青藏高原青稞蛋白质含量空间分异规律及其与环境因子的关系
2017-04-07王建林钟志明冯西博付刚侯维海王改花大次卓嘎
王建林,钟志明,冯西博,付刚,侯维海,王改花,大次卓嘎
(1西藏农牧学院,西藏林芝 860000;2中国科学院地理科学与资源研究所,北京 100101)
青藏高原青稞蛋白质含量空间分异规律及其与环境因子的关系
王建林1,钟志明2,冯西博1,付刚2,侯维海1,王改花2,大次卓嘎1
(1西藏农牧学院,西藏林芝 860000;2中国科学院地理科学与资源研究所,北京 100101)
【目的】揭示不同环境因子对青藏高原青稞籽粒蛋白质含量(GPC)的影响程度,完善青稞GPC空间分异与环境因子的关系,明确青藏高原不同地区青稞GPC的环境响应。【方法】利用农学和地理学相结合的研究方法,研究青藏高原青稞GPC的分布特征及其与环境因子之间的关系。【结果】在地理水平方向上,青藏高原青稞GPC总体呈现出斑块状交错分布的格局和南高北低的态势,并形成了2个青稞GPC高值区。其中一个是介于东经100.0°—102.5°、北纬35.0°—37.5°,以青海共和、贵德、门源、同德和甘肃合作为中心的青藏高原东北部高值区,这一区域青稞GPC平均值为(13.1163±0.5939)%;另一个是介于东经86.0°—92.0°、北纬28.0°—29.0°,以西藏贡嘎、拉孜、尼木、扎囊、聂拉木、堆龙德庆、桑日、康马为中心的青藏高原中南部高值区,这一区域青稞GPC平均值为(12.8715±0.6609)%;在地理垂直方向上,随着海拔升高,青稞GPC呈现出“倒N”型分布格局,即从海拔3 000 m以下的高值区(此海拔区间青稞GPC平均为(10.8650±1.8600%))随着海拔的升高,青稞GPC逐渐减少,在海拔3 000—3 300 m达到低值区。在海拔3 000—3 300 m,随着海拔的升高,青稞GPC逐渐增加,在3 600—3 900 m达到最高值区,此海拔区间青稞GPC平均为(10.8937±2.0719)%。此后,又随着海拔的升高,青稞GPC逐渐减少。影响青稞GPC的环境因子从大到小的顺序是土壤速效氮含量>抽穗-成熟期日照时数>出苗-分蘖期平均气温日较差>分蘖-拔节期平均气温日较差>拔节-抽穗期相对湿度。【结论】影响青稞GPC最大的环境因子主要是土壤因子,其次是气候因素,地理因子无明显影响。影响青稞GPC的土壤因子主要是土壤速效氮含量,气候因子主要是抽穗-成熟期日照时数、出苗-分蘖期平均气温日较差、分蘖-拔节期平均气温日较差和拔节-抽穗期相对湿度。青稞GPC含量随着分蘖-拔节期平均气温日较差、拔节-抽穗期相对湿度的增加而显著增加,但随着抽穗-成熟期日照时数、出苗-分蘖期平均气温日较差和土壤速效氮含量的增加而显著降低。
青藏高原;青稞;蛋白质含量;空间分异规律;环境因子
0 引言
【研究意义】大麦是全球栽培的第四大谷类作物,包括皮大麦(通常俗称为大麦,Hordeum vulgare L.)和裸大麦(通常俗称为青稞、元麦、裸麦,Hordeum vulgare var.nudum HK f.)2种。其适应性强,利用价值广泛,不仅广泛用于食品和饲料,还用于啤酒酿造。无论是食用、饲用还是酿造啤酒,大麦籽粒蛋白质含量(grain protein content,GPC)均是一个重要的品质性状。前者要求GPC尽可能的高,而后者则需要GPC在适当的水平。大麦GPC既受遗传控制,又易受环境的影响。根据研究,遗传因素对GPC的影响仅占20%,而环境的影响高达80%[1-7]。青藏高原既是世界大麦起源中心之一,又是中国重要的青稞种植区,这里蕴藏着多种现代农业所急需的抗旱、抗寒、耐盐碱、高品质的大麦遗传基因[8-10]。因此,研究GPC与环境因子的关系,无疑具有重要的学术价值和积极的生产意义。