排列并编号(图1b)。取分散良好的5个中期相进行染色体配对, 并按降序排列成5幅核型图, 利用Image-pro plus 6.0软件对染色体的长度、面积、宽度和IOD进行测量, 通过Excel计算核型图的同源染色体的相对长度、相对面积、相对宽度和相对IOD, 并统计5幅核型图的平均值, 结果如表1所示。数据表明,眼斑拟石首鱼染色体的相对长度在2.62%—5.45%、相对面积在2.20%—5.88%、相对宽度在3.63%—4.67%、相对IOD在2.05%—6.14%, 分布连续。值得注意的是, 吉姆萨染色在1号染色体近着丝粒区域有明显的次缢痕, 而DAPI染色在相应位置出现阴性带(图2a、图2b)。
2.2 重复序列的FISH定位
以生物素标记的端粒重复序列作探针做FISH,用PI对染色体进行染色, 染色体染为红色, 端粒信号显示绿色(图1a)。绿色信号分布在染色体两端。两端端粒信号呈现强度不平衡的特点, 着丝粒端信号强, 远端信号弱, 因而根据端粒信号强弱可以轻易的判断染色体方向, 并直观显示眼斑拟石首鱼含48条端部着丝粒染色体。18S和5S rDNA双色FISH中, 18S rDNA用生物素标记, 5S rDNA用地高辛标记, 染色体用DAPI负染。结果显示(图2c), 染色体负染为蓝色; 整个染色体组仅有一对18S rDNA信号, 呈绿色, 分布于1号染色体次缢痕部位; 2对5S rDNA信号, 均呈红色, 但信号强度不同。强信号5S rDNA位点位于8号染色体近着丝粒区域, 弱信号5S rDNA位点位于3号染色体臂间。两类rDNA位点不具同线性(图2c、图2d)。雌雄性个体间未见明显的核型差异。
3 讨论
3.1 宏观核型的特征
目前认为, 2n=48t是硬骨鱼核型的共同祖征[12]。在海水鱼中, 大多数鲈形目鱼类具有这一保守核型,石首鱼是其中核型最为保守的一个科[1]。目前已知核型的石首鱼约38种, 核型包含48条染色体有36种,核型公式为2n=48t约32种[2]。眼斑拟石首鱼的核型公式为2n=48t, 与石首鱼科的主流核型一致, 具硬骨鱼原始核型特征
3.2 rDNA序列的定位

图1 基于FISH的眼斑拟石首鱼染色体核型图Fig. 1 Karyotype of Sciaenops ocellatus based on FISH
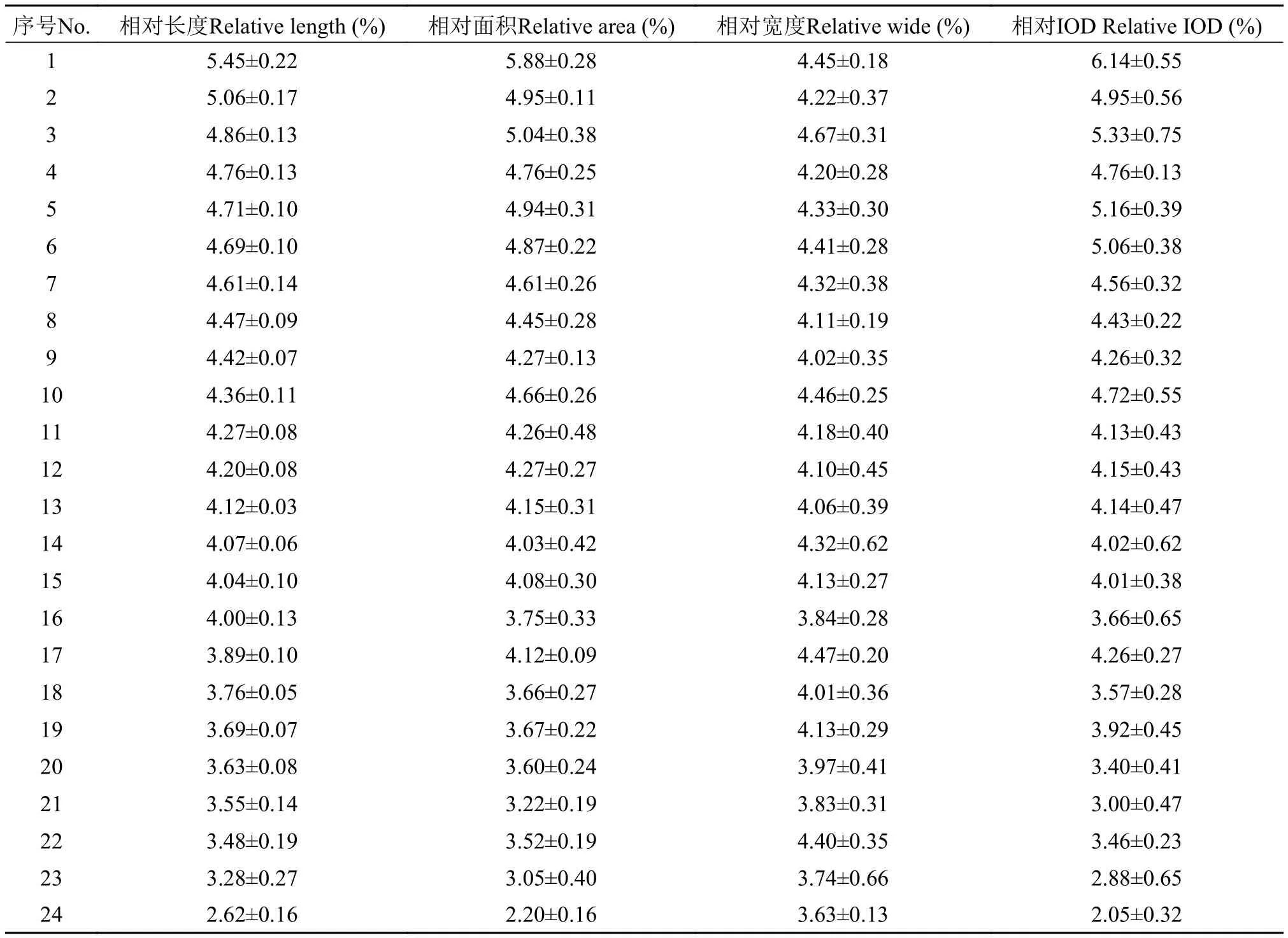
表1 眼斑拟石首鱼染色体的相对长度、相对面积、相对宽度和相对IODTab. 1 The relative length, relative area, relative wide and relative IOD of the chromosomes of Sciaenops ocellatus

图2 眼斑拟石首鱼染色体中期相Fig. 2 Metaphase of Sciaenops ocellatus
宏观核型稳定、保守是石首鱼科的基本细胞遗传学特征。然而, 在许多宏观核型稳定的海水鱼分类单元中常常观察到微小的染色体重排, 特别是rDNA位点及其邻近区域[13]。在真核生物细胞核中,rDNA组织为两个独立的基因家族, 串联重复单元分别为45S rDNA和5S rDNA。其中, 45S rDNA编码5.8S、18S和28S rRNA, 5S rDNA编码5S rRNA[14]。45S rDNA在染色体上的分布可以通过银染显示核仁组织中心(NOR)、DAPI染色显示阴性带或CMA染色体显示高亮带等方法来显示, 更直接的方法是以45S rDNA的部分序列(如18S rDNA)为探针作FISH来定位[15]。在石首鱼中, 进行过45S rDNA (或NOR)定位研究共有12种(含本研究), 其中开展过FISH定位的有5种(表2)。12种石首鱼均含有单对45S rDNA (或NOR)的位点, 但分布位置存在种间变异。其中, 厦门白姑鱼(Argyrosomus amoyensis)、美洲无鳔石首鱼(Menticirrhus americanus)、箕作黄姑鱼(Micropogonias furnieri)、黄姑鱼(N. albiflora)和眼斑拟石首鱼等5种的分布模式一致, 均在1号染色体近着丝粒区域; 另外4种分布模式各不相同, 还有3种分布位置尚未明确。根据原始性状的普遍性原则可以推断, 单对45S rDNA单对分布于1号染色体近着丝粒位置是石首鱼原始核型的特征。可见, 眼斑拟石首鱼18S rDNA位点的染色体分布仍保留着石首鱼共有祖先的特征。
