有关母系遗传机制的分析
2017-08-08刘明泉
刘明泉
(山东省淄博市第七中学 255499)
母系遗传是由细胞质基因控制的,但又表现出多种形式的复杂性。近年来,随着电子显微技术与分子生物学的迅猛发展,将电子显微技术与DNA分子标记应用于母系遗传的研究,使人们对母系遗传的机制有了更加深入的了解。
1 母系遗传的概念和发现
1.1 母系遗传的概念 母系遗传,是指两个具有相对性状的亲本杂交,不论正交或反交,子一代总是表现为母本性状的遗传现象。这是因为这些性状是由核外基因(如线粒体DNA、叶绿体DNA)所控制的,因此表现出母系遗传的特点。
1.2 母系遗传的发现 1909年,德国植物学家C.Correns发表了关于紫茉莉叶色遗传实验的论文,首次发现母系遗传现象。同年,植物学家E.Baur研究了天竺葵的叶片遗传,也发现类似的现象,并认为是叶绿体遗传所致。1952年,M.B.Mitchell用脉孢菌的小菌落突变品系与野生型(大菌落)进行杂交实验,其结果为后代表现型都和母本相同,以后的研究发现控制该性状的基因位于线粒体上。1962 年,Lufe 等发现一位年轻的瑞典妇女有异常增高的基础代谢率,同时伴有线粒体结构的异常和氧化磷酸化功能的异常。这是人类首次认识线粒体与人类疾病的发生有关。1962 年,H. Ris 和W. Plaut 用电子显微镜和细胞化学方法发现衣藻的叶绿体中含有DNA。同年,Nass 夫妇在小鸡线粒体中发现了DNA。细胞器DNA 的发现为母系遗传概念奠定了物质基础。1987年,Wallace等通过对线粒体DNA突变与Leber病(又称leber遗传性视神经病变,或称家族性视神经病变)之间的关系的研究,提出线粒体DNA突变可引起人类疾病,目前已发现100余种线粒体DNA突变引起的疾病,这些病也属母系遗传。从1909年至今,许多遗传学家用各种实验材料做了大量的实验,发现母系遗传是真核生物普遍的遗传方式。
2 母系遗传的机制
2.1 雄配子不贡献细胞质 C.Correns研究紫茉莉叶色遗传时发现,杂种植株的表现型完全取决于母本枝条的表现型,与提供花粉的枝条无关。无论正反交,完全表现为母系遗传,控制其遗传的基因在细胞质的质体中。由于紫茉莉胚珠中的雌配子含有大量的细胞质,其中含有大量的包括叶绿体在内的质体,而花粉管中的雄配子很少有细胞质,且不含包括叶绿体在内的质体,所以后代只有母方提供的质体。由于细胞质分裂是随机的,细胞质中的质体随机地分布到子细胞中。如受精卵的质体全部为叶绿体,则后代的枝条都为绿色;如受精卵的质体全部为白色体,则后代的枝条都为白色;如受精卵的质体既有叶绿体又有白色体,则后代的枝条有花斑、绿色、白色三种。因此紫茉莉叶色遗传符合母系遗传的特征。
南黄大麦是科学家培育的一个大麦品种,其叶浅黄色,经与叶绿色的大麦进行正反交实验,F1代的叶色不同,均表现母系遗传现象。超微结构的研究发现:传粉后,萌发出的花粉管不断伸长并首先进入助细胞,将内容物(包括一对精子,营养核和少量细胞质)从花粉管顶端的小孔释放出来,进入胚囊。在大麦等植物中,精子的细胞质在受精时确实没有进入卵细胞,而是留在了解体的助细胞中[1]。由于精子的细胞质没有进入卵细胞,当然精子的叶绿体也没有进入卵细胞,由叶绿体控制的叶色性状就只能表现为母系遗传了。
2.2 雄配子的质体、线粒体丢失或退化 通过对包括毛新杨×毛白杨、毛新杨×银腺杨等在内的22个白杨杂交组合的研究发现,其由叶绿体DNA控制的性状表现为母系遗传。进一步的研究表明,毛白杨早期的小孢子中质体是随机分布的,但在有丝分裂前期质体却成群地分布在小孢子的中央或靠近萌发孔处,而核分裂是在远离萌发孔处。这样分裂后质体就全部分配到营养细胞中,而从雄性生殖细胞中丢失了,受精卵的叶绿体全部来自母方。