【前人研究进展】吕潇等[11]和王文正等[12]研究指出,大麦GPC与生育后期的日平均气温、月平均日照呈显著正相关,与月平均降水量呈显著负相关;MOLINA-CANO等[13]认为GPC主要受环境因素的影响,遗传因素的影响较小;吴晓明等[14]认为GPC在品种间、地区间以及年度间均存在着显著差异;GLEN等[15]认为饲料GPC受遗传和环境显著性影响;PRŽULJ等[16]认为大麦品质性状受温度、有效水、氮肥、土壤类型等环境因子的影响较大;CARL等[17]研究表明,不同年份、不同品种间蛋白质差异达到极显著水平。但是,不同的环境因子是如何影响大麦GPC的,目前尚存在很大争议[18-20];另外一些研究者则认为,不同地区间大麦GPC没有明显的差异[21]。【本研究切入点】迄今为止,除零星分析部分青藏高原青稞GPC与个别气象因子的关系外,尚未发现有关青藏高原青稞GPC与环境(地理、气候、土壤)因子关系的系统性研究。【拟解决的关键问题】本研究试图以青藏高原为研究区,以藏区群众主要食粮的青稞为对象,运用农学和地理学相结合的研究方法,从大尺度上研究青稞 GPC的空间分异规律,以期为揭示环境(地理、气候、土壤)因子对青藏高原青稞GPC积累的影响程度,完善大麦GPC空间分异与环境因子的关系,明确青藏高原不同地区青稞GPC的环境效应,为指导青藏高原大麦育种、栽培和加工提供理论依据。
1 材料与方法
1.1 研究区概况
研究区涉及四川、甘肃、青海、西藏等4个省(区)主要青稞种植区。研究区跨越山地草原、高寒草甸草原、高寒灌丛草甸、山地针叶林、山地灌丛草原、山地荒漠、半荒漠等 7个自然植被带,年均日照时数1 642—3 393 h,年均温-0.6—12.9℃,年均降水量150—890 mm。研究对象为当地种植的青稞,涉及 83个品种,其中地方农家品种 67个,当地推广品种 16个。
1.2 样品采集与分析
1.2.1 样点布置 根据青藏高原大气温度和降水分布格局,于不同生态种植区(北纬 27°—38°、东经79°—104°)设置83个典型样点。在样点设置时,重点选择长势整齐、成片种植的青稞田,所有样点的具体地理位置如图1所示,所有样点的地点与青稞如电子附表1所示,每个样点面积不小于120 m2。
1.2.2 植物样品采集与分析 于2014年7—9月在各样点青稞成熟期进行采样。采样时,先用GPS定位,并在每个样点随机采收青稞种子2 000 g,共采集83个样点的青稞种子。在采样的基础上,于 2015年 3月在西藏农牧学院实习农场对所采集的 83份种质进行大田种植,小区面积6 m×4 m=24 m2,随机区组排列,3次重复,田间管理略高于当地大田水平。在青稞生长期进行物候期调查,收获后及时进行考种,并用凯氏定氮法测定不同来源地和同一环境种植后青稞籽粒的蛋白质含量。
1.2.3 土壤样品采集与分析 在每个样点进行青稞种子采样的同时,设5—6个样方,于每个样方(面积不小于20 m2)中随机采集耕作层(0—30 cm)土壤样品 3份,将其装入土壤布袋,送回实验室风干后,磨细过1 mm筛,测定土壤全氮、全磷、全钾、速效氮、速效磷、速效钾、pH、有机质等理化指标。其中,土壤全氮和速效氮采用半微量凯氏法,土壤全磷和速效磷测定采用钼锑抗比色法,土壤全钾和速效钾测定采用原子吸收分光光度法,土壤有机质采用重铬酸钾氧化—外加热法、土壤pH采用电位法进行测定。

图1 采样点分布图Fig.1 The locations of sampling sites

表1 不同来源地青稞籽粒蛋白含量的F检验Table 1 F-test of GPC of naked barley varieties from different places
1.3 数据分析与处理
首先,利用同一环境条件下种植后 83份种质的GPC数据,利用ARCGIS9.3软件作其空间分布图。其次,基于在目前条件下在青藏高原跨越10个纬度、26个经度范围内,安排数十个试验点、数个品种进行统一种植试验的困难性和认为对不同来源地的青稞同一环境条件下种植后样本的GPC数据进行多重比较,找出生物统计学上蛋白质含量差异不显著的最大样点数,并利用所分析的这些样点所采收来的青稞种子的蛋白质含量与原产地的地理、土壤和气候数据,研究青稞GPC与环境因子之间的关系,其方法是可行的,其结果也是可靠的,运用逐步回归分析法建立青稞GPC与地理、气候(注:气候资料来源于中国国家气象信息中心)、土壤因子之间的关系方程。在此基础上,采用随机森林回归分析法,综合分析与青稞GPC相关性显著的各个环境因子对青藏高原青稞GPC影响的重要程度。