5S rDNA位点的染色体分布模式只能通过FISH定位确定, 因此迄今开展过5S rDNA定位研究的石首鱼仅有4种。然而, 现有数据已初步显示, 石首鱼5S rDNA位点在染色体上分布模式存在较高水平的种间变异: 厦门白姑鱼具有1对信号; 大黄鱼具有9—11对信号; 黄姑鱼和眼斑拟石首鱼均具有2对信号, 但分布位置不同(表2)。可见, 在石首鱼进化进程中, 染色体曾发生过活跃的染色体重排, 但是这些重排没有改变宏观核型结构。这种宏观核型稳定而rDNA位点分布模式高度变化的现象也见于其他硬骨鱼类群。例如,科(Serranidae)中多数鱼类核型包含48条单臂染色体, 但45S rDNA (或NOR)存在很大的种间变化; 银鲈科尖口银鲈属(Eucinostomus)中也观察到稳定的核型和两种rDNA的位置变动[1,23]。导致rDNA位点位置差异的机制可能是转座子的作用或同源染色不平衡交换等[24,25]。在石首鱼中, 什么因素导致5S rDNA的种间变异尚需进一步研究。
双色FISH直观显示, 眼斑拟石首鱼的18S rDNA和5S rDNA位点不具有同线性。多数鱼类的45S rDNA和5S rDNA位点分布在不同的染色体上,大多数的高等植物和动物也具有同样的特征[14]。45S rDNA和5S rDNA位点分别位于不同染色体, 可以避免两个基因簇之间的易位, 并使其可以独立进化[26]。但是, 也有一些鱼类的两种rDNA位点位于同一条染色体上, 甚至在相邻的位置上, 如黄姑鱼(Nibea albiflora)有2对5S rDNA位点, FISH信号较强的一对与18S rDNA间隔分布在同一条染色体上[9];牙鲆(Paralichthys olivaceus)、大西洋鲑(Salmosalar)、杂斑盔鱼鱼(Coris julis)等鱼类的两类rDNA位点则串联分布于同一染色体[27,28]。
3.3 端粒序列的定位
端粒是一种在动植物中广泛分布的重复序列,是染色体的重要功能元件, 主要功能是维持染色体的完整性和独立性[29]。目前在定位过端粒序列的80多种鱼中, 存在端部以外端粒序列杂交信号的约42%[30]。眼斑拟石首鱼的端粒序列杂交信号均出现在染色体的两端, 未见臂间端粒信号, 说明眼斑拟石首鱼在进化过程中没有发生涉及端粒的染色体倒位或融合。值得注意的是, 眼斑拟石首鱼的端粒信号呈现出一个有趣的现象, 各条染色体上着丝粒端的端粒信号较强, 而远着丝粒端的端粒信号弱。这种染色体两端端粒信号强度不一致的现象并未见于其他石首鱼, 但偶见于其他脊椎动物和植物, 如人类和菠菜(Spinacia oleraceaL.)[29,31]。导致端粒信号强度不一致的可能原因有: (1)着丝粒端粒序列长度长于远端着丝粒; (2)着丝粒DNA含有端粒序列, 两种来源的端粒信号重叠。虽然, 导致眼斑拟石首鱼两端端粒信号不一致的原因仍然未明, 但这一现象可以直观地呈现染色体的方向, 并证明眼斑拟石首鱼的染色体全部为端部着丝粒染色体。

表2 石首鱼科鱼类的核型特点和rDNA位点定位研究概况Tab. 2 The karyotypes characteristics and summary of locations of rDNA on the chromosome of Sciaenidae
4 结论
本研究首次利用双色FISH分析了眼斑拟石首鱼的核型特征, 丰富了该物种的染色体辨识标记,并为研究石首鱼染色体进化提供了基础数据。结合其他石首鱼核型数据, 可以推断: 2n=48t的核型及单对近着丝粒分布的18S rDNA位点是石首鱼的共同祖征。尽管开展过石首鱼5S rDNA定位的石首鱼仅有4种, 现有数据已初步揭示, 石首鱼染色体进化过程中, 染色体上曾发生过活跃但不影响宏观核型的小规模重排。FISH是分析核型特征和染色体进化的有力工具。以FISH为核心技术的分子细胞遗传学是联结信息学、分子生物学和细胞学的桥梁。在鱼类基因组学和分子生物学研究飞速发展的今天, 鱼类分子细胞遗传学研究将迎来新的发展契机和更广阔的应用前景。
[1]Galetti Jr P M, Aguilar C T, Molina W F. An Overview of Marine Fish Cytogenetics [M]. Marine Genetics. Springer Netherlands. 2000, 55—62
[2]Arai R. Fish Karyotypes: a Check List [M]. Springer Science amp; Business Media. 2011, 3—4
[3]You F, Liu J, Xu C. Study on the karyotype in the red drum,Sciaenops ocellatus[J].Marine Sciences, 1998,22(2): 51—53 [尤锋, 刘静, 徐成. 美国红鱼(Sciaenops ocellatus)的核型研究. 海洋科学, 1998, 22(2): 51—53]
