解淀粉芽孢杆菌产凝乳酶发酵优化
2016-12-28淳于彦洁吴雅雯李金宝丁重阳
王 楠,苑 畅,汪 瑞,淳于彦洁,吴雅雯,李金宝,丁重阳
(1. 江南大学生物工程学院工业生物技术教育部重点实验室,江苏无锡214122;2. 江南大学粮食发酵工艺与技术国家工程实验室,江苏无锡214122)
解淀粉芽孢杆菌产凝乳酶发酵优化
王 楠1,2,苑 畅1,汪 瑞1,淳于彦洁1,吴雅雯1,李金宝1,丁重阳1,2
(1. 江南大学生物工程学院工业生物技术教育部重点实验室,江苏无锡214122;2. 江南大学粮食发酵工艺与技术国家工程实验室,江苏无锡214122)
对实验室前期筛选到的一株解淀粉芽孢杆菌BacillusamyloliquefaciensJNU002进行培养基和培养条件优化。单因素实验确定最佳碳源为乳糖,除麸皮外,其他氮源对产酶无明显促进作用,KH2PO4和CaCO3等对产酶有促进作用。通过正交试验确定最佳培养基成分为麸皮16 g/L、乳糖10 g/L、KH2PO41 g/L、CaCO31 g/L,其中麸皮对解淀粉芽孢杆菌产凝乳酶影响较大。在发酵温度37 ℃、种子液培养时间18 h、接种量0.5%、500 mL摇瓶装液量100 mL时,较适宜解淀粉芽孢杆菌产凝乳酶。优化后解淀粉芽孢杆菌产凝乳酶活力达到12 766 SU/mL,比优化前提高了3.5倍。
解淀粉芽孢杆菌;凝乳酶;优化
凝乳酶是干酪制造过程中起凝乳作用的关键性酶,可专一性水解κ-酪蛋白肽链的Phe105-Met106之间的肽键,破坏牛奶稳定结构,使牛奶凝结,并对干酪的质构和特殊风味的形成起重要作用[1]。目前,将能够使乳凝固的一类蛋白酶(如胃蛋白酶、植物蛋白酶、微生物蛋白酶等)统称为凝乳酶[2]。
传统用于奶酪生产使用的小牛皱胃酶受到来源和道德人文等多方面限制,已经不能满足工业生产需求,各国学者开始不断寻找凝乳酶新来源。近些年来,凝乳酶替代品的研究主要集中在3个方面:动物、植物和微生物。研究证实,哺乳动物如小猪、海豹、羊羔、水牛、猫等的胃中[3-6]和木瓜、无花果、菠萝、生姜、合欢等[7-10]植物中均发现有凝乳效果的蛋白酶。微生物生长周期短、产酶种类多,不受原料供应、季节等因素的限制,后期提取方便,在凝乳酶替代品的研究中显现出明显的优势,如微小毛霉[11]、米黑毛霉[12-14]、枯草芽孢杆菌和粟疫霉等均可产凝乳酶。丁明亮等[15]对一株产凝乳酶的枯草芽孢杆菌进行发酵条件优化,凝乳酶活达1 129 SU/mL。Wu等[16]对一株产凝乳酶的枯草芽孢杆菌进行响应面优化,得到凝乳酶活为1 048.02 SU/mL,较初始提高了428%。杭锋等[17]筛选到一株高产凝乳酶的类芽孢杆菌,麸皮是其发酵产酶的理想培养基质,对其进行培养基和培养条件优化后,凝乳酶活达6 000 SU/mL。
本研究以实验室前期筛选到的1株解淀粉芽孢杆菌为出发菌株,通过单因素实验和正交设计,进行发酵培养基和培养条件的优化,以期为其工业化生产应用提供一定参考。
1 材料和方法
1.1 实验材料
解淀粉芽孢杆菌Bacillusa-m-y-l-o-l-i-q-u-e-f-a-c-i-e-n-sJNU002,江南大学国家工程实验室筛选并在中国典型培养物保藏中心保藏(CCTCC No.M2011045)。