2 结果
2.1 青藏高原青稞GPC的分布特征
2.1.1 青藏高原青稞GPC的水平分布特征 对83个不同来源地的青稞在同一环境条件下种植后籽粒的蛋白质含量进行了测定与方差分析。结果表明,83个不同来源地的青稞GPC变幅为6.10%—14.43%,平均(10.59±2.08)%,变异系数19.69%,不同来源地的青稞GPC存在显著性差异(表1)。在此基础上,采用ARCGIS9.3软件作其空间分布图(图2),青稞GPC在青藏高原形成了2个高值区。其中,一个介于东经 100.0°—102.5°、北纬 35.0°—37.5°,以青海共和、贵德、门源、同德和甘肃合作为中心的青藏高原东北部高值区,这一区域青稞 GPC平均为(13.1163±0.5939)%;另一个介于东经86.0°— 92.0°、北纬28.0°—29.0°,以西藏贡嘎、拉孜、尼木、扎囊、聂拉木、堆龙德庆、桑日、康马为中心的青藏高原中南部高值区,这一区域青稞 GPC平均为(12.8715± 0.6609)%。同时,从图2还可以看出,青藏高原青稞GPC的水平分布总体呈现出斑块状交错分布和南高北低的趋势。

图2 青藏高原青稞GPC水平分布状况Fig. 2 GPC horizontal distribution of naked barley varieties in Qinghai-Tibet plateau
2.1.2 青藏高原青稞 GPC的垂直分布特征 从表 2可以看出,随着海拔升高,青稞GPC的变化呈现出“倒N”型的分布格局。即从海拔3 000 m以下的高值区(此海拔区间青稞 GPC平均为(10.8650± 1.8600)%)随着海拔的升高,青稞GPC逐渐减少,在海拔3 000—3 300 m达到低值区。在海拔3 000—3 300 m以上,随着海拔的升高,青稞GPC逐渐增加,在3 600—3 900 m达到最高值区,此海拔区间青稞GPC平均为(10.8937±2.0719)%。此后,又随着海拔的升高,青稞GPC逐渐减少。同时,从表2还可以看出,青稞GPC变异系数随着海拔的升高而呈现有波动性变化。

表2 不同海拔高度青稞GPC的分布状况Table 2 Naked barley varieties GPC distribution along the altitudes in Qinghai-Tibet plateau
2.2 青藏高原青稞GPC与环境因子的关系
经多重比较,发现83个样点采收来的青稞种子在同一环境条件下种植后青稞 GPC在生物统计学上差异不显著的最大样点数为36个,其青稞GPC的变幅在9.0%—11.0%。为此,利用所分析的这36个样点采收来的青稞种子的蛋白质含量与原产地的地理、土壤和气候数据,建立青稞GPC与环境因子之间的关系方程,现分述如下:
2.2.1 青稞 GPC与地理因子的关系 基于逐步回归分析法,建立青稞 GPC与地理纬度(X1)、地理经度(X2)、海拔(X3)等地理因子的关系方程如下:

从式(1)可以看出,青稞GPC(Y)与地理经度(X1)、地理纬度(X2)、海拔高度(X3)均呈负相关关系。根据标准误差检验,该回归方程未通过 α=0.05水平的显著性检验。表明青稞 GPC随着地理经度、纬度和海拔高度的增加而增加。同时也表明,地理因子对青稞 GPC的影响未达到显著性水平(P<0.05)。
2.2.2 青稞 GPC与气候因子的关系 基于逐步回归分析法,建立青稞GPC与抽穗-成熟期日照时数(X1)、出苗-分蘖期平均气温(X2)、分蘖-拔节期平均气温(X3)、拔节-抽穗期平均气温(X4)、抽穗-成熟期平均气温(X5)、出苗-分蘖期平均气温日较差(X6)、分蘖-拔节期平均气温日较差(X7)、拔节-抽穗期平均气温日较差(X8)、抽穗-成熟期平均气温日较差(X9)、出苗-分蘖期平均相对湿度(X10)、分蘖-拔期节相对湿度(X11)、拔节-抽穗期相对湿度(X12)、抽穗-成熟期相对湿度(X13)、出苗-分蘖期降水量(X14)、分蘖-拔节期降水量(X15)、拔节-抽穗期降水量(X16)、抽穗-成熟期降水量(X17)等气候因子的关系方程如下:

从式(2)可以看出,影响青稞 GPC(Y)的主要气候因子是抽穗-成熟期日照时数、出苗-分蘖期平均气温日较差、分蘖-拔节期平均气温日较差、分蘖-拔期节相对湿度、拔节-抽穗期相对湿度、抽穗-成熟期降水量,而出苗-分蘖期平均气温、分蘖-拔节期平均气温、拔节-抽穗期平均气温、抽穗-成熟期平均气温、拔节-抽穗期平均气温日较差、抽穗-成熟期平均气温日较差、出苗-分蘖期平均相对湿度、抽穗-成熟期相对湿度、出苗-分蘖期降水量、分蘖-拔节期降水量、拔节-抽穗期降水量的影响则很小。
同时,从式(2)还可以看出,青稞GPC与分蘖-拔节期平均气温日较差、拔节-抽穗期相对湿度的偏相关系数均达到显著或极显著正相关差异水平,与抽穗-成熟期日照时数、出苗-分蘖期平均气温日较差的偏相关系数均达到极显著负相关差异水平,但是与分蘖-拔期节相对湿度、抽穗-成熟期降水量的偏相关关系未达到显著性(P<0.05)差异。根据标准误差检验,该回归方程通过了α=0.01水平的显著性检验。表明青稞GPC随着分蘖-拔节期平均气温日较差、拔节-抽穗期相对湿度的增加和抽穗-成熟期日照时数、出苗-分蘖期平均气温日较差的减少而显著增加。同时也表明,气候因子对青稞GPC的影响达到极显著性水平(P<0.01)。
由于式(2)中未能反映出青稞 GPC与各生育时期平均温度的关系,为此基于逐步回归分析法,建立青稞 GPC与出苗-分蘖期平均气温(X2)、分蘖-拔节期平均气温(X3)、拔节-抽穗期平均气温(X4)、抽穗-成熟期平均气温(X5)的关系方程如下:

从式(3)可以看出,影响青稞GPC(Y)的平均气温是主要拔节-抽穗期平均气温和抽穗-成熟期平均气温,而出苗-分蘖期平均气温、分蘖-拔节期平均气温的影响则很小。同时,从式(3)还可以看出,青稞GPC与拔节-抽穗期平均气温呈正相关关系,与抽穗-成熟期平均气温呈负相关关系,但是其与青稞GPC的偏相关系数均未达到显著性(P<0.05)差异。根据标准误差检验,该回归方程未通过α=0.05水平的显著性检验。表明青稞GPC随着拔节-抽穗期平均气温(X4)的增加和抽穗-成熟期平均气温(X5)的下降而增加。同时也表明各生育时期平均温度因子对青稞 GPC的影响不显著。
2.2.3 青稞GPC与土壤因子的关系 基于逐步回归分析法,建立青稞GPC与土壤有机质(X1)、全氮(X2)、全磷(X3)、全钾(X4)、速效氮(X5)、速效磷(X6)、速效钾含量(X7)和土壤 pH(X8)等土壤因子的数学模型:

从式(4)可以看出,影响青稞GPC(Y)的主要土壤因子是全磷(X3)、全钾(X4)、速效氮(X5)、速效钾含量(X7)、土壤pH值(X8),而土壤有机质(X1)、全氮(X2)、速效磷(X6)的影响则很小。同时从式(4)还可以看出,青稞GPC与速效氮含量呈显著负相关关系。根据标准误差检验,该回归方程未通过 α=0.05水平的显著性检验。表明青稞GPC随着土壤速效氮含量的增加而显著减少。同时也表明,土壤因子对青稞GPC的整体影响未达到显著性水平(P<0.05)。
2.2.4 青稞GPC与综合因子的关系 通过以上分析可以看出,青稞 GPC与分蘖-拔节期平均气温日较差、拔节-抽穗期相对湿度的偏相关系数均达到显著或极显著正相关差异水平,与抽穗-成熟期日照时数、出苗-分蘖期平均气温日较差、土壤速效氮含量的偏相关系数均达到极显著负相关差异水平。但是,这些因子对青稞GPC的影响程度如何尚不清楚。为此,运用随机森林回归分析法,对青稞GPC影响显著的这5个因素进行综合分析(表3),这5个变量对青稞 GPC影响从大到小的顺序是土壤速效氮含量>抽穗-成熟期日照时数>出苗-分蘖期平均气温日较差>分蘖-拔节期平均气温日较差>拔节-抽穗期相对湿度。