[4]Meilahn C W, Davis D A, Arnold C R. Effects of commercial fish meal analogue and menhaden fish meal on growth of red drum fed isonitrogenous diets [J].The Progressive Fish-culturist, 1996, 58(2): 111—116
[5]Wang X Y, Wang S F, Zhang J S,et al. The Karyotype,Ag-NORs and C-banding ofSciaenops ocellata[J].Oceanologia et Limnologia Sinica, 2012, 43(1): 84—87[王晓艳, 王世锋, 张建设, 等. 美国红鱼(Sciaenops ocellata)的核型、Ag-NORs和C-带分析. 海洋与湖沼, 2012,43(1): 84—87]
[6]Chen Z Y. Cytogenetic Study onLarimichthys croceaandNibea albiflora[D]. Thesis for Master of Science. Jimei University, Fujian. 2013 [陈紫莹, 大黄鱼与黄姑鱼细胞遗传学初步研究. 硕士学位论文, 集美大学水产学院,福建. 2013]
[7]Ijdo J W, Wells R A, Baldini A,et al. Improved telomere detection using a telomere repeat probe (TTAGGG) n generated by PCR [J].Nucleic Acids Research, 1991,19(17): 4780
[8]Chen Z Y, Wang H H, Wang K W,et al. The procedure of DPI staining to display nucleolar organizer regions in Yellow Croaker (Nibea alibiflora) [J].Journal of Jimei University, 2014, 19(3): 161—167 [陈紫瑩, 王浩华, 王凯武, 等. DPI染色显示黄姑鱼NOR的方法研究. 集美大学学报: 自然科学版, 2014, 19(3): 161—167]
[9]Zheng J, Cao K, Yang A R,et al. Chromosome mapping using genomic DNA and repetitive DNA sequences as probes for somatic chromosome identification inNibea albiflora[J].Journal of Fisheries of China, 2016, 40(8):1156—1162 [郑娇, 曹款, 杨安冉, 等. 黄姑鱼染色体识别与重复序列定位. 水产学报, 2016, 40(8): 1156—1162]
[10]Fujiwara A, Abe S, Yamaha E,et al. Uniparental chromosome elimination in the early embryogenesis of the inviable salmonid hybrids between masu salmon female and rainbow trout male [J].Chromosoma, 1997, 106(1):44—52
[11]Cai M Y, Liu X D, Chen Z Y,et al. Characterization of Pacific abalone (Haliotis discus hannai) karyotype by C-banding and fluorescencein situhybridization with rDNA[J].Journal of Fisheries of China, 2013, 37(7): 1002—1008 [蔡明夷, 刘贤德, 陈紫瑩, 等. 皱纹盘鲍染色体C带和rDNA定位. 水产学报, 2013, 37(7): 1002—1008]
