铜、铁、锌、铝通过Akt/GSK-3β信号通路诱导SH-SY5Y细胞的凋亡
2017-08-23朱敬丽路赵硕吴越徐彦吉姚建王鑫鑫王德才
朱敬丽 路赵硕 吴越 徐彦吉 姚建 王鑫鑫 王德才
铜、铁、锌、铝通过Akt/GSK-3β信号通路诱导SH-SY5Y细胞的凋亡
朱敬丽 路赵硕 吴越 徐彦吉 姚建 王鑫鑫 王德才
目的 探究铜、铁、锌、铝对人神经母细胞瘤细胞(SH-SY5Y)的神经毒性作用和机制。 方法 采用50、200、400 μmol/L CuSO4,1、2、4 mmol/L FeSO4和AlCl3,50、100、200 μmol/L ZnSO4分别作用于SH-SY5Y细胞,通过噻唑蓝(MTT)法检测细胞生长活性。采用ELISA法检测细胞内Akt、磷酸化Akt(Ser473)、GSK-3β及磷酸化GSK-3β(Ser9)蛋白的表达。采用Annexin-V/PI双染法检测细胞凋亡率。 结果 铜、铁、锌、铝能明显抑制细胞活性,并呈剂量依赖性,与对照组比较差异有统计学意义(P<0.05)。铁和铝各剂量组Akt表达较对照组降低(P<0.05);各剂量组GSK-3β表达无明显差异(P>0.05);各剂量组Akt及GSK-3β的磷酸化水平低于对照组(P<0.05)。细胞凋亡率随染铜、铁、锌、铝剂量的加大而升高(P<0.05)。 结论 铜、铁、锌、铝可引起神经细胞毒性,抑制Akt活性,激活蛋白激酶GSK-3β,导致细胞凋亡。
铜; 铁; 锌; 铝; SH-SY5Y神经细胞; Akt; GSK-3β
阿尔茨海默病(AD),是一种以脑内胆碱能神经元的大量缺失和死亡为特点的神经退行性疾病。PI3K/Akt通路是一条重要的细胞存活信号通路,参与调节许多细胞凋亡的过程。通过PI3K通道,Akt在Ser473位点发生磷酸化,介导了Akt的激活[1],从而对神经元具有保护作用[2]。糖原合成酶激酶3β(GSK-3β)是最早发现的Akt的直接底物之一,活化的Akt与GSK-3β结合,使GSK-3β在Ser9位点上发生磷酸化,抑制GSK-3β的活性。Jayapalan等[3]的实验显示GSK-3β通过磷酸化Tau蛋白导致AD的发生。
研究表明,在Aβ、tau蛋白聚集区发现有高浓度的金属离子,包括铜、铁、锌、铝等[4]。铜、铁、锌、铝在阿尔茨海默病中的神经毒性作用是否通过Akt/GSK-3β信号通路诱导?为此,本研究以人神经母细胞瘤细胞为模型,评价铜、铁、锌、铝的神经毒性作用和机制。
1 材料与方法
1.1 材料 人神经母细胞瘤细胞(SH-SY5Y)由北京协和细胞资源中心提供,1640RPMI培养基购自Hyclone公司,胎牛血清购自Gibco公司,MTT购自碧云天生物技术研究所,Phospho-Akt (S473) Cell-Based Colorimetric ELISA Kit和Phospho-GSK3β (S9) Cell-Based Colorimetric ELISA Kit 购自美国ImmunoWay公司,FITC Annexin V Apoptosis Detection Kit I购自美国BD公司。细胞培养箱(美国TE-HER),倒置显微镜(日本OLYMPUS),医用超净工作台(苏州安泰空气技术有限公司),流式细胞仪(美国BD公司)。
1.2 细胞的培养及分组 实验分为对照组,铜、铁、锌、铝(高、中、低)剂量组:铜(50、200、400) μmol/L,铁(1、2、4) mmol/L,锌(50、100、200) μmol/L,铝(1、2、4) mmol/L,共计13个剂量组。细胞用含15%胎牛血清的1640培养基,置于C02培养箱(37 ℃,5%C02,100%湿度)中进行培养。