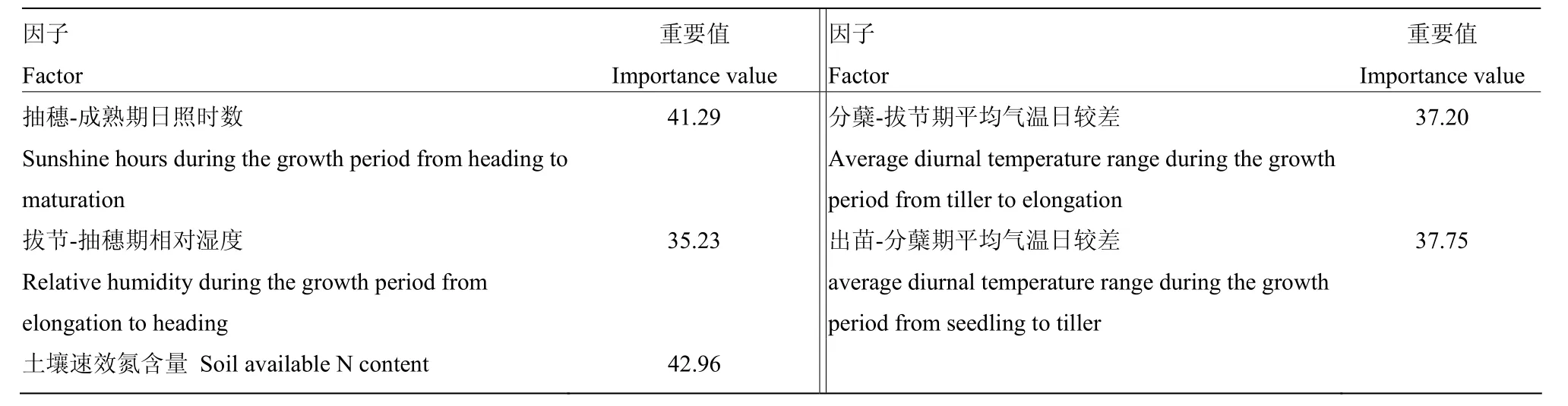
表3 不同因子对青稞GPC影响的重要程度Table 3 Importance values of different influencing factors on GPC of naked barley
3 讨论
本文认为影响青稞GPC最大的环境因子是土壤因子,其次是气候因素,地理因子无明显影响。其中,影响青稞GPC的土壤因子是土壤速效氮含量,气候因子主要是抽穗-成熟期日照时数、出苗-分蘖期平均气温日较差、分蘖-拔节期平均气温日较差和拔节-抽穗期相对湿度。本文这一研究结果与许多研究者认为大麦GPC属于多基因控制的数量性状,其表现型不仅受品种本身的遗传特性影响,也与外界的环境条件密切相关的结论一致[22-24],而与CARL等[17]对克罗地亚青稞的研究结论相反。同时,本研究认为青稞GPC与抽穗-成熟期日照时数、抽穗-成熟期平均气温、抽穗-成熟期降水量负相关关系,这一研究结果与ULLA等[25]研究表明,随着日照时间的缩短,英国二棱大麦GPC增多;NOVO等[26]认为欧洲潘诺尼亚区灌浆期较少的降水有利于蛋白质含量的增加;COLES等[27]、MAENIEOL等[28]和SAVIN等[29]认为在干旱条件下,大麦GPC显著提高;王文正等[12]研究指出大麦GPC与月平均降水量呈显著负相关;WANG等[24]认为浙江省8个大麦主栽品种在6个不同生态地区的GPC与平均降雨量显著负相关;孙立军[30]认为加拿大啤酒大麦GPC与降雨量呈负相关;AHOKAS等[31]认为降水量与法国大麦 GPC呈显著负相关关系;IMRUL等[19]认为干旱有助于西藏大麦GPC增加;张桂珍等[32]认为大麦 GPC与抽穗-成熟期降雨量呈负相关;WILLIAM等[33]认为灌溉与美国北达科他州大麦GPC呈显著负相关关系的研究结论一致,但是与DINAH等[34]认为随着生育期温度的升高,德国大麦GPC上升;CATHRINE等[35]认为灌浆成熟期温度升高有助于丹麦大麦 GPC提高;COLES等[27]、MAENIEOL等[28]、SAVIN等[29]认为灌浆-成熟期热胁迫条件下,大麦GPC显著提高;王文正等[12]研究指出大麦生育后期的日平均气温与 GPC呈显著正相关;MARTIN等[36]认为随着灌浆期降水的增多和生育后期温度的升高,德国大麦GPC上升;NOVO等[26]认为欧洲潘诺尼亚区灌浆期较高的温度有利于 GPC增加的研究结果相反,其原因尚不清楚,有待进一步深入研究。
4 结论
4.1 在地理水平方向上,青藏高原青稞GPC的水平分布总体呈现出斑块状交错分布和南高北低的格局,并形成了以青海共和、贵德、门源、同德和甘肃合作为中心的青藏高原东北部和以西藏贡嘎、拉孜、尼木、扎囊、聂拉木、堆龙德庆、桑日、康马为中心的青藏高原中南部等2个青稞GPC高值区。
4.2 在地理垂直方向上,青稞 GPC的变化呈现出“倒N”型分布格局,在海拔<3 000 m和3 600—3 900 m分别形成2个高峰区,这2个高峰区青稞GPC 的平均值分别为(10.8650±1.8600)%和(10.8937± 2.0719)%。表明,今后要选择高蛋白质的青藏高原青稞,可能从这些区域的地方品种中进行选择可能会有更大的收获。
4.3 环境因子对青稞GPC影响从大到小的顺序是土壤速效氮含量>抽穗-成熟期日照时数>出苗-分蘖期平均气温日较差>分蘖-拔节期平均气温日较差>拔节-抽穗期相对湿度。
[1] GUO B J, LUAN H Y, LIN S, LÜ C, ZHANG X Z, XU R. Comparative proteomic analysis of two barley cultivars (Hordeum vulgare L.) with contrasting grain protein content. Frontiers in Plant Science, 2016, 25: 1-11.