[12]Brum M J I, Galetti Jr P M. Teleostei ground plan karyotype [J].Journal of Computational Biology, 1997, 2(2):91—102
[13]de Mello Affonso P R A, Galetti Jr P M. Chromosomal diversification of reef fishes from genusCentropyge(Perciformes, Pomacanthidae) [J].Genetica, 2005, 123(3):227—233
[14]Martins C, Wasko A P. Organization and evolution of 5S ribosomal DNA in the fish genome [J].Focus on Genome Research, 2004: 335—363
[15]Gornung E. Twenty years of physical mapping of major ribosomal RNA genes across the teleosts: a review of research [J].Cytogenetic and Genome Research, 2013,141(2—3): 90—102
[16]Liao M X, Wang Z Y, Zhang J,et al. The comparison of chromosome mapping of rDNA inArgyrosomus amoyensisandLarimichthys crocea[J].Journal of Fisheries of China, 2017, 41(9): 1338—1344 [廖梦香, 王志勇, 张静,等. 核糖体DNA在厦门白姑鱼和大黄鱼染色体上的比较定位. 水产学报, 2017, 41(9): 1338—1344]
[17]Gomes V, Vazzoler A E A D M, Phan V N. Estudos cariotípicos de peixes da familiaSciaenidae(Teleostei,Perciformes) da região de Cananéia, SP, Brasil: 2. sobre o cariótipo deMenticirrhus americanus(Linnaeus, 1758)[J].Brazilian Journal of Oceanography, 1983, 32(2):187—191
[18]Reggi R, Périco E, Suninsky M,et al. Estudos citogenéticos em papa-terra,Menticirrhus litoralis(Perciformes, Serranidae) [J].Simpósio de Citogenética Evolutiva e Aplicada de Peixes Neotropicais,Botucatu,SP. Unesp,Botucatu, 1986, 57
[19]Gomes V, Vazzoler AEM, Phan VN. Estudos cariotípicos de peixes da famíliaSciaenidae(Teleostei, Perciformes)da região de cananéia, sp, brasil, 1. sobre ocariótipo deMicropogonias Furnieri(Desmarest, 1823) [J].Brazilian Journal of Oceanography, 1823a, 32(2): 137—142
[20]Ojima Y, Kikuno T. Karyotypes of a gobiesociform and two perciform fishes (Teleostei) [J].Proceedings of the Japan Academy Ser B Physical amp; Biological Sciences,1987, 63(6): 201—204
[21]Accioly I V, Molina W F. Cytogenetic studies in Brazilian marine Sciaenidae and Sparidae fishes (Perciformes)[J].Genetics and Molecular Research, 2008, 7(2): 358—370