麸皮,购于江苏省无锡市青山市场;完达山脱脂奶粉,黑龙江省完达山乳业股份有限公司;福林酚试剂为生物纯,其余试剂均为市售国产分析纯。
1.2 培养基
种子培养基/初始发酵培养基(g/L):麸皮10,无水葡萄糖18,CaCO33,NaCl 5,MgSO45,KH2PO42;pH自然,装液量100 mL,121 ℃灭菌30 min。
牛奶平板培养基(g/L):脱脂奶粉30、琼脂粉20;pH自然,115 ℃灭菌20 min。
1.3 培养方法
平板活化:挑取甘油管中保藏的菌种在牛奶平板划线,37 ℃培养36~40 h。
种子培养:从牛奶平板上挑取一环活化后的单菌落,接种于装有100 mL液态培养基的500 mL三角瓶中,37 ℃、200 r/min条件下培养20 h。
发酵培养:以体积分数0.3%的接种量接入发酵培养基中,培养条件与种子液相同。
1.4 酶活的测定
粗酶液的制备:取发酵液于12 000 r/min离心5 min去除菌体和不溶物,上清液即为待测粗酶液。
凝乳酶活力(MCA)测定采用Arima法[11]。
蛋白酶活力(protease activity,PA)的测定方法采用Arima 1967年提出的修正的Anson的方法:向2.5 mL、1.2%的酪蛋白溶液中加入0.5 mL酶溶液,35 ℃下准确反应10 min后加入2.5 mL(0.44 mol/L)三氯乙酸(TCA)结束反应。取1 mL反应后的上清液加入5 mLNa2CO3(0.55 mol/L)溶液中,并加1 mL福林酚试剂。混合物在40 ℃下保温15 min显色,并于660 nm处测定其吸光度。
(1)
式中:OD660—在660 nm波长下吸光度;M—吸光常数;V—反应液体积;T—酶液水解反应时间(10 min);D—酶液稀释倍数。
1.5 发酵培养基成分对产酶的影响
1.5.1 麸皮添加量对产酶的影响
由于实验菌株筛选自麸皮中,前期实验已验证麸皮对产酶有明显促进作用,改变初始培养基中麸皮质量分数为1.0%、1.2%、1.4%、1.6%、1.8%和2.0%,测定产酶情况,确定麸皮添加量。
1.5.2 碳源对产酶的影响
在麸皮添加量优化后的基础上,分别选取1.8%蔗糖、乳糖、半乳糖、麦芽糖、糊精、马铃薯淀粉、可溶性淀粉、玉米粉和甘油等为碳源,其他条件不变,以葡萄糖培养组为对照,测定产酶情况,确定最优碳源。
选取最优碳源,调整添加量为0.5%、1%、1.5%、2%、2.5%、3%和3.5%,其他条件不变,测定产酶情况,确定最佳碳源添加量。
1.5.3 氮源对产酶的影响
在上述麸皮和碳源优化基础上,添加0.5%的有机氮源如酵母粉、胰蛋白胨、鱼粉蛋白胨、脲、干酪素、玉米浆、牛肉膏、豆粕粉等和0.25%无机氮源如KNO3、NH4Cl、NH4HCO3、(NH4)2SO4,其他条件不变,以不添加氮源培养基为对照,测定产酶情况,确定最优氮源。
1.5.4 无机盐对产酶的影响
以优化麸皮和碳源后的培养基为对照(不添加无机盐),分别添加1、3和5 g/L的NaCl、KCl、NaH2PO4、KH2PO4、MgSO4、MgCl2和0.2、0.6、1.0 g/L的CaCl2和CaCO3,测定产酶情况,确定无机盐作用。
1.5.5 微量金属离子对产酶的影响
以优化麸皮和碳源后的培养基为对照(不添加无机盐),分别添加2和5 mg/L的微量金属离子,测定产酶情况,确定金属离子作用。