[2] CAI S G, YU G, CHEN X H, HUANG Y C, JIANG X G, ZHANG G P. Grain protein content variation and its association analysis in barley. BMC Plant Biology, 2013, 13: 13-35.
[3] CLANCY J A, HAN F, ULLRICH S E. The North American barley genome project: Comparative mapping of β-amylase activity QTLs among three barley crosses. Crop Science, 2003, 43: 1043-1052.
[4] SEE D K, KEPHART V, BLAKE K. Mapping genes controlling variation in barley grain protein concentration. Crop Science, 2002, 42(3): 680-685.
[5] 黄志仁, 周美学, 黄圣友, 张志胜. GPC的遗传研究. 中国大麦文集: 第二集. 西安: 陕西科学技术出版社, 1991: 56-61.
HUANG Z R, ZHOU M X, HUANG S Y, ZHANG Z S. Genetic research on protein content of barley//Collection of Barley Essays in China: Second volume. Xi’an: Science and Technology of Shaanxi Press, 1991: 56-61. (in Chinese)
[6] 陆炜, 孙立军, 张京. 我国大麦品种蛋白质、赖氨酸、淀粉含量. 中国大麦文集: 第二集. 西安: 陕西科学技术出版社, 1991: 32-35.
LU W, SUN L J, ZHANG J. Protein, lysine and starch content of barley varieties in our country. Collection of Barley Essays in China: Second volume. Xi’an: Science and Technology of Shaanxi Press, 1991: 32-35. (in Chinese)
[7] XU S YI, CAI R X, WANG R B, CHENG B, ZHU J. Genetic analysis of Some kernel quality characters in two-rowed barley. Acta Agriculturae Zhejiangensis, 1995, 9(2): 71-77.
[8] 马得泉. 中国西藏大麦遗传资源. 北京: 中国农业出版社, 2000.
MA D Q. Genetic Resources of Barley in Tibet, China. Beijing: China Agricultural Press, 2000. (in Chinese)
[9] 王建林, 栾运芳, 大次卓嘎. 西藏栽培大麦变种组成和分布规律研究. 中国农业科学, 2006, 39(11): 2163-2169.
WANG J L, LUAN Y F, DA C Z G. Research on variant composition and distribution regulation of cultivated barley in Tibet. Scientia Agricultura Sinica, 2006, 39(11): 2163-2169. (in Chinese)
[10] 王建林. 西藏高原作物栽培学. 北京: 中国农业出版社, 2012. WANG J L. Crop Cultivation in Tibet Plateau. Beijing: China Agricultural Press, 2012. (in Chinese)
[11] 吕潇, 林澄菲, 杨铮. 中国大麦品种资源蛋白质含量的生态分析//中国大麦文集:第三集. 南昌: 江西科学技术出版社, 1993: 41-44.
LÜ X, LIN C F, YANG Z. Ecological analysis of protein content of barley varieties resources in China//Collection of Barley Essays in China: Third volume, Nanchang: Science and Technology of Jiangxi Press, 1993: 41-44. (in Chinese)
[12] 王文正, 吕潇, 黎秀卿. 黄淮冬大麦区蛋白质含量与气候因子的关系. 大麦科学, 1998, 2: 9-10.
WANG W Z, LÜ X, LI X Q. The relationship between protein content and climatic factors in winter barley zone in Huanghe-Huaihe plain. Barley Science, 1998, 2: 9-10. (in Chinese)
[13] MOLINA-CANO J L, FRANCESCH M. PEREZ-VENDRELL A M, RAMO T, VOLTTAS J, BRUFAU J. Genetic and environemental variation in malting and feed quality of barley. Journal of Cereal Science, 1997, 25: 37-47.
[14] 吴晓明, 陈锦新, 吴桂玉. 啤酒大麦品种的蛋白质含量和环境变异研究. 大麦科学, 2004(1): 27-29.