1.3 细胞活性测定(MTT法) SH-SY5Y细胞以2×105/mL的密度接种于96孔培养板中:① 对照组:不加入药品;② 实验组:铜、铁、锌、铝分组给药,每组设6个平行孔,培养24 h,每孔加10 μL5 mg/mLMTT,置于37 ℃培养箱中孵育4 h,每孔加入100 μL的DMSO,振荡10 min,使结晶物充分溶解。于570 nm波长处测定其吸光度(OD),细胞存活率(%)=(细胞处理组OD值-空白孔OD值)/(细胞对照孔OD值-空白孔OD值)×100%。
1.4 ELISA检测P-Akt,Akt和GSK-3β、p-GSK-3β 取对数生长期的细胞接种在96孔板中,待细胞长到75%-90%融合时,按实验分组进行染毒。染毒24 h后,每孔加100 μL4%甲醛固定25 min。每孔加100 μLQuench buffer,室温25 min。每孔加100 μLBlocking buffer,室温1.5 h,洗涤细胞三次。每孔加50 μL第一抗体,封板膜封板,4 ℃冰箱过夜,(注:阴性对照孔不加第一抗体)。每孔加50 μL酶标抗体,室温在振动筛上轻轻振1.5 h。每孔加50 μL底物液TMB,室温避光显色15 min,每孔加50 μL终止液,450 nm处测定各孔吸光度值。读板后,每孔加50 μL结晶紫,室温振动筛上振30 min,每孔加100 μL1%SDS,振动筛上室温振动1 h。595 nm处测定内参值,测定的相对值=(细胞内OD450-阴性对照OD450)/(内参OD595-内参阴性对照OD595)。
1.5 细胞凋亡率测定 神经细胞处理与染毒同前,于接种后24 h做检测。用0.25%胰酶-EDTA 消化细胞,于4 ℃ 1300 r/min离心 5 min,100 μL1XBinding Buffer重新悬浮细胞。将用来做补偿的细胞分到3管中,每管105个。FITC标记的Annexin-V 5 μL,再加入PI 5 μL,避光反应 15 min 后,加入400 μL Binding Buffer。立即用FACSCalibur进行流式细胞术定量检测(一般不超过 1 h), 同时以不加AnnexinV-FITC及PI的一管作为阴性对照。

2 结果
2.1 铜、铁、锌、铝对SH-SY5Y细胞活力的影响 随铜、铁、锌、铝浓度的增加,细胞活力逐渐降低。染铜组细胞活力均低于对照组 (P<0.05);200、400 μmol/L组细胞活力低于50 μmol/L组 (P<0.05)。2、4 mmol/L FeSO4组细胞活力低于对照组和1 mmol/L组 (P<0.05)。染锌组细胞活力低于对照组(P<0.05);100、200 μmol/L组细胞活力低于50 μmol/L组 (P<0.05)。4 mmol/L AlCl3组细胞活力低于对照组和1 mmol/L组(P<0.05);2 mmol/L组细胞活力低于对照组(P<0.05),见表1。
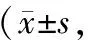
表1 铜、铁、锌、铝对SH-SY5Y细胞存活率影响%, n=6)
注:与对照组相比,*P<0.05;与低剂量组相比,△P<0.05;低、中、高剂量金属组:CuSO4分别为50、200、400 μmol/L,FeSO4分别为1、2、4 mmol/L,ZnSO4分别为50、100、200 μmol/L,AlCl3分别为1、2、4 mmol/L。