1.5.6 正交试验
选取单因素实验中对产凝乳酶影响较大的几种离子进行正交试验。
1.6 发酵条件对产酶的影响
将优化培养基接种后分别放置在31、33、35、37和39 ℃摇床上培养,比较其产酶情况。种子液接种量分别为0.1%、0.2%、0.3%、0.5%、0.7%、0.9%、1%、3%、5%和7%,37 ℃培养,比较其产酶情况。将培养12、16、18、20、22、24、28、32和36 h种子液按最佳接种量接种,37 ℃下培养,比较其产酶情况。500 mL三角瓶中装液量依次采用60、80、100、120、140、160、180和200 mL,比较其产酶情况。
2 结果与讨论
2.1 培养基成分对产酶的影响
2.1.1 麸皮含量对产酶的影响
麸皮成分复杂,纯麸皮培养基可支持B.amyloliquefaciensJNU002正常生长,前期已验证麸皮对产酶有明显促进作用。培养基中麸皮质量分数与产酶关系如图1所示。由图1可知:初始培养基摇瓶培养24 h后,各组酶活均达最高值,且其产酶水平在一定范围内随麸皮质量分数增加而上升,麸皮质量分数为16 g/L时,凝乳酶活最高,此时MCA/PA也为最高值。根据MCA和MCA/PA变化,选取16 g/L麸皮含量进行下一步优化。
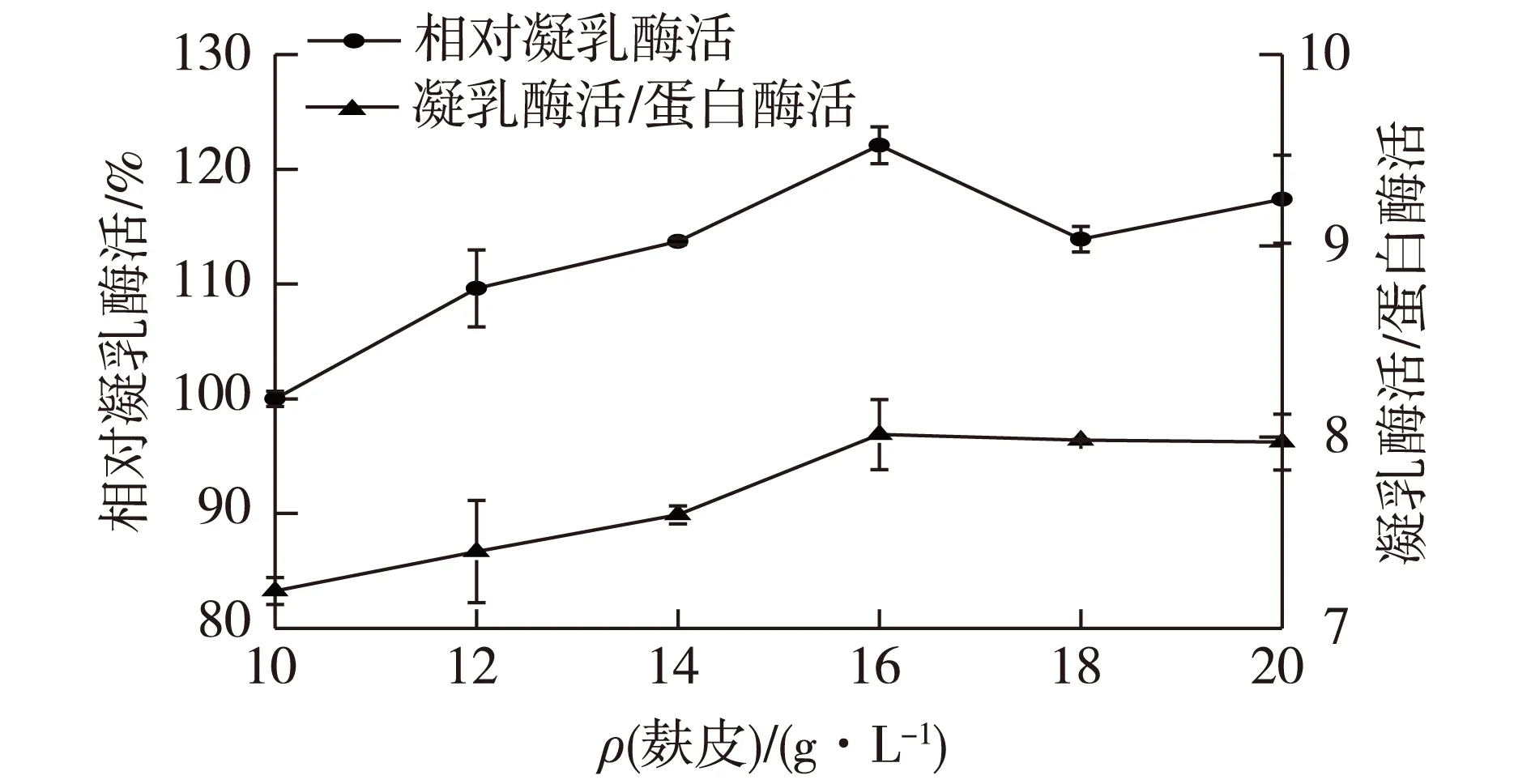
图1 麸皮质量分数对凝乳酶活力和MCA/PA值的影响Fig.1 Effects of wheat bran concentration on milk-clotting enzyme activity and MCA/PA
2.1.2 不同碳源对产酶的影响
不同碳源对B.amyloliquefaciensJNU002产酶的影响结果如图2所示。由图2可见:壳聚糖对其产酶有抑制作用,添加后明显抑制菌体生长及产酶。葡萄糖、蔗糖、麦芽糖和可溶性淀粉等碳源效果相近,乳糖和半乳糖为碳源时,菌体产酶水平明显高于其他碳源,综合考虑生产成本等原因,选取乳糖为最优碳源。

图2 不同碳源对解淀粉芽孢杆菌产酶影响Fig.2 Effects of different carbon sources on milk-clotting enzyme activity and MCA/PA
2.1.3 乳糖添加量的确定
乳糖添加量对B.amyloliquefaciensJNU002产酶的影响结果如图3所示。由图3可知:当乳糖添加量大于5 g/L时,解淀粉芽孢杆菌产酶已经达到较高水平。随着乳糖量的增加,MCA逐渐上升,但当乳糖超过20 g/L时,MCA有下降趋势;乳糖质量浓度为35 g/L时,由于碳源过量对菌体生长及产酶产生抑制作用,其产酶水平明显降低。综合考虑产酶成本与酶活水平,确定乳糖添加量为10 g/L。

图3 乳糖添加量对产酶的影响Fig.3 Effects of additive amount of lactoseon milk-clotting enzyme activity
2.1.4 不同氮源对产酶的影响