WU X M, CHEN J X, WU G Y. Research on content and environmental variation of protein in malting barley varieties. Barley Science, 2004(1): 27-29. (in Chinese)
[15] GLEN F M, GABRIELA B, PETER F, DAVID P. Genetic and environmental analysis of NIR feed quality predictions on genotypes of barley (Hordeum vulgare L.). Field Crops Research, 2011, 120: 380-386.
[16] PRŽULJ N V, MOMČILOVIĆ J S, MIROSAVLJEVIĆ M. Effect of growing season and variety on quality of spring two-rowed barley. Genetika, 2014, 46(1): 59-73.
[17] CARL G, WYNSE B, MICHAEL K, WADE T. Grain composition of Virginia winter barley and implications for use in feed, food, and bio-fuels production. Journal of Cereal Science, 2010, 51: 41-49.
[18] CATHRINE H I, RENÉ G, JOHANNES R J, TEIS N M, ANDERS S, RIKKE B J. Grain protein concentration and harvestable protein under future climate conditions. A study of 108 spring barley accessions. Journal of Experimental Botany, 2016(17): 1-8.
[19] IMRUL M A, CAO F B, HAN Y, UMME A N, ZHANG G P, WU F B. Differential changes in grain ultra-structure, amylase, protein and amino acid profles between Tibetan wild and cultivated barleys under drought and salinity alone and combined stress. Food Chemistry, 2013, 141: 2743-2750.
[20] AHOKAS I, NASKALI L. Geographic variation of a-amylase, β-amylase, β-glucanase, pullulanase and chitinase activity in germinating Hordeum spontaneum barley from Israel and Jordan. Genetica, 1990, 82: 73-78.
[21] ŠIMIC G, LALIC A, HORVA D, ABIČIĆ I, BERAKOVIC I. β-glucan content and β-glucanase activity of winter and spring malting barly culitivar. Acta Alimentaria, 2015, 44 (4): 542-548.
[22] BATHGATE G N. Quality requirement for malting. Aspects Applied Biology, 1987, 15: 18-32.
[23] SMITH D B. Barley seed protein and its effects on malting and brewing quality. Plant Varieties Seeds, 1990, 3: 63-80.
[24] WANG J M, ZHANG G P, CHEN J X. Cultivar and environmental effects on protein content and grain weight of malting barley. Journal of Zhejiang University, 2001, 27: 503-507.
[25] ULLA R M H, ANNIKA W, SILJA H, KAISA P, PETER R S. Day-length effects on protein localization affect water absorption in barley (Hordeum vulgare) grains. Journal of the Science of Food and Agriculture, 2012; 92: 2944-2951.
[26] NOVO P, VOJISLAVA M, JASMINA S, MILAN M. Effect of growing season and variety on quality of spring two-rowed barley. Gentika, 2014, 46: 59-73.
[27] COLES G D, JAMIESON P D, FLASLEMORE R M. Effects ofmoisture stress on malting quality in Triumph barley. Journal of Cereal Science, 1991, 14: 161-177.
[28] MACNICOL P K, JACOBSEN J V, KEYS M M, STUART L M. Effects of heat and water stress on malt quality and grain parameters of Schooner barley grown in cabinets. Journal of Cereal Science, 1993, 18: 61-68.
[29] SAVIN R S, NICOLAS M E. Effects of short periods of drought and high temperature on grain growth and starch accumulation of two malting barley cultivars. Australia Journal of Plant Physiology, 1996, 23: 201-210.
[30] 孙立军. 赴美国、加拿大啤酒大麦考察. 大麦科学, 2001, 3: 1-4.
SUN L J. Investigation of malting barley in America and Canada. Barley Science, 2001, 3: 1-4. (in Chinese)
[31] AHOKAS H I, NASKALI L. Geographic variation of a-amylase, β-amylase, β-glucanase, pullulanase and chitinase activity in germinating Hordeum spontaneum barley from Israel and Jordan. Genetica, 1990, 82: 73-78.
[32] 张桂珍, 邱以孝. 啤酒大麦品质性状的研究. 河北农业技术师范学院学报, 1998(3): 46-48.
ZHANG G Z, QIU Y X. Research on the quality traits of malting barley. Journal of Hebei Agrotechnical Teachers College, 1998(3): 46-48. (in Chinese)
[33] WILLIAM B S, UPENDRA M, SAINJU T, CAESAR-TON T, WILLIAM M I. Malt barley yield and quality affected by irrigation. tillage. crop rotation and nitrogen fertilization. Crop Economics, Production & Management, 2015, 7(6): 7-19. (in Chinese)
[34] DINAH R, GISELA J, SYLVIA S, BETTINA E L. Temperature stress during flowering time affects yield and quality parameters of waxy barley. Landbauforschung Volkenrode, 2013, 63(1): 79-84.