2.2 Akt与磷酸化Akt(Ser473)蛋白表达结果 随铜浓度的增加,各剂量组Akt蛋白表达差异无统计学意义(P>0.05);200、400 μmol/L组Akt磷酸化水平低于对照组和50 μmol/L组 (P<0.05)。随铁浓度增加,各剂量组Akt蛋白表达均低于对照组(P<0.05),但各剂量组间未见明显趋势变化;2、4 mmol/L组Akt磷酸化水平低于对照组和1 mmol/L组(P<0.05)。随锌浓度的增加,各剂量组之间Akt蛋白表达差异无统计学意义(P>0.05);各剂量组Akt磷酸化水平均低于对照组(P<0.05),100、200 μmol/L组磷酸化水平低于50 μmol/L组 (P<0.05)。随铝浓度增加,1、2 mmol/L组Akt蛋白表达低于对照组(P<0.05),呈逐渐降低趋势;各剂量组Akt磷酸化水平低于对照组(P<0.05),4 mmol/L组磷酸化水平低于1 mmol/L组 (P<0.05),见表2。


组别p⁃Akt(Ser473)AktCuSO4对照组0 189±0 0090 301±0 017低剂量组0 182±0 0240 286±0 013中剂量组0 112±0 018∗△0 271±0 024高剂量组0 065±0 011∗△0 277±0 012FeSO4对照组0 189±0 0090 301±0 017低剂量组0 177±0 0060 264±0 017∗中剂量组0 115±0 011∗△0 259±0 019∗高剂量组0 066±0 008∗△0 264±0 018∗ZnSO4对照组0 189±0 0090 301±0 017低剂量组0 164±0 012∗0 261±0 03中剂量组0 089±0 011∗△0 246±0 013高剂量组0 073±0 009∗△0 258±0 026AlCl3对照组0 189±0 0090 301±0 017低剂量组0 137±0 028∗0 266±0 004∗
续表:

组别p⁃Akt(Ser473)Akt中剂量组0 109±0 011∗0 260±0 015∗高剂量组0 075±0 005∗△0 259±0 015
注:与对照组相比,*P<0.05;与低剂量组相比,△P<0.05;低、中、高剂量金属组:CuSO4分别为50、200、400 μmol/L,FeSO4分别为1、2、4 mmol/L,ZnSO4分别为50、100、200 μmol/L,AlCl3分别为1、2、4 mmol/L。
2.3 GSK-3β和磷酸化GSK-3β(Ser9)蛋白表达结果 随铜、铁、锌、铝浓度的增加,各剂量组GSK-3β蛋白表达差异无统计学意义(P>0.05)。200、400 μmol/L CuSO4组GSK-3β磷酸化水平低于对照组和50 μmol/L组 (P<0.05)。各染铁组GSK-3β磷酸化水平均低于对照组(P<0.05),2、4 mmol/L FeSO4组磷酸化水平低于1 mmol/L组 (P<0.05)。100、200 μmol/L ZnSO4组GSK-3β磷酸化水平低于对照组和50 μmol/L组 (P<0.05)。2、4 mmol/L AlCl3组磷酸化水平低于对照组和1 mmol/L组 (P<0.05),见表3。
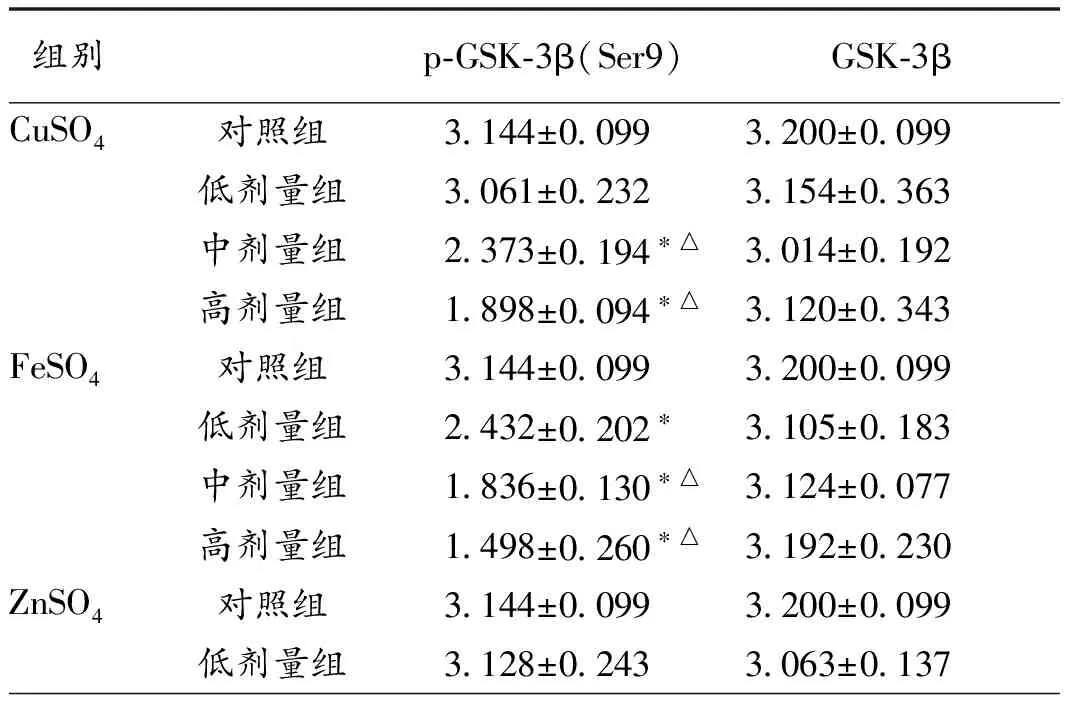
组别p⁃GSK⁃3β(Ser9)GSK⁃3βCuSO4对照组3 144±0 0993 200±0 099低剂量组3 061±0 2323 154±0 363中剂量组2 373±0 194∗△3 014±0 192高剂量组1 898±0 094∗△3 120±0 343FeSO4对照组3 144±0 0993 200±0 099低剂量组2 432±0 202∗3 105±0 183中剂量组1 836±0 130∗△3 124±0 077高剂量组1 498±0 260∗△3 192±0 230ZnSO4对照组3 144±0 0993 200±0 099低剂量组3 128±0 2433 063±0 137