[22]Feldberg E, Porto J I R, Santos E B P,et al. Cytogenetic studies of two freshwater Sciaenids of the genusPlagioscion(Perciformes, Sciaenidae) from the Central Amazon[J].Genetics and Molecular Biology, 1999, 22(3):351—356
[23]Calado L L, Bertollo L A C, Cioffi M B,et al. Evolutionary dynamics of rDNA genes on chromosomes of theEucinostomusfishes: cytotaxonomic and karyoevolutive implications [J].Genetics and Molecular Research, 2014,13(4): 9951—9959
[24]Coen E S, Dover G A. Unequal exchanges and the coevolution of X and Y rDNA arrays inDrosophila melanogaster[J].Cell, 1983, 33(3): 849—855
[25]Merlo M A, Cross I, Manchado M,et al. The 5S rDNA high dynamism inDiplodus sargusis a transposon-mediated mechanism. Comparison with other multigene families and Sparidae species [J].Journal of Molecular Evolution, 2013, 76(3): 83—97
[26]Mantovani M, Abel S, Moreira-Filho O. Conserved 5S and variable 45S rDNA chromosomal localisation revealed by FISH inAstyanax scabripinnis(Pisces, Characidae) [J].Genetica, 2005, 123(3): 211—216
[27]Fujiwara A, Fujiwara M, Nishida-Umehara C,et al. Characterization of Japanese flounder karyotype by chromosome bandings and fluorescencein situhybridization with DNA markers [J].Genetica, 2007, 131(3): 267—274
[28]Mandrioli M, Colomba M S, Vitturi R. Chromosomal analysis of repeated DNAs in the rainbow wrasseCoris julis(Pisces, Labridae) [J].Genetica, 2000, 108(2):191—195
[29]Lan T Y, Liu B, Dong F P,et al. Multicolor FISH analysis of rDNA and telomere on spinach [J].Frontiers of Agriculture in China, 2007, 29(11): 1405—1408 [兰添颖,刘博, 董凤平, 等. 菠菜rDNA及端粒多色荧光原位杂交分析. 遗传, 2007, 29(11): 1405—1408]
[30]Ocalewicz K. Telomeres in fishes [J].Cytogenetic and Genome Research, 2013, 141(2—3): 114—125
[31]Lansdorp P M, Verwoerd N P, Fm V D R,et al. Heterogeneity in telomere length of human chromosomes [J].Human Molecular Genetics, 1996, 5(5): 685—691
LOCATION OF REPETITIVE DNA SEQUENCES ON THE CHROMOSOME OF SCIAENOPS OCELLATUS
ZHU Qi-Chun, ZHENG Jiao, ZHANG Jing, LIU Xian-De, WANG Zhi-Yong and CAI Ming-Yi