通过对线粒体DNA突变引起的Leber病的研究,发现虽然精子含有线粒体且随精子细胞核进入卵细胞中,但遗传分析显示父本的线粒体不能存活,也就不能遗传给后代。进入卵细胞内的精子的线粒体被认为受到破坏并退化[2]。在超显微结构上,衰退的线粒体表现为髓磷脂状,这种衰退的细胞器的DNA不能传递给后代,从而导致线粒体的母系遗传。
2.3 雄配子的质体、线粒体DNA被核酸酶特异性识别降解 研究人员对母系遗传植物的花粉核酸酶的检测结果表明,雄性生殖细胞中细胞质DNA的消失、减少与高活性花粉核酸酶具有正相关,并且认为DNA的消失现象是一种特异性降解,也是线粒体的一种主动行为。此外,利用DNA免疫电子显微术研究发现,大麦精细胞中线粒体的DNA免疫标记比早期生殖细胞中减少了97%,说明父系线粒体DNA可能发生了降解,拷贝数的减少,使父系线粒体DNA的传递中断,导致线粒体的母系遗传[3]。
3 母系遗传中少量后代表现出父本性状或双亲性状的机制
3.1 一种配子的细胞质基因接到另一种配子的叶绿体或线粒体的DNA上 衣藻是一种单倍体生物,它有两根鞭毛和一个含有多个DNA的叶绿体。研究发现,莱茵衣藻有两种交配型(mt+和mt-),受一对等位基因控制。1954年美国遗传学家R.Sager用衣藻中链霉素抗性突变体(smr)与敏感性(sms)的不同交配型杂交:smrmt+×smsmt-→全部后代的95%为smr;smsmt+×smrmt-→全部后代的95%为sms。尽管两个亲本对合子来说贡献了相等的细胞质,但所有的后代(约95%)表现出mt+亲本对链霉素抗性的表现型,就像母系遗传现象一样。通过对杂交中两个亲本的叶绿体DNA分别以14N和15N进行标记,测得其浮力密度分别为1.69和1.70。然后按不同的组合杂交,经密度梯度离心测得合子叶绿体DNA的浮力密度(表1)[4]。
在对合子和亲本的叶绿体DNA进行比较后发现,来自mt-的叶绿体DNA丢失了,或许是被核酸酶特异性地识别而降解了。因此,杂交子代只保留下mt+亲本的叶绿体DNA,表现出母系遗传。但后代中只是95%显示单亲遗传,另外的5%为什么表现出双亲遗传呢?后来发现,某些mt-的叶绿体DNA与mt+叶绿体的DNA结合而被保存下来。
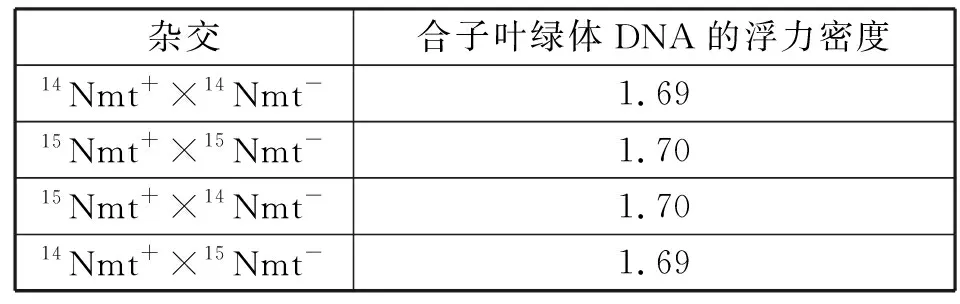
表1 莱茵衣藻杂交后合子的叶绿体DNA的浮力密度表
3.2 进入卵细胞的少量精子的质体或线粒体DNA未被识别降解 天竺葵为多年生草本,通常作一二年生花卉栽培。其突变体白色叶中只有白色质体,当天竺葵的白色突变体同正常野生型杂交时,F1植株除白色和绿色两种植株外,还出现花斑形状的植株,其比例在正反交之间也有很大的不同,也不符合孟德尔遗传。正常野生型绿叶中只有叶绿体,而花斑叶中具有绿色和白色两类质体。E.Baur研究了天竺葵的叶片遗传,做了如下的杂交组合:♀绿叶枝条×♂花斑叶枝条→F1大部分绿叶植株,少数花斑叶植株;♀花斑叶枝条×♂绿叶枝条→F1大部分花斑叶植株,少数绿叶植株。结果是:杂交后代的表现型比例在正反交之间有很大的不同,大部分表现为母本的性状。对上述结果的合理解释是:绿叶、白叶、花斑叶性状是通过细胞质中质体DNA遗传的,卵细胞贡献了大部分细胞质,精子中有质体,且精子的细胞质进入卵细胞中[1]。尽管精子带有的细胞质比卵细胞少得多,质体的总量也少得多,但这少数叶绿体DNA却未被识别降解。由于细胞质分裂是随机的,来自父本的少量质体分配到胚细胞中传给后代,从而决定了只有少数后代表现出父本的性状。近年来PCR技术证实,精子也会对受精卵提供一些线粒体DNA,这是造成线粒体DNA的异序性的原因之一[5]。