氮源是微生物生长和合成酶必需的营养成分之一,常用氮源可分为两大类:有机氮源和无机氮源。以不添加氮源培养基为对照,研究不同氮源对B.a-m-y-l-o-l-i-q-u-e-f-a-c-i-e-n-sJNU002产酶的影响,结果如图4所示。由图4可以看出:添加有机氮源时,培养基产酶水平较之空白对照均下降;添加无机铵盐后,菌体产凝乳酶水平明显下降,添加硝酸盐对产酶无明显影响。可见添加氮源并不利于解淀粉芽孢杆菌产凝乳酶。此结果与吴进菊等[18]、王艳萍等[19]实验结果一致,可能由于麸皮本身已含有足够解淀粉芽孢杆菌正常生长代谢所需的氮源。
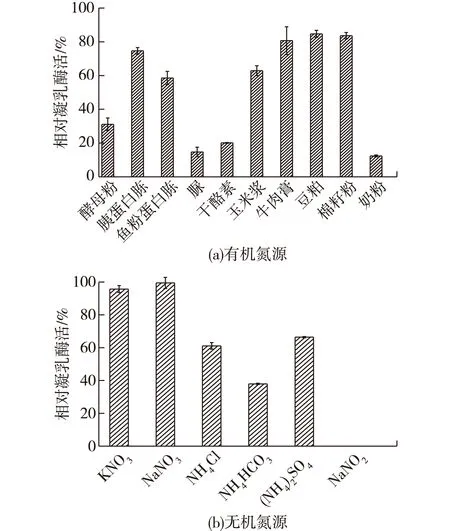
图4 氮源添加量对产酶的影响Fig.4 Effects of different nitrogen sourceon milk-clotting enzyme activity
2.1.5 无机盐对产酶的影响结果
无机盐对微生物生长有重要作用,如构成细胞的组成成分、调节细胞渗透压、参与酶的合成和分泌等。考察无机盐对B.amyloliquefaciensJNU002产酶的影响,结果见图5。由图5可知:各组产凝乳酶水平均有所上升,其中添加5 g/L 的MgSO4、1 g/L的CaCl2和0.2 g/L的CaCO3效果最好。由于CaCO3添加量少且实验组在24 h即达到最高产酶水平,后期可能由于发酵液pH上升导致发酵液酶活下降,而其余实验组均在48 h达到最高产酶水平。从发酵周期考虑,选取CaCO3为首选无机盐离子,其他无机盐离子将在此基础上进一步选择。

图5 添加不同无机盐对产酶的影响Fig.5 Effects of different inorganic salton milk-clotting enzyme activity

图6 添加CaCO3基础上不同无机盐对产酶的影响Fig.6 Effects of different inorganic salt on milk-clotting activity after adding CaCO3
为防止由于添加不同无机盐离子间相互作用使培养基有效成分减弱,故在添加CaCO3基础上再次进行无机盐离子优化,以添加CaCO3后培养基为对照,结果如图6所示。由图6可知:添加MgSO4和MgCl2后酶活明显下降,钾盐和钠盐对产酶有明显促进作用,且KH2PO4和NaH2PO4较KCl和NaCl对解淀粉芽孢杆菌产凝乳酶促进效果更佳,可能是由于磷酸盐不仅为微生物生长提供了磷源,KH2PO4和NaH2PO4与CaCO3还可以形成缓冲体系,为微生物提供稳定的生长环境。
2.1.6 微量金属离子对产酶的影响结果
复杂成分培养基往往已经含有多种微量金属离子,以不添加金属离子实验组为对照,研究微量金属离子对产酶的影响,结果如图7所示。由图7可知:添加CuCl2、ZnSO4、MnCl2、NiCl2和CoCl2等微量金属离子后,解淀粉芽孢杆菌产酶水平略有所上升。但考虑到几种金属离子均为重金属离子,为方便后续处理和环境因素,故培养基不添加微量金属离子。
2.1.7 正交试验结果与分析
选取单因素实验中对产酶影响明显的4个因素麸皮、乳糖、KH2PO4和CaCO3进行正交试验,正交试验结果和极差分析如表1和表2所示。
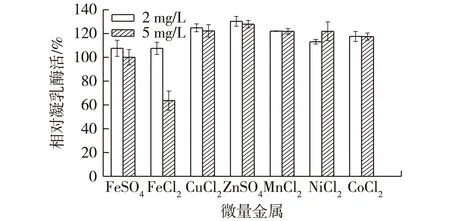