续表:

组别p⁃Akt(Ser473)Akt中剂量组2 215±0 245∗△3 251±0 128高剂量组1 335±0 176∗△3 053±0 311AlCl3对照组3 144±0 0993 200±0 099低剂量组2 782±0 2963 186±0 273中剂量组1 956±0 249∗△3 014±0 110高剂量组1 367±0 206∗△3 172±0 188
注:与对照组相比,*P<0.05;与低剂量组相比,△P<0.05;低、中、高剂量金属组:CuSO4分别为50、200、400 μmol/L,FeSO4分别为1、2、4 mmol/L,ZnSO4分别为50、100、200 μmol/L,AlCl3分别为1、2、4 mmol/L。
2.4 细胞凋亡率测定结果 随染铜、铁、锌、铝浓度的增加,细胞凋亡率均高于对照组(P<0.05)。 各染铜组细胞凋亡率高于对照组(P<0.05),200、400 μmol/L组细胞凋亡率高于50 μmol/L组 (P<0.05)。2、4 mmol/L FeSO4组细胞凋亡率高于对照组和1 mmol/L组 (P<0.05)。100、200 μmol/L ZnSO4组细胞凋亡率高于对照组和50 μmol/L组 (P<0.05)。2、4 mmol/L AlCl3组细胞凋亡率高于对照组和1 mmol/L组 (P<0.05),见表4。
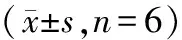
表4 铜、铁、锌、铝对SH-SY5Y细胞凋亡率影响
注:与对照组相比,*P<0.05;与低剂量组相比,△P<0.05;低、中、高剂量金属组:CuSO4分别为50、200、400 μmol/L,FeSO4分别为1、2、4 mmol/L,ZnSO4分别为50、100、200 μmol/L,AlCl3分别为1、2、4 mmol/L。
3 讨论
现有研究表明,体内微量元素动态平衡的紊乱可能与AD的形成和病理改变有关,在AD病人脑组织老年斑内存在高浓度的金属离子,主要包括铜、铝、锌、铁等[4]。GSK-3β作为多条信号通路的主要调控酶,它活性的高低与体内稳态以及疾病的发生、发展有关, GSK-3β的活化参与AD的病理生理过程[5]。GSK-3β在Ser9位点发生磷酸化后,GSK-3β的活性降低。Akt(又名蛋白激酶B,PKB)作为重要的抗凋亡调节因子, 能介导多种生物学效应。通过PI3K通道,Akt在磷酸肌醇依赖性蛋白激酶(PDKl)作用下,促使Thr308位点发生磷酸化;Akt在PDK2作用下,使Ser473位点发生磷酸化。研究表明,抑制PI3K/Akt通道可引起GSK-3β的激活[6],GSK-3β的激活可导致酶的失活以及细胞凋亡[7],同时可抑制蛋白磷酸酯酶2A(PP2A)活性,从而导致神经元功能、结构改变,学习、记忆障碍。同时研究证实,AD病人体内GSK-3β磷酸化水平增高时,可抑制tau蛋白的过度磷酸化,改善NFTs和Aβ的聚集[8]。
有研究表明,在SH-SY5Y细胞内摄入大量铜,会抑制GSK-3β磷酸化水平[9];Lovell等[10]报道给予Fe2+可提高大鼠皮质神经元tau蛋白过度磷酸化的程度和GSK-3β的活性;铁和Aβ的存在能够显著减少蛋白激酶C(PKC),降低Akt的活性[11]。景小鹏[12]研究发现,在小鼠体内摄入大量锌,GSK-3β丝氨酸 9位点磷酸化水平显著降低;Singla等[13]提出过量铝可以上调GSK-3β的表达,同时Zhao等[14]研究也发现,铝可以上调GSK-3β的表达,下调PP2A的表达,从而使tau蛋白异常磷酸化。