(The Key Laboratory of Helthy Mariculture for the East China Sea, Ministry of Agriculture, Fisheries College, Jimei University,Xiamen 361021, China)
The chromosome characteristics of red drum (Sciaenops ocellatus) were analyzed to enhance the cytogenetic markers inSciaenops ocellatusby fluorescencein situhybridization (FISH) with 18S rDNA, 5S rDNA and telomeric repetitive DNA as probes. The karyotype of red drum consists of 48 telomeric chromosomes. FISH with 18S rDNA probes showed a single pair of signals at the secondary constriction sites of Chromosome 1. Whereas, two pairs of signal with different intensity were detected with the 5S rDNA probes: the strong signals were on the proximal of Chromosome 8, and the weak ones were on the interstitial of Chromosome 3. The signals of telomeric repetitive DNA appeared at both ends of all chromosomes with uneven intensity, and signals at the centromeric ended stronger than those at the distal ends, which facilitated determining the direction of the chromosome. These results, together with the available cytogenetic data of other sciaenids, suggested that the karyotype of 2n=48t and single 18S rDNA locus were the symplesiomorphic cytogenetic features of sciaenids, and the active and cryptic chromosomal rearrangements may have occurred along the evolution of sciaenids. The distribution patterns of these three kinds of repetitive DNAs would be useful to assist identifying the chromosomes of red drum, and be the basic data for studying chromosome evolution in Sciaenidae.
Sciaenops ocellatus; Karyotype; Chromosome; FISH; Ribosomal DNA; Telomere
Q343.2
A
1000-3207(2017)06-1218-07
2016-12-28;
2017-04-26
国家自然科学基金(41706157和31272653); 福建省自然科学基金(2017J01449)资助 [Supported by the National Natural Science Foundation of China (41706157, 31272653); the Natural Science Foundation of Fujian Province (2017J01449)]
朱齐春(1991—), 男, 安徽安庆人; 硕士研究生; 主要从事水产生物遗传育种研究。E-mail: 17859736800@163.com
蔡明夷(1973—), 女, 教授, 博士; 主要从事水产生物遗传育种研究。E-mail: myicai@jmu.edu.cn