图7 添加微量金属对产酶的影响Fig.7 Effects of different amounts of other metal ionson milk-clotting activity

表1 正交实验结果Table 1 The result analyses of orthogonal experiment
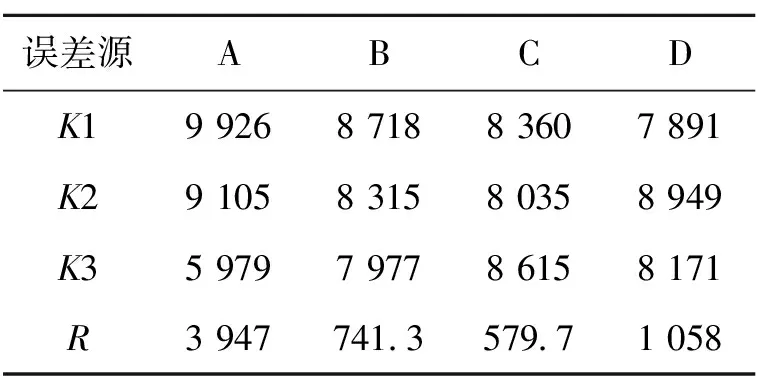
表2 极差分析Table 2 Range analysis
由表1和表2可知,4个考察因素的极差大小顺序从大到小依次为A(麸皮添加量)、 D(CaCO3添加量)、B(乳糖质量浓度)、C(KH2PO4质量浓度)。由此可见,麸皮的添加量对其凝乳酶影响最大。实验确定最优组合培养基为麸皮16 g/L,乳糖10 g/L,KH2PO41 g/L,CaCO31 g/L。
2.1.8 验证实验结果
由于得到的最优发酵培养基参数不在正交试验设计的9组内,因此需要进行验证实验。采用最优组合培养基进行平行验证实验,得到凝乳酶最高酶活为10 369 SU/mL。
2.2 培养条件优化对产酶的影响
2.2.1 发酵温度对产酶的影响
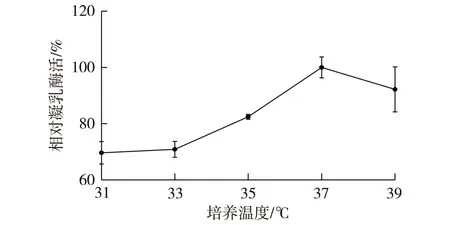
图8 发酵温度对菌株产酶的影响Fig.8 Effect of culture temperature onmilk-clotting enzyme activity
发酵温度对微生物生长、产物合成代谢、发酵液的性质等都有重要影响。发酵温度过低,菌体生长缓慢,产酶延滞期较长,增加生产成本;发酵温度过高,不适宜菌体生长,可能影响发酵产物活性。菌体发酵温度对B.amyloliquefaciensJNU002产凝乳酶影响如图8所示。由图8可知:在一定范围内,随着发酵温度的提高,菌体产酶水平逐渐上升,37 ℃为最适培养温度。温度大于37 ℃时,产酶水平下降,故选取37 ℃为最适培养温度。
2.2.2 接种量对产酶的影响
接种量指接入种子液和培养基体积的比值,对菌体的生长速度和发酵周期都有一定影响。以优化前接种量0.3%为对照,比较不同接种量对B.a-m-y-l-o-l-i-quefaciensJNU002产酶的影响,结果如图9所示。由图9可以看出,接种量对解淀粉芽孢杆菌产酶影响相对较小,接种量在0.3%~0.7%范围内,产酶能保持较高水平,其中接种量为0.5%时产酶效果最好,而后随着接种量的增加,产酶水平逐渐下降并趋于稳定。

图9 不同接种量对产酶的影响Fig.9 Effects of inoculum amount on milk-clotting enzyme activity
2.2.3 种子液培养时间对产酶的影响
种子液培养时间影响发酵过程中菌体的生长状态,适宜菌龄的种子液可以缩短发酵周期。以培养24 h种子液为对照,研究种子液培养时间对B.amyloliquefaciensJNU002产酶的影响,结果如图10所示。由图10可知:种子液培养时间对接种后发酵液最终酶活影响较小,为缩短接种后延滞期,确保最佳接种状态,选择指数生长中后期即18 h种子液接种。

图10 种子液培养时间对凝乱酶活力的影响Fig.10 Effects of culture time on milk-clotting enzyme activity
2.2.4 摇瓶装液量对产酶的影响