铜、铁、锌、铝作用于SH-SY5Y细胞后表现出一定的神经毒性。铜、铁、锌、铝进入细胞后,激活了蛋白激酶GSK-3β,下调了Akt及GSK-3β的磷酸化水平,其机制与PI3K-AKT/PKB-GSK3β信号通路有关。随铜、铁、锌、铝浓度的增加,细胞凋亡率呈逐渐增高趋势。由此可见,铜、铁、锌、铝可通过Akt/GSK-3β信号通路诱导SH-SY5Y细胞的凋亡。因此,对于Akt及GSK-3β的活性调节,可作为AD发病机制和药物靶向治疗等方面的研究重点。虽然目前金属离子在AD病理过程中的作用还没有最后定论,但是,金属离子和AD密切相关是毋庸置疑的, 测定体内金属离子的含量可能作为无创性预防AD痴呆前阶段(pre-MCI、MCI)患病风险的重要评估指标, 同时调节金属离子稳态对预防AD、研究其发病机制和研发治疗药物等都具有重要意义。
[1] Tato I,Bartrons R,Ventura F,et al. Amino acids activate mammalian target of rapamycin complex 2 (mTORC2) via PI3K/Akt signaling [J].J Biol Chem,2011,286(8):6128-6142.
[2] Griffin RJ,Moloney A,Kelliher M,et al.Activation of Akt/PKB, increased phosphorylation of Akt substrates and loss and altered distribution of Akt and PTEN are features of Alzheimer’S disease pathology [J]. J Neurochem, 2005, 93(1):105-117.
[3] Jayapalan S,Natarajan J.The role of CDK5 and GSK-3β kinases in hyperphosphorylation of microtubule associated protein tau (MAPT) in Alzheimer’s disease[J].Bio Information, 2013, 9(20): 1023-1030.
[4] Nuttall JR, Oteiza PI.Zinc and the aging brain [J].Genes Nutirt,2014, 9(1):1-11.
[5] Sato S,Tatebayashi Y,Akagi T,et al. Aberrant tau phosphorylation by glycogen synthase kinase-3beta and JNK3 induces oligomeric tau fibrils in COS-7 cells [J].J Biol Chem,2002,277 (44): 42060-42065.
[6] Wang JZ, Grundke-Iqbal I, Iqbal K. Kinases and phosphatases and tau sites involved in Alzheimer neurofibrillary degeneration[J]. Eur J Neurosci, 2007, 25(1): 59-68.
[7] Kaytor MD,Orr HT. The GSK3 beta signaling cascade and neurodegenerative disease [J].Curr Opin Neurobiol,2002,12(3):275-278.