[35] CATHRINE H I, RENÉ G, JOHANNES R J, TEIS N M, ANDERS S, RIKKE B J. Journal of Experimental Botany, 2016, 17: 1-8.
[36] MARTIN E, REMY M, GISELA J, SYLVIA S, ANDREAS P, HANS-JOACHIM W. Effects of free-air CO2enrichment and nitrogen supply on grain quality parameters and elemental composition of wheat and barley grown in a crop rotation. Agriculture, Ecosystems and Environment, 2010, 136: 59-68.
(责任编辑 李莉)
Spatial Distribution Regulation of Protein Content of Naked Barley Varieties and Its Relationships with Environmental Factors in Qinghai-Tibet Plateau
WANG JianLin1, ZHONG ZhiMing2, FENG XiBo1, FU Gang2, HOU WeiHai1, WANG GaiHua2, Da-cizhuoga1
(1Tibet Institute of Agriculture and Animal Husbandry, Linzhi 860000, Tibet;2Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101)
【Objective】 The objective of this study is to reveal the effect of different environmental factors on grain protein content (GPC) of naked barley, to understand the relationship between naked barley GPC distribution characteristics andenvironmental factors, and to get clear on GPC acclimation of naked barley varieties from different areas to the environment of the Qinghai-Tibet plateau. 【Method】 Based on the method of agronomy combined with geography, naked barley GPC distribution characteristics in Qinghai-Tibet plateau was studied. 【Result】 Along the horizontal direction in Qinghai-Tibet plateau, GPC spatial distribution characteristics of naked barley varieties appeared in a macrocosm trend of high-in-south and low-in-north with a staggered pattern, and there developed two regions of higher GPC. One of the regions of higher GPC exists between the area with longitudes from 100.0°E to 102.5°E and with latitudes from 35.0°N to 37.5°N. And the average GPC of barley varieties in this region is (13.1163±0.5939)% in north-eastern Qinghai-Tibet plateau with distribution centers including Gonghe County, Guide County, Menyuan County and Tongde County in Qinghai Province, and Hezuo County in Gansu Province. Another region of higher GPC existed between the area with longitudes from 86.0°E to 92.0°E and with latitudes from 28.0°N to 29.0°N. And the average GPC of barley varieties in this region is (12.8715±0.6609)% in south-central Qinghai-Tibet plateau with distribution centers including Gongga County, Lazi County, Nimu County, Zhanang County, Nielamu County, Duilongdeiqing County, Sangri County and Kangma County in Tibet. Along the vertical direction with altitudes, GPC distribution pattern appeared in shape of inverted letter N. The average GPC value is (10.8650±1.8600)% at the altitudes below 3 000 m. GPC gradually decreased with altitude increasing, and then reached to the lower value at the altitudes from 3 000 m to 3 300 m. And GPC gradually increased with altitude increasing at the altitudes above from 3 000 m to 3 300 m, and then reached to the highest value at the altitudes from 3 600 m to 3 900 m with average GPC value of (10.8937±2.0719)%. Hereafter, GPC of barley varieties decreased gradually with altitude increasing. The magnitude of effect of environmental factors on naked barley GPC follows an order of soil available N content > sunshine hours during the growth period from heading to maturation > average diurnal temperature range during the growth period from seedling from tiller > average diurnal temperature range during the growth period from tiller to elongation > relative humidity during the growth period from elongation to heading. 【Conclusion】 Soil factors are the main environmental factors with the greatest impact on GPC, and followed by climatic factors, the effect of geographical factors on GPC is not apparent. Soil available N content is the main soil factor influencing barley GPC, and sunshine hours during the growth period from heading to maturation, average diurnal temperature range during the growth period from seedling from tiller, average diurnal temperature range during the growth period from tiller to elongation and relative humidity during the growth period from elongation to heading are the main climatic factors. Barley GPC increased with the increase of average diurnal temperature range during the growth period from tiller to elongation and relative humidity during the growth period from elongation to heading, though decreased significantly with the increase of sunshine hours during the growth period from heading to maturation and average diurnal temperature range during the growth period from seedling to tiller and soil available N content.
Qinghai-Tibet plateau; naked barley; protein content; spatial distribution regulation; environmental factor
2016-10-14;接受日期:2016-12-16
国家自然科学基金(31360300、31370458、31560362)、西藏自治区重点项目(XZXTCX-2015-03、XZXTCX-2016、XZZWCXPT-2015-01)、西藏农牧学院雪域英才工程人才发展支持计划(2015XYB01)、作物学科建设项目(2015ZWXKJS、2016ZWXKJS)
联系方式:王建林,E-mail:xzwangjl@126.com。通信作者钟志明,E-mail:zhongzm@igsnrr.ac.cn