解淀粉芽孢杆菌是耗氧微生物,装液量多少在很大程度上影响发酵液溶氧水平,装液量越少,传氧系数越大,进而影响菌体生长和产酶。考察摇瓶装液量对B.amyloliquefaciensJNU002产酶的影响,结果如图11所示。由图11可知:随着摇瓶装液量的增加,酶活逐渐下降,装液量大于140 mL时,酶活迅速下降。考虑到发酵过程装液量过少时相对蒸发损耗较多,故选择500 mL摇瓶中100 mL装液量。

图11 装液量对凝乱酶活力的影响Fig.11 Effect of liquid volume on milk-clottingenzyme activity
3 结论
从实验室前期筛选到的1株解淀粉芽孢杆菌B.amyloliquefaciensJNU002出发,采用单因素实验和正交试验实验对菌株产凝乳酶进行了培养基优化和培养条件优化,结果表明,麸皮16 g/L、乳糖10 g/L、KH2PO41 g/L、CaCO31 g/L、温度37 ℃、种子液培养时间18 h、接种量0.5%、500 mL摇瓶装液量100 mL时,较适宜解淀粉芽孢杆菌产凝乳酶。优化后培养基酶活达12 766 SU/mL,比优化前提高了3.5倍。
[1] 张红梅,刘钟滨.凝乳酶的研究进展[J].同济大学学报(医学版),2004(3):254-257.
[2] 普燕,张富春.干酪用牛凝乳酶替代品的研究进展[J].食品与发酵工业,2015,41(5):227-233.
[3] FOLTMANN B,LONBLAD P,AXELSEN N H.Demonstration of chymosin (EC 3.4.23.4) in the stomach of newborn pig[J].Biochem J,1978,169(2):425-427.
[4] JENSEN T,AXELSEN N H,FOLTMANN B.Isolation and partial characterization of prochymosin and chymosin from cat[J].Biochim Biophys Acta,1982,705(2):249-256.
[5] SHAMSUZZAMAN K,HAARD N F.Purification and characterization of a chymosinlike protease from the gastric mucosa of harp seal (Pagophilusgroenlandicus)[J].Can J Biochem Cell Biol,1984,62(8):699-708.
[6] BAUDYS M,FOUNDLING S,PAVLIK M,et al.Protein chemical characterization ofMucorpusillusaspartic proteinase:amino acid sequence homology with the other aspartic proteinases,disulfide bond arrangement and site of carbohydrate attachment[J].FEBS Lett,1988,235(1/2):271-274.