[8] 王鹏军,张昱,宋荣蓉,等.IGF-1对Aβ-40诱导PCI2细胞tau蛋白过度磷酸化的保护作用及其机制研究[J].四川大学学报:医学版,2010,41(6):960-964.
[9] Hickey JL,Crouch PJ, Mey S. Copper(Ⅱ) complexes of hybrid hydroxyquin-oline-thiosemicarbazone ligands: GSK3β inhibition due to intracellular delivery of copper[J]. Dalton Transactions,2011,40(6):1338-1347.
[10]Lovell MA,Xiong S,Xie C,et al. Induction of hyperphosphorylated tan in primary rat cortical neuron cultures mediated by oxidative stress and glycogen synthase kinase-3β[J].Alzheimers Dis,2004,6(6):659-671.
[11]Kuperstein F, Brand A, Yavin E. Amyloid Abeta1-40 preconditions non-apoptotic signals in vivo and protects fetal rat brain from intrauter ineischemic stress[J].J Neurochem, 2004, 91(4): 965-974.
[12]景小鹏.高锌摄入导致AD样病理改变及记忆障碍[D].华中科技大学,2011.
[13]Singla N,Dhawan DK.Regulatory role of zinc during aluminium-induced altered carbohydrate metabolism in rat brain[J]. J Neurosci Res,2012, 90(3): 698-705.
[14]ZhaoHH,Di J,Liu WS,et al.Involvement of GSK 3 and PP2A in ginsenoside Rb1’s attenuation of aluminium-induced tau hyperphosphorylation[J]. Behav Brain Res,2013,241(1):228-234.
Copper, iron, zinc and aluminum induce the apoptosis of SH-SY5Y cells through Akt/GSK-3β signaling pathways
ZHUJing-li,LUZhao-shuo,WUYue,XUYan-ji,YAOJian,WANGXin-xin,WANGDe-cai.
DepartmentofNutritionandFoodHygiene;XUYan-ji.DepartmentofToxicology,PublicHealthCollege,HarbinMedicalUniversity,Harbin150081,China
Objective To explore the effects of copper, iron, zinc and aluminum on the neurotoxi-city and mechanism in SH-SY5Y cells. Methods SH-SY5Y cells were treated with different concentrations of copper sulfate (50,200,400μmol/L),iron sulfate and aluminum chloride (1,2,4 mmol/L),and zinc sulfate(50,100,200 μmol/L) respectively. Cell viabilities were measured by MTT assay. ELISA kit method were performed to evaluate the deliverances of glycogen synthase kinase-3β(GSK-3β), phosphorylation of the sites in GSK-3β-Ser9, Akt and phosphorylated Akt(Ser473). The apoptosis rate of detected cells was by double-staining with Annexin-V and PI. Results With the increase of the concentration of copper, iron, zinc and aluminum increased, the viability of SH-SY5Y cell decreased gradually. The cell viability of each dose group were significantly lower than that of control group(P<0.05). The expressions of Akt were significantly lower in iron and aluminum group, than that in control group (P<0.05). The expression of GSK-3β had no significant difference(P>0.05). The levels of phosphorylation of GSK-3β and Akt in all groups were significantly lower than those of control group(P<0.05). With the concentration of copper, iron, zinc and aluminum increased, the apoptosis rates were significantly higher than that of control group (P<0.05). Conclusions Copper, iron, zinc, aluminum could induce neurotoxicity, inhibit the activity of Akt, activate the glycogen synthase kinase 3β and lead to the neuronal apoptosis.
copper; iron; zinc and aluminum; SH-SY5Y cell; Akt; GSK-3β
国家自然科学基金(81273193)
150081黑龙江省哈乐滨市,哈尔滨医科大学公共卫生学院营养与食品卫生学教研室(朱敬丽,路赵硕,吴越,姚建,王鑫鑫,王德才)毒理学教研室(徐彦吉)
王德才,Email:decaiwang@live.cn
R 742; R 153.3
A
10.3969/j.issn.1003-9198.2017.08.007
2016-09-27)