[7] 孟广震.凝乳酶研究进展[J].微生物学通报,1987(2):92-94.
[8] BELOZERSKY M A,SARBAKANOVA S T,DUNAEVSKY Y E.Aspartic proteinase from wheat seeds: isolation,properties and action on gliadin[J].Planta,1989,177(3):321-326.
[9] MAZORRA-MANZANO M A,PEREA-GUTIERREZ T C,LUGO-SANCHEZ M E,et al.Comparison of the milk-clotting properties of three plant extracts[J].Food Chem,2013,141(3):1902-1907.
[11] ARIMA K,YU J,IWASAKI S,et al.Milk-clotting enzyme from microorganisms: V.purification and crystallization of mucor rennin fromMucorpusillusvar.Lindt[J].Appl Microbiol,1968,16(11):1727-1733.
[12] GARG S K,JOHRI B N.Rennet:current trends and future research[J].Food Rev Int,1994,10(3):313-355.
[13] STERNBERG M Z.Crystalline milk-clotting protease fromMucormieheiand some of its properties[J].J Dairy Sci,1971,54(2):159-167.
[14] 李学朋,师希雄,冯瑞章,等.米黑毛霉产凝乳酶固体发酵培养基优化[J].食品工业科技,2013(7):130-133.
[15] 丁明亮,欧阳安然,王望斐,等.枯草芽孢杆菌产凝乳酶发酵条件的优化[J].食品科学,2011(3):156-160.
[16] WU F C,CHANG C W,SHIH I L.Optimization of the production and characterization of milk clotting enzymes byBacillussubtilisnatto[J].Springerplus,2013,2(1):1-10.
[17] 杭锋,洪青,陶源,等.Paenibacillussp.BD3526发酵小麦麸皮生产凝乳酶[J].食品与发酵工业,2015,42(2):35-40.
[18] 吴进菊,徐尔尼,张凤英,等.中国曲中凝乳酶高产菌株的筛选及产酶条件的研究[J].中国食品学报,2009,9(1):124-129.
[19] 王艳萍,闵军涛,姚四平,等.印度毛霉MJ229固态发酵产凝乳酶的研究 [J].中国酿造,2013,32(1):17-21.
(责任编辑 管珺)
Optimizing of medium and fermentation conditions for milk-clottingenzyme produced by Bacillus amyloliquefaciens
WANG Nan1,2,YUAN Chang1,WANG Rui1,CHUNYU Yanjie1,WU Yawen1,LI Jinbao1,DING Zhongyang1,2
(1. Key Laboratory of Industrial Biotechnology of the Ministry of Education,School of Biotechnology,Jiangnan University,Wuxi 214122,China; 2. National Engineering Laboratory for Cereal Fermentation Technology,Jiangnan University,Wuxi 214122,China)
We optimized of culture medium and fermentation conditions to produce milk-clotting enzyme produced byBacillusamyloliquefaciensJNU002.The results of single factor experiments showed that the best carbon source was lactose;nitrogen source had no obvious promoting effect;KH2PO4and CaCO3had promoting effect on enzyme production.Through the orthogonal experiment, the optimum culture medium composition was obtained: bran 16 g/L,lactose 10 g/L,KH2PO41 g/L,CaCO31 g/L.Bran had great influence on milk-clotting enzyme production.The culture temperature was 37 ℃,incubation time of liquid seed was 18 h,inoculum amount was 0.5%,the liquid volume 100 mL/500 mL.In the optimized medium and under the optimized culture condition,milk-clotting enzyme activity ofBacillusamyloliquefaciensJNU002 reached 12 766 SU/mL an increase of 3.5 times.
Bacillusamyloliquefaciens; milk-clotting enzyme; optimization
10.3969/j.issn.1672-3678.2016.06.010
2016-04-04
“十二五”国家科技支撑计划(2013BAD18B02);国家星火计划重点项目(2015GA690004);江苏省重点研发计划(现代农业)项目(BE2015306);江苏省产学研合作项目(BY2015019-36)
王 楠(1990—),女,河北石家庄人,研究方向:发酵过程优化控制;丁重阳(联系人),教授, E-mail:zyding@jiangnan.edu.cn
TQ925
A
1672-3678(2016)06-0054-07
